纯林和混交林3 种植物光合特征研究
2023-03-31陈峰峰赵江平乔雨宁陈云明
陈峰峰 赵江平 乔雨宁 赵 敏 陈云明,3
(1. 西北农林科技大学水土保持研究所,黄土高原土壤侵蚀与旱地农业国家重点实验室,陕西 杨凌 712100;2. 水利部黄河水利委员会黄河上中游管理局,陕西 西安 210021;3. 中国科学院水利部水土保持研究所,陕西 杨凌 712100)
绿色植物光化学反应是生态系统能量流动和物质循环的基础,以光合作用为基础研究植物生产力形成过程及其生长发育的状况一直是植物生理学的研究热点[1]。光合作用日变化进程反映植物生理生态规律,通过观测植物光合日动态过程,明确光合作用日变化规律及光温因子对日变化的影响,可为植物生长及科学管理提供一定理论依据[2];光响应曲线描述净光合速率(Pn)与光合有效辐射(PAR)的相关关系,通过光响应模型拟合的表观量子效率(Φ)、光饱和点(LSP)、光补偿点(LCP)、最大净光合速率(Pnmax)等光合参数,能揭示植物光合能力及叶片活性强弱[3],二者均为探究植物光合生理特性及环境适应性的重要手段[4]。
常见的光响应模型有:直角双曲线模型(RH)、非直角双曲线模型(NRH)、指数方程模型(EM)和直角双曲线修正模型(MRH)[5]。光响应模型在不同树种间会显示出其各自的适应性,其中,毛竹(Phyllostachys Pubescens)适宜RH、NRH 模型,而MRH 是长白落叶松(Larix olgensis)最优拟合模型[6-7],朱世忠[8]通过对山西太行山和陕西黄陵3 个油松(Pinus tabuliformis)种源的光响应曲线进行拟合,结果表明MRH 模型拟合效果最佳。因此,筛选不同植物适宜的光响应模型对探究植物光合适应性及环境适应性有重要意义。有研究表明,在不同造林模式下,植物Φ、LSP、LCP 和暗呼吸速率(Rd)等光合参数也有所差异[9]。汪荣等[10]研究毛乌素沙地灌灌混交林植物光合生理特性,得出长柄扁桃(Amygdalus pedunculata)与紫穗槐(Amorpha fruticosa)混交林光合参数均优于纯林;吴旭等[5]在探究沙棘(Hippophae rhamnoides)、油松与刺槐(Robinia pseudoacacia)乔灌混交林光合生理特性中发现混交沙棘光合生理参数整体呈高态势,混交油松整体呈低态势。
黄土丘陵区,生态环境脆弱[11],人工林成为该区生态系统恢复的关键因素与主要途径[12]。抗寒耐旱能力较强的油松、连翘(Forsythia suspensa)和柠条(Caragana korshinskii)在该区域广泛种植,且多为纯林[13],相较于纯林,混交林可以显著地改变林内光照、温度、水分等环境条件[14],导致林内各组成树种光合生理特性的差异[6]。本研究以黄土丘陵区油松、连翘、柠条为研究对象,在植物生长旺季7 月,测定纯林与混交林中3 个树种叶片光合日变化及光响应曲线,通过分析3 个树种光合日变化规律及其差异特征,Pn与影响因子的相关性,光响应参数特征,明确不同造林模式下3 个树种的光响应适宜拟合模型和混交林的生理生态适应性,为黄土丘陵区人工林植物的光能利用稳定性及造林模式的优化配置提供参考依据。
1 研究区概况
研究区位于陕西省延安市安塞区纸坊沟流域内(109°13′46″~109°16′03″E, 36°46′42″~36°46′28″N),属于典型的黄土丘陵沟壑区,海拔1 010~1 400 m,暖温带半干旱季风气候;年均气温8.8 ℃,干燥度1.5,无霜期157~194 d。年均降水量504 mm,7—9 月降雨量占年降雨量的61.1%,且多为暴雨;年均蒸发量为1 463 mm。土壤类型为黄绵土,植被类型为暖温带落叶阔叶林向温带草原过渡的森林草原带,天然林已遭破坏,现有植被主要为20 世纪80 年代初实施综合治理及退耕还林过程中形成的人工林和封禁后恢复的天然灌丛及草地,乔木树种以油松、刺槐、侧柏(Platycladus orientalis)为主,灌木树种以连翘、柠条、沙棘和紫穗槐为主,乔、灌木林地均为单一树种生长,无混交林存在。
2 研究方法
2.1 样地布置
根据纸坊沟丘陵地坡面状况,选取坡面较为平整,坡长(超过60 m)较长的1 个典型坡面(109°15′10″E, 36°44′54″N),于中坡位设置坡度(25°)、坡向(半阳坡)均一致的5 个标准径流小区(20 m × 5 m),小区之间设置有过渡带(1 m × 20 m),采用“品”字形种植方式,在其中3 个小区内分别栽植油松、连翘和柠条纯林,在另2 个小区分别栽植混交林(油松混交连翘、油松混交柠条),混交林采用1 行乔木、2 行灌木的栽植方式进行栽植(表1),所有小区植株均为4 年生,在植物生长季内,每月对林下杂草进行清除,以保证植株正常生长。

表1 样地基本特征Table 1 Basic characteristics of the plot
将径流小区按长边平均分为上、中、下3 等份,在3 个纯林小区内,每等份中随机选取1 株(丛)待测植物,每个小区共选取3 株(丛)待测植物,在2 个混交林小区内,每等份中随机选取1 株乔木与1 丛灌木,每个小区共选取6 株(丛)待测植物,共计21 株(丛)待测植物,每株(丛)选取中上部受光充足的3 枚(簇)健康叶片,共计63 枚(簇)待测叶片。
2.2 测定方法
2.2.1 光合生理指标测定
光合作用日变化测定于2020 年7 月中旬,选择2 个连续无云的晴朗天气,采用Li-6400XT 光合测定系统(LI-COR, Inc., 美国),测定Pn等光合生理参数,测定时间为07:00—17:00,每2 h测定1 次,重复测定4 次,取3 组稳定值进行分析;气象指标采用样地旁U30-NRC 小型气象站(Onset HOBO, 美国)进行同步监测,指标包括空气温度(Tr)、大气CO2浓度(Ca)、空气相对湿度(RH)、PAR 等。按公式(1)~(2)计算水分利用效率(WUE)、气孔限制值(Ls)。
2.2.2 光响应曲线测定
光响应曲线测定于2020 年7 月中旬,选择2 个连续晴朗天气,采用便携式Li-6400XT 测定植物叶片光响应曲线,将CO2浓度设定为环境CO2浓度(约为400 μmol/mol),光强梯度设定为0、20、50、100、150、200、400、600、800、1 000、1 200、1 400、1 600、1 800、2 000 μmol/(m2·s),对所选63 枚(簇)待测叶片进行测定。第1 天上午9:00—11:30 对 油 松 纯 林 和 混 交 林 中27 枚(簇)叶片光响应曲线进行1 次测定,每枚(簇)叶片测定时间约为5 min,共耗时150 min;第2 天上午9:00—12:00 对不同造林模式下柠条(18 枚(簇))和连翘(18 枚(簇))叶片光响应曲线进行1 次测定,每枚(簇)叶片测定时间约为5 min,共耗时180 min。
2.3 光响应模型建立
采用RH 模型对光响应曲线进行拟合[15],表达式如下:
采用NRH 模型对光响应曲线进行拟合的表达式如下[16]:
采用EM 模型对光响应曲线进行拟合,假设Pn为0.999Pnmax所对应的光强为饱和光强。表达式如下[17]:
采用MRH 模型对光响应曲线进行拟合的表达式如下[18]:
式中:α为光响应曲线的初始斜率;I为光合有效辐射,后文用PAR 表示;Φ为表观量子效率;Ic为光补偿点,后文用LCP 表示;Isat为光饱和点,后文用LSP 表示,θ为曲线曲率,即反映非直角双曲线弯曲程度的曲角,β为光抑制项;γ为光饱和项。
2.4 分析方法
使用Microsoft Excel 2019 进行数据整理,采用Origin 2018 进行绘图,利用Pearson 相关性法分析植物叶片Pn与生理因子、气象因子的关系,使用光合4.1.1 软件进行模型拟合[19]。
3 结果与分析
3.1 光合生理指标日变化
由图1a 可知,纯林与混交林油松Pn日变化均呈“双峰型”曲线,且峰值出现的时间点均为10:00 和16:00,l峰 值 为8.73 μmol/(m2·s)和3.04 μmol/(m2·s),Yl峰 值 为7.86 μmol/(m2·s)和3.16 μmol/(m2·s),Yn峰 值 为5.56 μmol/(m2·s)和1.23 μmol/(m2·s);Y、Yl、Yn的Pn均值分别为2.18、2.92、2.94 μmol/(m2·s),大小次序为Yl>Yn>Y。
由图1b 可知,不同造林模式下,连翘Pn日变化均呈“双峰型”曲线,峰值出现的时间为10:00 和14:00,L峰值为12.28 μmol/(m2·s)和10.71 μmol/(m2·s),Ly峰 值 为10.06 μmol/(m2·s)和7.87 μmol/(m2·s);L、Ly的Pn均值分别为7.51、5.58 μmol/(m2·s)。
由图1c 可知,不同造林模式下,柠条Pn日变化均呈“单峰型”曲线,峰值均出现在12:00,N和Ny峰值分别为10.92 μmol/(m2·s)和12.73 μmol/(m2·s);N、Ny的Pn均值分别为4.37、6.86 μmol/(m2·s)。
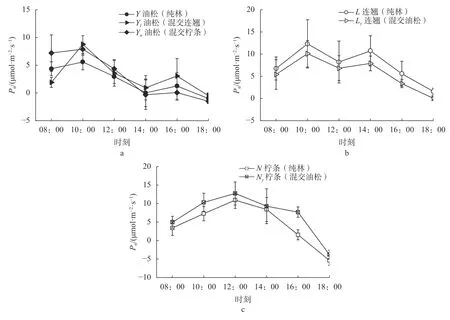
图1 不同造林模式下 3 个树种 Pn 日变化曲线Fig. 1 Diurnal Pn variation curves of 3 tree species under different afforestation modes
3.2 Pn 与影响因子的相关性
由表2 可知,所有测定树种Pn与气孔导度(Gs)、WUE、Ls等生理因子均表现出极显著正相关关系(P< 0.01),与胞间CO2浓度(Ci)呈极显著负相关(P< 0.01);仅Ly的Pn与饱和水汽压差(Vpd)呈极显著的正相关关系(P< 0.01),其余植物Pn与Vpd 显著性不强;Y、Yl、Yn的Pn与叶片温度(Tleaf)未表现出显著相关性,L、Ly、N、Ny的Pn与Tleaf均表现出极显著正相关关系(P< 0.01);
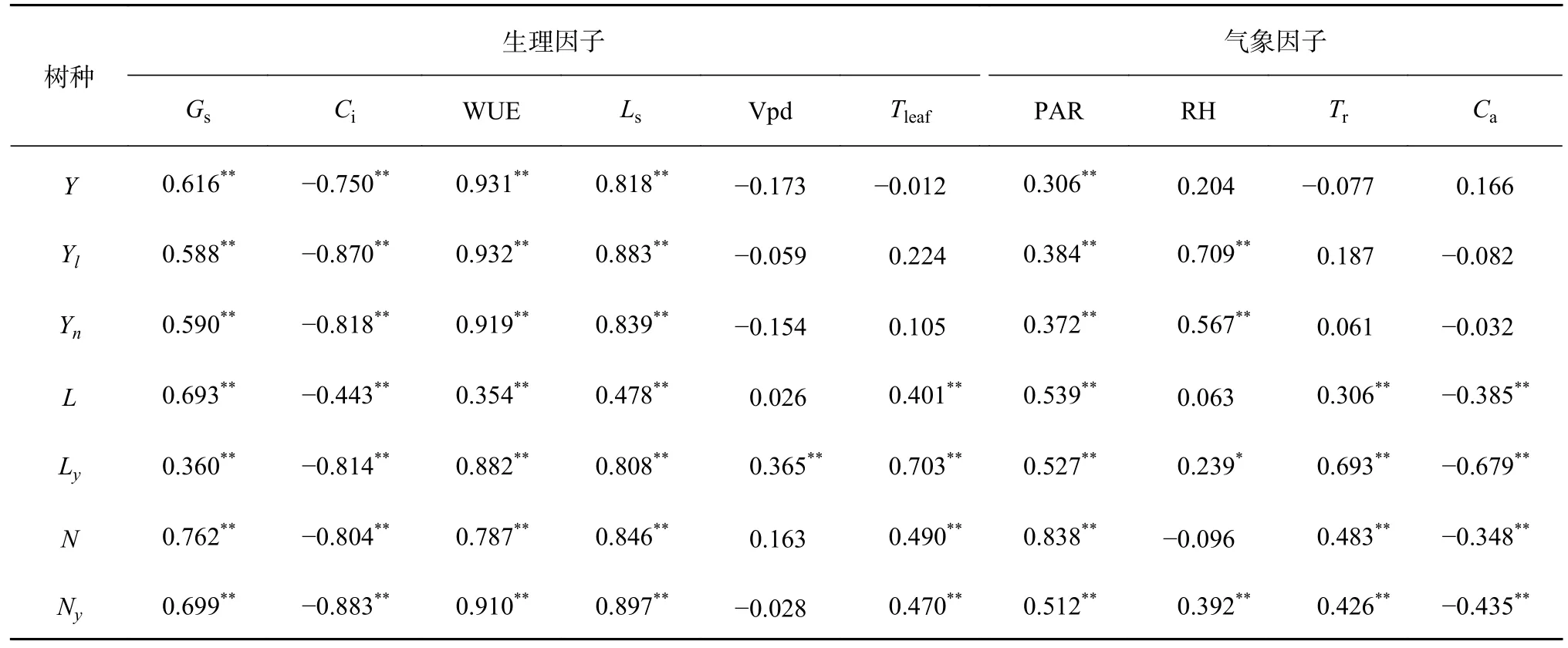
表2 Pn 与其影响因子的关系分析Table 2 Analysis of the relationship between Pn and its impact factors
所有测定树种Pn与气象因子相关性如表2 所示,均与PAR 均呈极显著正相关(P< 0.01);Y的Pn与RH 相关性不强,而Yl、Yn的Pn与RH表现出极显著正相关关系(P< 0.01),而L的Pn与RH 的相关性不强,Ly的Pn与RH 成显著正相关关系(P< 0.05),N的Pn与RH 相关性不强,而Ny的Pn与RH 呈极显著正相关(P< 0.01);L、Ly、N、Ny的Pn均与Tr呈极显著正相关关系(P< 0.01),与Ca呈极显著负相关(P< 0.01);Y、Yl、Yn的Pn与Tr、Ca相关性不强。
3.3 光响应过程
由图2 可知,不同造林模式下所有测定植物Pn随PAR 的变化趋势基本一致,当PAR < 200 μmol/(m2·s),纯林与混交林植物Pn均未显示出明显差异,油松、连翘和柠条的Pn分别迅速增加至3、8、4 μmol/(m2·s);在PAR > 200 μmol/(m2·s)的范围里,Pn上升逐渐减缓,最终趋于饱和,纯林与混交林植物Pn逐渐产生差异,当PAR 为2 000 μmol/(m2·s)时,Yl的Pn高 于Y42.1%,Yn的Pn高于Y10.5%(图2a);而Ly的Pn低于L40.4%(图2b);Ny的Pn高于N74.0%(图2c)。

图2 不同造林模式下 3 个树种光响应曲线Fig. 2 Light response curves of 3 tree species under different afforestation modes
3.4 光响应曲线拟合
4 种模型拟合的所有测定植物Φ均在0~0.125理论范围之内(表3),但RH、NRH 拟合的Pnmax均远高于实测值,拟合的LSP 均远低于实测值,EM 拟合的结果优于上述2 种模型,但拟合的LSP 仍远低于实测值,MRH 拟合的结果好于上述3 种模型。由于除MRH 外,其余模型的LSP均无法直接计算,因此在Pnmax和LSP 的估算中,采用MRH 模型能够较为准确地反映。另外,RH 与EM 模型拟合的所有油松的LCP 均远低于实测值,且RH 拟合连翘、柠条的LCP 均远高于实测值,EM 拟合的连翘、柠条的LCP 均远低于实测值。且EM 模型的Rd与实测值相比偏小,其他3 种模型拟合的Rd较接近实测值。在不同造林模式下除EM 模型拟合的油松决定系数(R2)相对较低外,其余3 个模型的R2均大于0.99(表3),拟合程度较高。
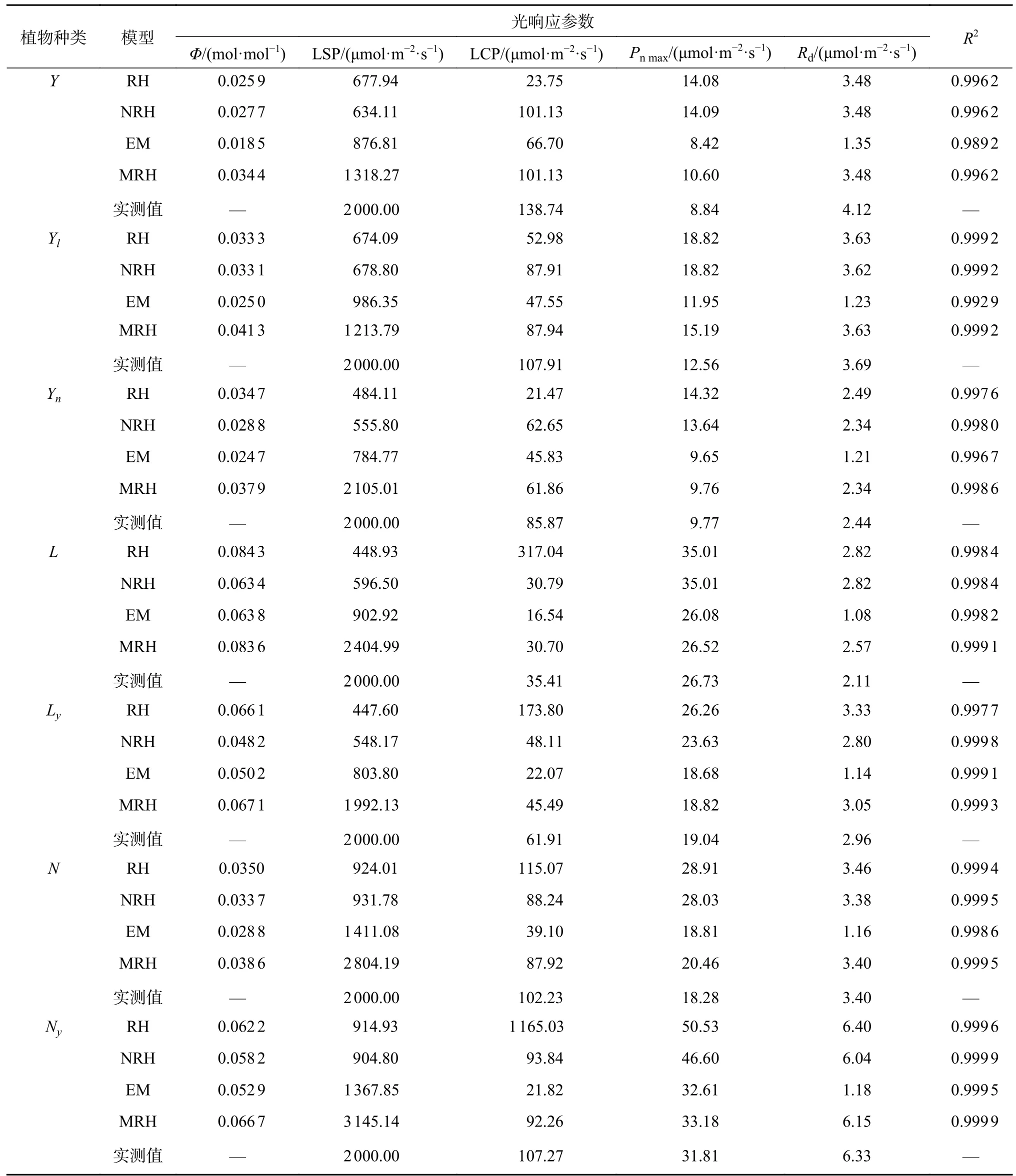
表3 光合参数的模型估计值与实测值Table 3 Model estimated and measured values of photosynthetic parameters
依据4 种模型的优劣性及对光合参数的求解精度不同,得出MRH 拟合效果最好,故采用MRH 求解的光合参数(Φ、LSP、LCP、Pnmax、Rd)进行植物的光合生理特性分析。
4 结论与讨论
4.1 讨论
4.1.1Pn日变化特征及其与影响因子的关系
Pn是植物生长过程中有机物积累的关键,对植物的生长和抗逆性非常重要[20-21],本研究中Y、Yl、Yn的Pn日变化均呈“双峰型”曲线,存在“光合午休”现象,这是植物长期进化过程中所形成的减少水分散失的方式[22];L、Ly的Pn日变化均呈“双峰型”曲线,这与王建华等[23]对4 月连翘生长初期的研究结果不同,这可能与本研究测定时间在7 月高温生长盛期有关。某些植物Pn日变化进程在夏季是双峰,而在5 月生长初期、10 月生长末期,单峰型居多[24];N、Ny的Pn日变化呈“单峰型”曲线,这是由于柠条可通过C4 光合关键酶转录与翻译水平上的积极响应来适应干旱环境,获得较高的Pn[25],邵玲玲等[26]的研究也表明柠条并没有“光合午休”现象。本研究结果表明,植物在不同造林模式下的Pn日变化趋势均未产生差异,仅Pn值的大小有所差异,这是由于在测定时间内,时间较短,内部生理因子相对稳定,Gs、WUE、Ls等生理因子与Pn均呈极显著的正相关关系也表明造林模式并未对植物Pn日变化趋势产生明显影响。
而纯林和混交林由于林内树种组成不同,生态环境则会产生差异[27],因此,外部环境就成了Pn的主要影响因素[28]。Yl和Yn的Pn日均值均高于Y,Ny的Pn日均值高于N,表明混交林油松、柠条的光合能力高于纯林;L的Pn日均值高于Ly,表明纯林连翘的光合能力高于混交林。本研究中Y、L、N的Pn与RH 相关性不强,而Ly的Pn与RH 呈显著正相关关系,Yl、Yn、Ny的Pn与RH 均表现出极显著正相关关系。这主要是由于混交有助于调节环境小气候,使得林内温度低,湿度大,而RH 大可以刺激气孔开放,增加CO2传输速率,加快植物光化学反应,促进光合作用[29]。因此,与纯林相比,油松混交林林内植物光合能力的变化主要与林内空气温、湿度有关。
4.1.2 光响应曲线特征
光响应曲线反映Pn随PAR 的变化趋势,是判定植物光合效率的重要方法[30]。本试验中3 个树种在低光强(PAR < 200 μmol/(m2·s))下,Pn迅速直线上升,处于光诱导期初期,在这一阶段,光合作用高速运转,Ci相对饱和,羧化限制等非气 孔 限 制 占 优;当PAR > 200 μmol/(m2·s)时,Pn增速减缓,进入光诱导后期,气孔限制已成为主要的限制因素,在这个阶段,虽然Gs增加,Pn也增加,但两者的增加速率不同步,CO2的吸收不能与光能的吸收相协调,光合电子的传递受到阻碍,并且Rubisco 羧化的活性降低,Pn增加缓慢[31]。有研究表明,植物的物质积累能力与Pn呈显著正相关关系(P< 0.05)[32]。本试验结果表明,在PAR > 200 μmol/(m2·s)后的各个光强梯度下,Yl、Yn的Pn均高于Y,Ny的Pn高于N,表明Yl、Yn物质积累能力均高于Y,Ny物质积累能力高于N,这与徐永兴[33]的研究结果一致。植物物质积累能力高,会对其叶片内其他生理过程如抗氧化酶活性、PSII 电子传递效率、叶绿素含量等产生积极影响,最终促进植物的生长[34]。
4.1.3 造林模式与光合参数的关系
Φ反映了植物光合作用的光能利用效率,尤其是对弱光( < 200 μmol/(m2·s))的利用能力[35],Φ高的植物,能在弱光环境下以最大能力利用低光量子密度,增强捕获光量子用于光合作用的能力,从而提高有机物的积累,提高其生存能力[36]。与纯林相比,Yl、Yn、Ny的Φ值分别提高了20.1%、10.2%、72.8%,弱光利用能力增强,这与李勇[37]对山西太岳山油松林人工林的研究结果一致,而Ly的Φ值降低了19.7%,弱光利用能力减弱,这可能与植物自身的生理特性及对环境的适应能力有关。
LCP 和LSP 的高低,反映了光合机构暗反应过程对同化力需求量的多少,是判断植物需光性的指标,代表植物对强光合弱光的利用能力[38]。一般来说,植物LCP、LSP 均高,则其适应强光的能力较强;植物LCP、LSP 均低,表明其具有很强的耐荫性;而植物LCP 低,LSP 高,表明适应多种光照环境能力较强[39]。本研究3 个树种对光照强度均表现出一定的适应性和可塑性。Yn与Y相比,LSP 高,LCP 低,说明Yn提升了对多种光照环境的适应能力;Ly与L相比,LSP 低,LCP高,表明Ly对光照的适应能力降低;Ny与N相比,LSP 高,LCP 高,Ny提升了其对强光的适应能力。植物光照生态幅比较中,Yn相较于Y提升了67.9%,Ny相较于N提升了12.4%,Yn与Ny均表现出显著较宽的光照生态幅,耐荫性和喜光性均增强[40],表明油松与柠条混交可以提升林内组成树种对光环境的适应性,这与谭淑娟等[41]研究结果一致。
Pnmax指在最适宜的环境条件下植物可表现的最大的净光合能力[42],本试验中不同造林模式下Pnmax大小次序为Ny>L>N>Ly>Yl>Y>Yn,相比与纯林,Yn的Pnmax降低了7.9%,而Ny的Pnmax提升了62.2%。说明Ny有助于提升其光合潜力,而Yn的Pnmax最低,这可能与油松叶片排列有关,针叶树种叶片密集排列相互遮荫可能会限制其叶束的Pn,这与吴旭等[6]的研究结果一致。Rd反映植物消耗光合产物的速率[43],是叶片活性的重要体现,本试验中植物Rd由高到低顺序依次为Ny>Yl>N>Y>Ly>L>Yn,与纯林相比,Ly、Ny的Rd分别升高了18.7%、80.9%,表明Ly、Ny叶片活性较高;Yn的Rd下降了32.8%,表明Yn通过降低叶片活性,减弱呼吸作用,减少对光合产物的消耗[44],以适应生态环境和促进干物质量的积累。因此,采用油松与柠条混交,可提升植物的光合能力及光照适应性,并促进植物生长,是人工林优化改造过程中切实可行的造林模式。
4.2 结论
与纯林相比,油松与连翘、柠条的2 种混交林中,植物Pn日变化趋势并未发生改变,仅Pn值的大小产生差异,油松为Yl>Yn>Y;连翘为L>Ly;柠条为Ny>N。纯林和混交林中植物Pn产生差异的原因主要是RH 不同,混交林中,Yl、Yn、Ny的Pn与RH 呈极显著正相关,纯林中Y、N的Pn与RH 相关性不强。与纯林相比,油松与柠条混交林中植物对弱光的利用性及对光强的适应性更好,其生存能力更强。
