河南洛阳马尾松树干液流昼夜变化特征及其影响因子分析
2023-03-30徐军亮章异平罗鹏飞师志强候佳玉翟乐鑫
竹 磊,徐军亮,章异平,罗鹏飞,师志强,候佳玉,翟乐鑫
(河南科技大学园艺与植物保护学院,河南 洛阳 471023)
树干液流是指在植物体蒸腾失水、根系吸收水分后,由茎输导组织运输至冠层的上升液流[1]。它是土壤-植物-大气连续体中的关键链接,决定了冠层的蒸腾量,反映了植物的水分运输状况以及植物对水分的利用特征[2]。正常情况下,植物的蒸腾耗水量与树干液流通量相等[3],因此,通过树干液流研究可掌握植物蒸腾耗水与需水规律。植物在白天和夜间均存在蒸腾作用,但在早期的研究中,许多学者认为夜间植物的气孔处于关闭状态以防止水分的流失[4],因此在液流计算时忽略了夜间液流[5],进而低估了植物的实际耗水量[6]。随着液流计的广泛应用,液流监测技术得到了很大的进步和发展,能够在植物自然生长的条件下连续测定其用水情况[7]。研究表明,夜间多数植物有部分气孔打开[8],植物存在夜间液流现象[9],且在全天液流通量中占有较高比例,可达全天液流量的5%~50%[10-12]。因此,开展植物夜间液流的生理生态学作用研究,有助于准确量化植物蒸腾耗水量乃至整个林分的蒸腾耗水量,提高对水资源管理、决策的科学性以及加深对植物液流及其影响因子关系的理解。
夜间液流的作用在不同树种中存在差异。Daley等[13]研究发现,纸桦(Betulapapyrifera)的夜间液流主要用于夜间蒸腾和茎干补水,同时可能还具有为纸桦边材深层细胞供氧的作用,而红枫(Acerrubrum)和红橡(Quercusrubra)的夜间液流则主要用于自身水分的补充及水分平衡的调节。王艳兵等[14]在对华北落叶松(Larixgmeliniivar.principis-rupprechtii)的研究中也发现,树木夜间虽存在微弱液流,但与饱和水汽压差和风速并不相关,因此推测华北落叶松的夜间液流不是树木冠层的蒸腾,而是被用作树干的水分补充。与之相反,Phillips等[15]在对花旗松(Pseudotsugamenziesii)的研究中则认为,夜间液流主要用于夜间的蒸腾。除树种差异影响外,由于生长环境的不同,影响夜间液流的主要环境因子也存在差异。陈立欣等[16]和孔喆等[17]认为风速和饱和水汽压差与夜间液流显著相关,Hayat等[18]认为饱和水汽压差、温度与夜间液流显著相关,而周翠鸣等[19]则认为夜间液流与环境因子之间的关系并不明显。由此可见,夜间液流的作用及环境因子对液流的影响存在差异。
马尾松(Pinusmassoniana)是我国亚热带东部地区乡土树种,资源丰富,种植面积较大,其分布极广,北自河南及山东南部,南至两广、台湾,东至沿海,西至四川中部及贵州,遍布华中华南各地[20]。目前,有关马尾松树干液流的研究主要集中在我国南方地区,在相对干旱的马尾松分布北界较少报道。与湿润区域相比,相对干旱生境下马尾松昼夜的液流变化特征及其影响因子尚不清楚。基于此,本研究在位于河南洛阳北亚热带向暖温带过渡的气候带,即秦岭东延的余脉山区,对马尾松树干液流进行连续监测,同时结合小型气象站对周围环境因子的监测,以期揭示马尾松在其分布北界树干液流的昼夜特征及其与环境因子的关系,探索马尾松蒸腾耗水规律,为马尾松分布北界地区的植被建设提供科学依据。
1 材料与方法
1.1 研究区概况
研究地点位于河南省洛阳市河南科技大学校园内(112°41′E,34°60′N),属于暖温带。洛阳市2019全年日照1 976.7 h,年平均温度为15.7 ℃,6—8月温度最高;年降水量为580.6 mm,降水主要集中在6—10月,占全年降水量的90.15%。
在研究区内马尾松分布较为集中且无人工喷灌干扰的区域选取3株干形较直、长势良好、健康无病害的马尾松作为试验监测对象,样树基本情况见表1。
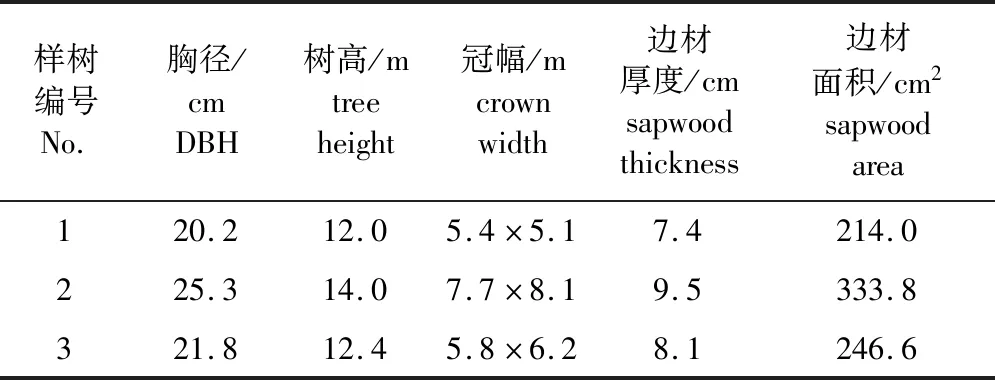
表1 样树基本特征Table 1 Information of sample trees
于2019年6—10月连续测定其树干液流速率。样地马尾松伴生树种有紫叶李(Prunuscerasifera)、石楠(Photiniaserratifolia)、圆柏(Sabinachinensis)、大叶黄杨(Euonymusjaponicus)和悬铃木(Platanusacerifolia)。
1.2 样树边材面积的确定
为避免对样树造成伤害,影响液流速率的监测,在样树附近另选12棵不同胸径的马尾松。在每棵树东西和南北两个方向用生长锥在胸径处钻取树芯,带回实验室进行固定和打磨后,通过颜色区分心边材,量取边材厚度(取东西、南北平均值)。将胸径和边材厚度进行拟合,得到马尾松胸径和边材厚度的线性拟合关系为:y=0.394 3x-0.522 2 (R2=0.97)。再根据边材厚度对样树胸径处边材面积进行计算。
1.3 树干液流的测定
在3株样树树干北面胸径处(距地约1.3 m),各安装1组SF-L型号的探针式液流计(TDP)和液流传感器(德国 Ecomatik 公司),用于监测马尾松树干液流;同时,探针与CR1000数据采集器(美国 Campbellsci 公司)相连,每10 min采集1次数据。
根据Granier[21]提出的热扩散探针间的温差与树干液流速率有密切关系的原理,利用Granier经验公式对马尾松树干液流速率进行计算:
(1)
式中:JS为树干液流速率,mL/(cm2·min);ΔTmax为每天最大的瞬时温差,℃;ΔT为上下探针的瞬时温差,℃。
树干累积日液流量为:
Q=∑(JS×AS×0.01)。
(2)
式中:Q为单位边材面积日总液流量,kg/d;JS为单位边材面积的平均液流速率,mL/(cm2·min);AS为边材面积,cm2;0.01为时间系数。
1.4 环境因子的测定
在样地内架设一台HOBO-U30便携式自动气象站(美国 ONSET 公司),对马尾松周边的环境因子进行同步监测,采集程序设置测量间隔为10 min,与液流计时间同步。主要监测指标有太阳辐射、空气温度、相对湿度和土壤含水量(土层深度10 cm与20 cm)。饱和水汽压差(vapor pressure deficit,VPD,式中记为pVPD)根据经验公式[22]计算:
(3)
式中:pVPD为饱和水汽压差,kPa;RRH为相对湿度,%;T为空气温度,℃。
1.5 数据处理
根据Daley等[13]对夜间液流时段的定义,将太阳辐射小于5 W/m2的时段定义为夜间,同时统计并区分出全天、白天和夜间的累积液流量,并按月取平均值。采用Pearson相关,分析日尺度(全天、白天和夜间)马尾松树干液流速率与环境因子的相关性。统计分析利用SPSS 25.0和Excel 2019完成,图表制作采用Orgin 2018完成。
2 结果与分析
2.1 马尾松树干液流量的昼夜变化
对研究区马尾松日液流量及夜间液流量测定结果见表2。由表2可以看出,6—10月马尾松的整日液流量均值为36.08 kg,夜间液流量为2.55 kg。整日液流量在6—10月整体呈先上升后下降的趋势,于7月达到最大值40.89 kg,10月达到最小值31.96 kg。与整日液流量的变化趋势相反,夜间液流量及夜间液流贡献率整体呈先下降后上升的趋势,于7月达到最小值(1.99 kg、4.87%),10月达到最大值(2.96 kg、9.27%),9—10月每小时平均夜间液流量为0.21~0.26 kg。零点前夜间液流量占总夜间液流量的83.42%~95.54%。
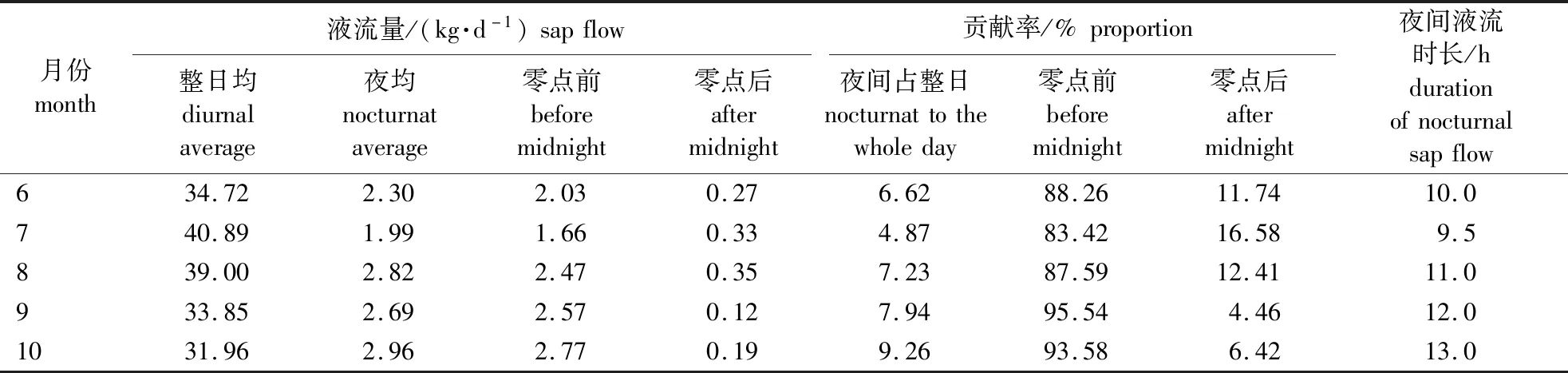
表2 马尾松各月日液流量及夜间液流量变化Table 2 Changes of diurnal and nocturnal sap flow of Pinus massoniana in different months
2.2 马尾松昼夜液流速率的变化
观察研究区不同月份马尾松树干液流速率变化特征(图1)可以看出,在日尺度上马尾松树干液流呈昼高夜低的单峰曲线。从6月开始,随着时间的推移,液流启动时间逐渐变晚,总体上从6:00—7:00开始。随着太阳辐射的加强和蒸腾作用的增大,液流速率逐渐上升,并于12:00—14:00达到峰值。之后随着太阳辐射的减弱,液流速率逐渐下降,当太阳辐射减弱到5.0 W/m2及以下时(夜间),液流速率下降放缓,趋近于零,直至第2天树干液流再次启动。
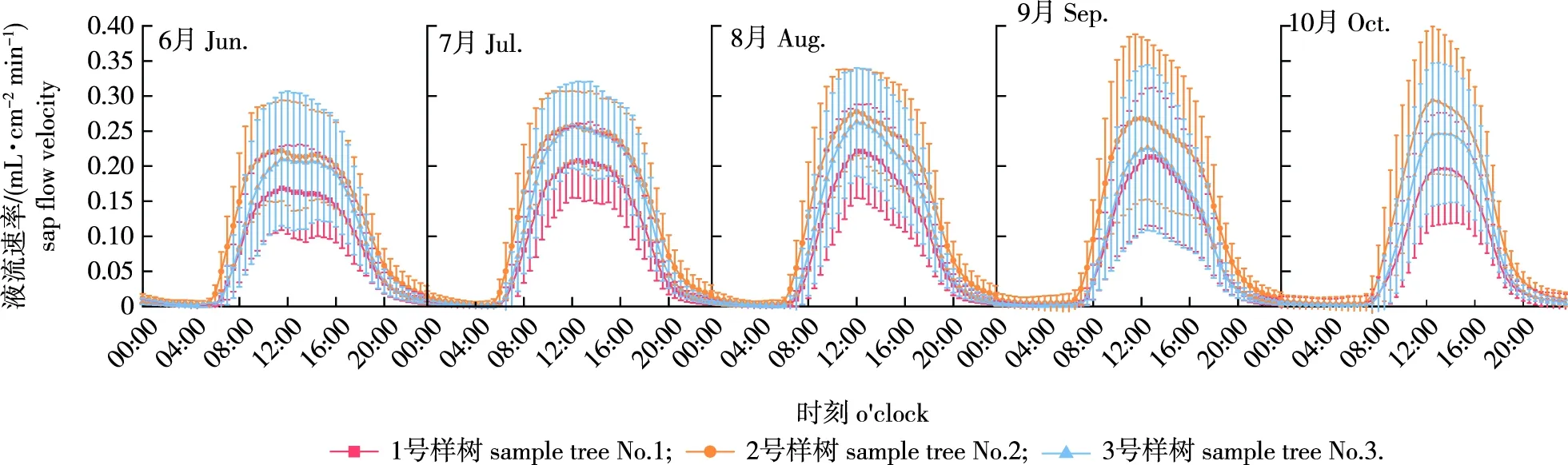
图1 马尾松6—10月树干液流速率日变化Fig.1 The diurnal variation of sap flow velocity of Pinus massoniana from June to October
在月尺度上树干液流速率曲线宽峰逐渐变窄,峰值变高。液流速率在7—10月呈上升的趋势,并在10月达到最大[0.294 mL/(cm2·min)]。虽然曲线中液流速率峰值逐月递增,但实际上各月平均液流速率大小依次排为:7月>8月>6月>9月>10月,其平均液流速率依次为0.105、0.100、0.088、0.085、0.080 mL/(cm2·min)。
而分析马尾松夜间树干液流速率(图2)的变化发现,不同月份马尾松夜间树干液流开始的时间(6—10月依次为19:30、20:00、19:00、18:30、18:00)和停止的时间(6—10月依次为05:30、05:30、06:00、06:30、07:00)与季节的变换有一定的关联。夏季夜间液流开始较晚、结束较早,秋季夜间液流开始较早、结束较晚。总体上看,夜间马尾松液流速率变化主要集中在零点前,占夜间液流速率变化的98.19%,零点后液流速率趋近于零且较为平缓。
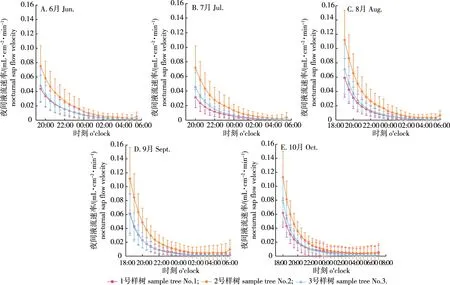
图2 马尾松6—10月夜间液流速率的变化Fig.2 The nocturnal sap flow velocity of P. massoniana from June to October
2.3 马尾松树干液流速率的影响因子
树干液流与环境因子关系密切,研究区6—10月环境因子的动态变化见图3。
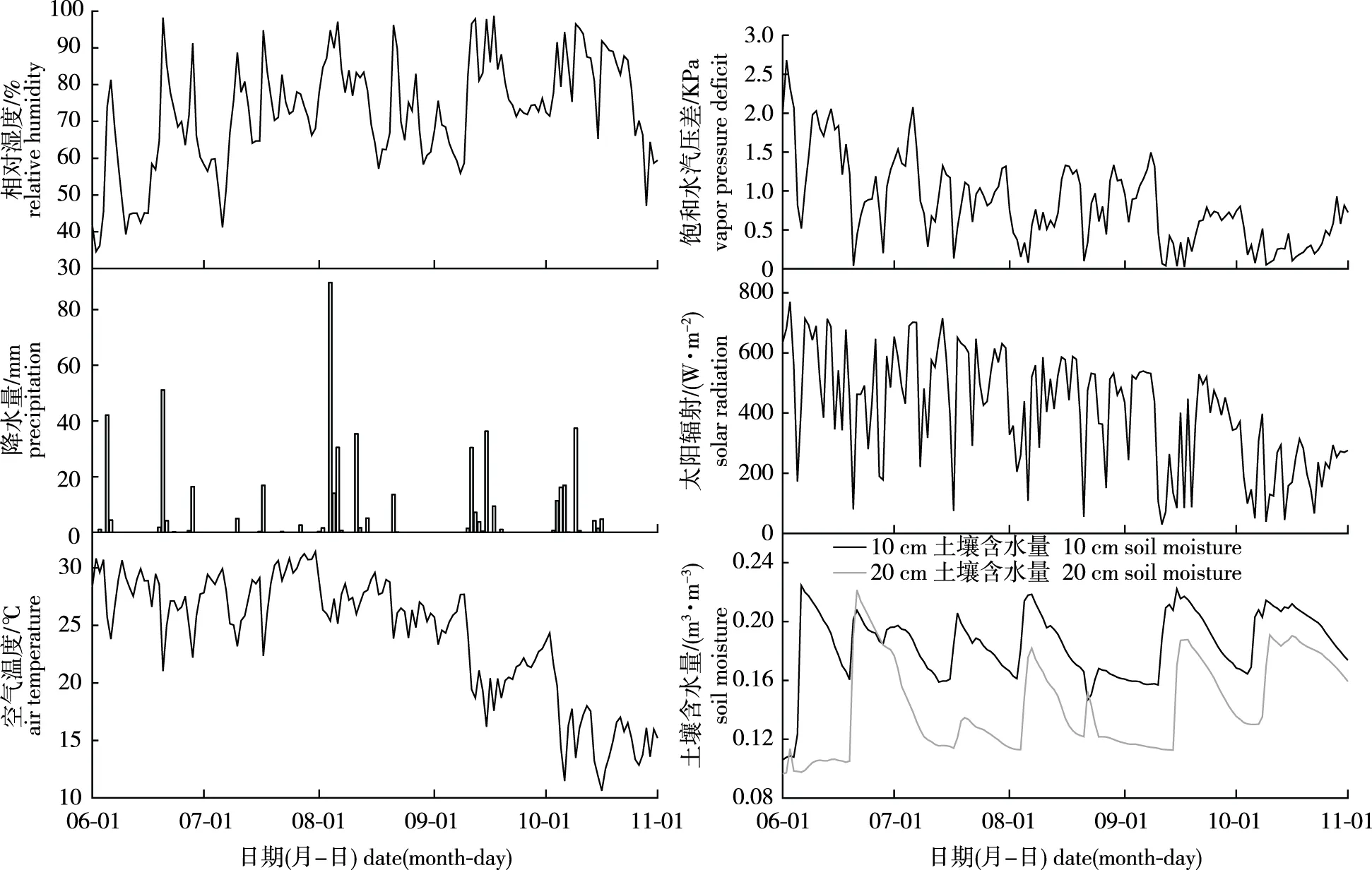
图3 研究区6—10月环境因子的动态变化Fig.3 Dynamic changes of environmental factors from June to October
从图3可以看出,6—10月环境日均相对湿度较为稳定,整体保持在72.54%左右;6—10月每个月都有降水事件发生,降水量大小顺序依次为8月(192.7 mm)>6月(121.8 mm)>10月(93.8 mm)>9月(90.0 mm)>7月(25.1 mm);日均空气温度、日均VPD和日均太阳辐射整体呈下降趋势,平均值分别为24.13 ℃、0.83 kPa和404.60 W/m2;10 cm和20 cm土壤含水量较为稳定,分别保持在0.18 m3/m3和0.14 m3/m3上下,当有降水发生时,10 cm和20 cm土壤含水量均呈上升趋势,随着土壤水分的蒸发,土壤含水量又呈下降趋势。
日尺度的线性回归分析结果表明(图4),3棵马尾松平均树干液流速率与饱和水汽压差、空气温度和太阳辐射呈极显著正相关关系(P<0.01),与相对湿度、降水量和风速呈显著负相关关系(P<0.05),按各指标相关系数绝对值顺序依次为太阳辐射(R2=0.567)>降水量(R2=0.270)>相对湿度(R2=0.233)>饱和水汽压差(R2=0.205)>空气温度(R2=0.162)>风速(R2=0.042)。
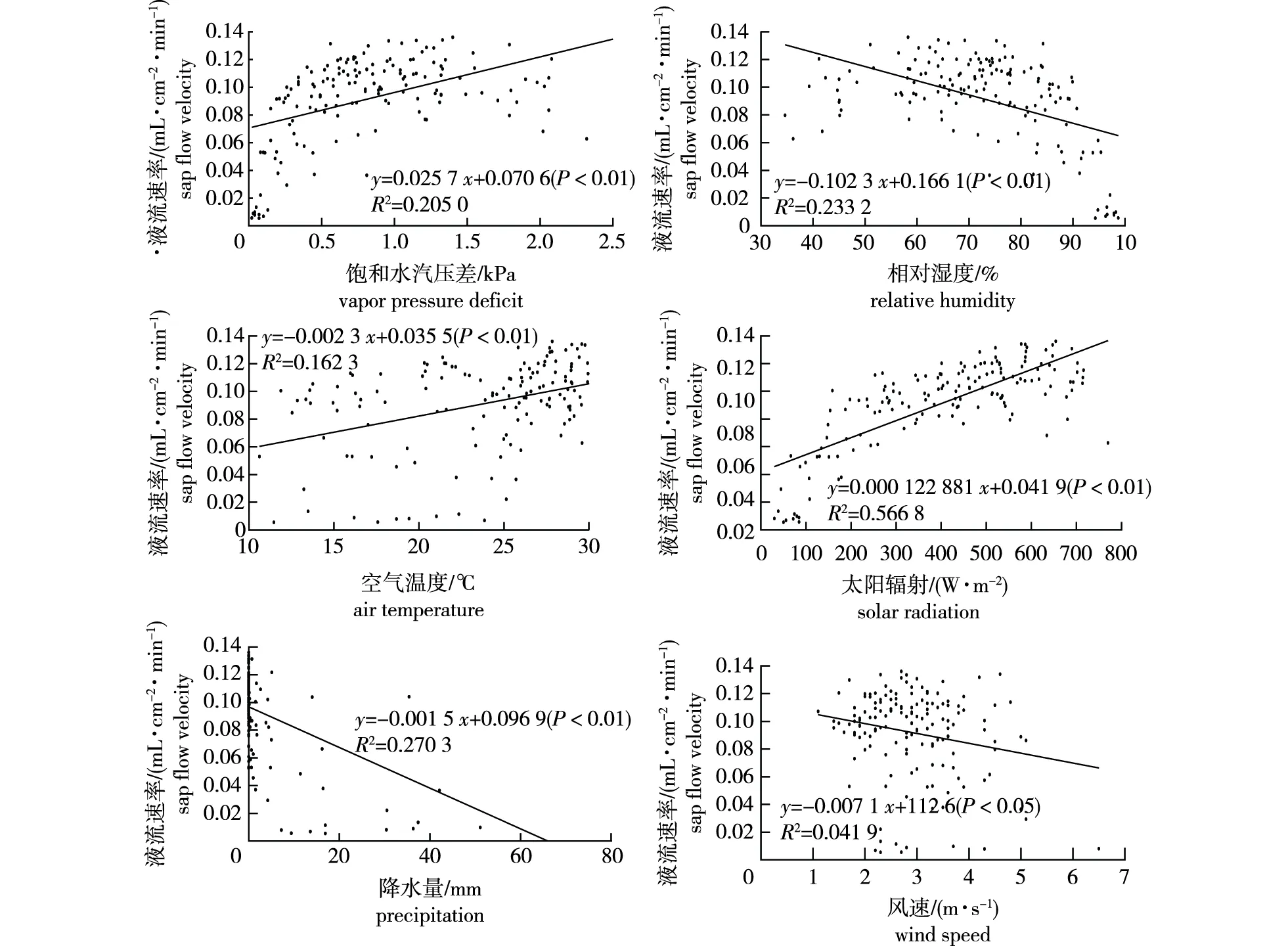
图4 马尾松液流速率与环境因子日均值的线性回归关系(N=153)Fig.4 Linear regression relationship between sap flow velocity and daily mean of environmental factors of P. massoniana(N=153)
马尾松日尺度树干液流速率与环境因子的Pearson相关系数见表3。由表3可以看出,在不同时间段内(全天、白天和夜间),太阳辐射是全天和白天时间段内液流速率的最大影响因子,VPD是夜间时间段内液流速率的最大影响因子。全天和白天时间段内,与树干液流速率相关系数较高的环境因子有太阳辐射、降水量、空气温度、相对湿度、风速和VPD(P<0.01)。与全天和白天相同,夜间液流速率与VPD、空气温度、相对湿度和降水量有显著的相关性(P<0.01),此外,与土壤含水量也有较高的相关性(P<0.05),而与太阳辐射和风速的相关性并不显著。可以看出,不同时间段内影响树干液流速率的环境因子有所不同。
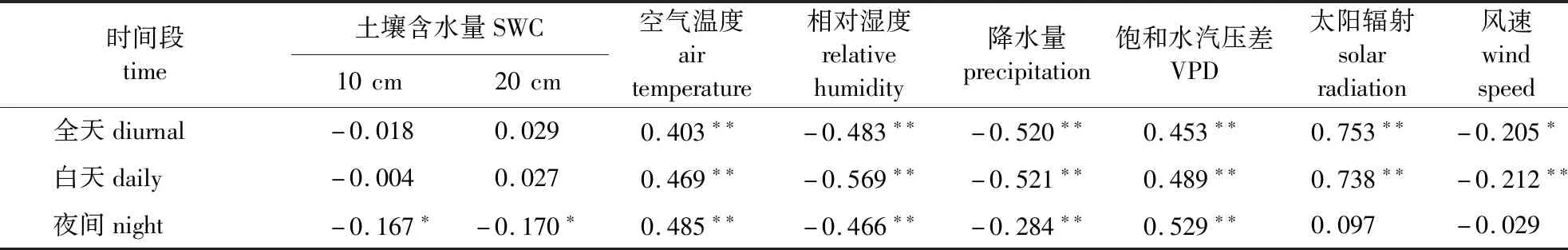
表3 马尾松日尺度树干液流速率与环境因子的Pearson相关系数Table 3 Pearson correlation coefficient between diurnal sap flow velocity and environmental factors of P. massoniana
3 讨 论
3.1 马尾松昼夜树干液流特征及树干液流量
马尾松树干液流昼夜特征呈昼高夜低的单峰曲线,与王立景等[23]、Xin等[24]和贾天宇等[25]的研究结果一致。树干液流特征均表现为在日出前后启动,随太阳辐射和蒸腾作用的增强而上升,午间前后达到最大值,随后又随着太阳辐射和蒸腾作用的减弱而下降,夜间表现为先迅速下降然后趋于平稳。俞新妥等[26]在对9种不同种源马尾松水分生理生态的研究中发现,马尾松树干液流速率峰值的季节变化规律为7—10月上升并于10月达到最大值,这与本研究结果基本一致。马尾松液流速率峰值虽然在6—10月逐月上升(图1),但各月平均液流速率结果显示,7月份液流速率最大[0.105 mL/(cm2·min)],10月份液流速率最小[0.080 mL/(cm2·min)],与涂洁等[27]的研究结果一致。液流速率日峰值和液流速率日平均值出现时间的差异性,可能是因为7月空气温度高,液流速率均值最大,但较少的降水量及过高的温度也会限制植物的蒸腾作用,使得植物体出现一定程度的水分亏缺,因此液流速率日平均值在7月达到最大值,而液流速率日峰值则出现在温度及水分相对适宜的10月。
不同月份,马尾松树干液流量与物候生长节律有关。从7月开始,马尾松树干平均日液流量逐月减少,波动范围为31.96~40.89 kg/d。这可能是因为马尾松形成层细胞的径向生长数量在7月达到峰值,此时马尾松处于快速生长期,生理活动活跃,日液流量较高。随后,形成层细胞分裂减缓、活动减慢,马尾松的生长逐渐变缓,日液流量逐渐降低。此外,随着季节的变化,饱和水汽压差和太阳辐射持续降低,树木生长需水量减少,冠层气孔导度减小,可能是导致马尾松平均日液流量逐月减少的另一个原因[28]。与平均日液流量相反,6—10月夜间液流量则呈逐月上升的趋势,波动范围为1.99~2.96 kg/d。这可能是因为6—10月每小时的平均夜间液流量变化不大(0.21~0.26 kg/h),而随着季节的变化,夜间时间变长,因此夜间液流量随着夜间时长的增加而增加。综上所述,在计算各月树木昼夜液流量时需要考虑树木的物候生长节律、环境因子的变化以及准确计算日间液流和夜间液流的时长等。
本研究区马尾松6—10月的平均日液流量为36.08 kg。程静等[29]在鼎湖山对不同径阶马尾松树干液流的研究中得出,马尾松湿季(7月)平均日蒸腾量为29.52 kg,干季(11月)平均日蒸腾量为20.91 kg;张璇[30]测得2016—2017年6—10月重庆缙云山马尾松单株平均日蒸腾量为36.68 kg;鲁小珍[31]在南京测得7年生(胸径10.2 cm)马尾松树干平均日液流量为9.33 kg。以上研究所在地区的年降水量在1 000~2 000 mm,远高于本研究区,但研究结果却较为相似,这可能是因为马尾松具有耐旱和高用水效率的特性[26]。
3.2 马尾松树干液流的环境影响因素
在马尾松的分布北界,树干液流主要受太阳辐射、VPD、温度、湿度和降水量的影响。目前有关马尾松液流与环境因子的相关研究,主要集中在我国南方地区,程静等[29]认为太阳辐射和VPD是控制鼎湖山马尾松树干液流的主要驱动因子;张璇等[32]在缙云山马尾松树干液流的研究中也认为气象因子对马尾松树干液流有较大影响,按影响程度由强到弱依次为太阳辐射、VPD、风速、湿度和温度,这与本研究的结果基本一致。这是因为太阳辐射决定了叶片气孔开放的大小[2],VPD的增加可以使叶片气孔导度增大,加快叶片表面的水汽交换[17],树木蒸腾随着太阳辐射和饱和水汽压差的增强而增强,是影响树干液流的主要限制因子。此外,树干液流不仅受到环境因子的影响,还受到自身生物学特性和土壤供水水平的影响,如于萌萌等[33]认为不同树种木质部解剖结构的差异及树木自身的生理特性会导致植物的水分消耗能力不同,树种间的树干液流可能存在差异;吴旭等[34]发现刺槐(Robiniapseudoacacia)在降水前气孔为不完全开放状态,蒸腾受到抑制,树干导水能力低,而当降水发生后,土壤水分得到暂时的补充,树干液流得到较大改善。由此可以看出,环境及植物自身生理特性的不同,对环境变化的响应也有所不同。
不同时段内,马尾松树干液流主要限制因素不同。白天马尾松树干液流主要受太阳辐射、降水量、VPD、温度、湿度和风速的影响,其中太阳辐射为最大影响因子;夜间主要受VPD、温度、湿度、土壤含水量和降水量的影响,VPD为最大影响因子。白天树干液流与风速极显著相关(P<0.01),在夜间却相关性不强。魏潇等[35]认为夜间液流与风速相关性不显著,而张婕等[36]和Phillips等[37]则认为风速与树干夜间液流显著相关。这可能是因为风速对树干液流的影响是较为复杂的,风速的增加可以加快树木蒸腾,但风速过大时会导致气孔导度降低,甚至关闭[38]。无论白天还是夜间,VPD、温度、湿度和降水量均对树干液流有较大影响,这可能是由于本研究地区降水较少(年均降水量为580.6 mm),当有降水发生时,树干液流与降水有极显著的相关性(P<0.01),液流活动较为活跃[16]。因此,由于研究地区的不同,导致研究结果存在差异。
总体看,我国中西部相对干旱地区为马尾松分布的北界,这一地区的马尾松树干液流呈昼高夜低的单峰曲线,6—10月的平均液流量为36.08 kg/d,夜均液流量为2.55 kg,每小时平均夜间液流量为0.21~0.26 kg,夜间液流量占整日液流量的4.87%~9.27%;零点前液流量占夜间液流量的83.42%~95.54%。研究区马尾松主要受太阳辐射、VPD、温度、湿度和降水量的影响,白天树干液流的主要影响因子是太阳辐射,夜间树干液流的主要影响因子是VPD、温度和湿度。因此,马尾松存在夜间液流,并占有一定的比例,在液流计算时不能忽略其夜间液流。
