云南省文山市鸟类多样性及垂直分布特征
2023-03-29王艳梅范仕祥张文文胡远芳
王艳梅,范仕祥,张文文,,曾 卫,胡远芳,崔 鹏,罗 旭①
(1.西南林业大学云南省森林灾害预警与控制重点实验室/ 西南林业大学生物多样性保护学院,云南 昆明 650224;2.生态环境部南京环境科学研究所/ 国家环境保护武夷山生态环境科学观测研究站/ 国家环境保护生物安全重点实验室,江苏 南京 210042;3.国家林业和草原局西南调查规划院,云南 昆明 650216)
优先区域或热点区域识别是实施生物多样性保护计划的首要步骤,其重要目的之一是为了将有限的保护资源分配到最值得保护的区域,以达到最佳保护效果[1-3]。国家生态环境部门根据种类的丰富和稀有濒危情况、生态系统类型的代表性以及区域的不可替代性而划定35个保护优先区域,这是我国生物多样性保护的重点和关键区域。保护好优先区域,对于保护生物多样性、维护生态平衡和实现人与自然和谐,具有十分重要的意义[4]。西双版纳生物多样性保护优先区域是35个保护优先区域之一,位于云南省南部,总面积为42 585 km2,涉及云南省17个县级行政区,包括6个国家级自然保护区[4]。
文山市位于云南省东南部,是西双版纳保护优先区域涉及县域之一。该区域是我国典型喀斯特地貌的核心地带,发育了喀斯特山地特有植被,保存了许多特有物种,生物多样性保护价值和地位极高[5]。鸟类作为生物多样性的重要组成部分,对环境变化相对敏感且分布广泛,因而常作为环境监测和评价的指示类群[6]。滇东南鸟类调查始于1956年中苏科考队在云南南部进行的鸟类调查,但调查范围未涉及文山市[7-8]。文山市境内有文山国家级自然保护区老君山片区,有关文山市鸟类资源的数据主要源于文山国家级自然保护区的综合科学考察报告[9],但该报告未区分西畴县小桥沟片区和文山市老君山片区的鸟类,因此文山市保护区内和保护区外的鸟类组成、多样性数据目前较为缺乏。
在当前全球气候变化背景下,生物物种多样性沿环境梯度的分布模式及其机理研究受到生物地理学家和生态学家的高度关注[10]。物种垂直分布主要有3种模式,即单调递增、单调递减和中峰模式[10]。研究不同地区尤其是生物多样性热点地区和空白地区鸟类多样性垂直分布格局,是生物多样性保护的关键。目前,国内鸟类垂直分布研究多在西南地区开展,结果均较好地支持中峰分布模式假说,即在中海拔多样性最高[11-13]。同时,随着季节变化,鸟类会发生垂直移动,即动物沿着海拔的季节性重复运动[14]。这种行为允许动物利用不同的环境来满足其季节性变化的需求,或遵循沿海拔分布的季节性变化[15]。相对于纬度移动,海拔移动是一种低成本、高效益的运动,因为它的迁移距离较短,需要的生理和形态适应更少[15]。高黎贡山季节性垂直分布研究结果表明:随着季节变化,鸟类丰富度峰值会发生改变,鸟类存在垂直移动现象,因此,就海拔跨度大的自然保护区而言,在制定鸟类保护措施时有必要考虑鸟类的季节性垂直变化[16]。该研究拟解决如下科学问题:(1)获取文山市鸟类物种组成本底数据;(2)明确文山市保护区内外鸟类物种组成和多样性差异;(3)揭示老君山保护区鸟类垂直分布和垂直移动的季节性变化。通过该研究以期为文山市林业、生态环境和自然保护区相关管理部门提供本底数据和管理依据。
1 研究区域
文山市位于云南省东南部(23°06′~23°44′ N,103°43′~104°27′ E),总面积为2 977 km2,属滇东南岩溶地貌。北回归线横穿文山市境,文山市属中亚热带季风气候区,大部地区冬无严寒,夏无酷暑,春秋长,冬夏短,年平均降水量为1 187.8 mm[17]。文山国家级自然保护区老君山片区位于文山市西南部(小桥沟片区位于西畴县),面积为229.6 km2,占保护区总面积的85.46%,该区域海拔落差较大,最低海拔为1 385 m,最高海拔为2 991.2 m[9]。
文山市大部分区域地势较为平缓,农田和村庄是主要景观,与之不同的是老君山山体独立,海拔落差大,森林覆盖度高。因此,将研究区域划分为自然保护区内、外两个调查单元。
2 研究方法
2.1 样线布设
依据《县域鸟类多样性调查与评估技术规定》(环境保护部2017年第84号公告)规定的科学性、全面性、重点性和可达性原则[4],采用样线法进行鸟类调查,在两个调查单元采用不同的样线设置方法。在保护区外,依据生态环境部提供的全国10 km×10 km网格,通过全国地理信息资源目录服务系统(https:∥www.webmap.cn/.)下载云南省文山市行政边界,设置文山市鸟类调查网格38个(不包含保护区),每个网格中至少设置1条样线,共41条(图1)。在保护区内,按网格设置样线难以充分体现山地丰富的鸟类多样性,因此以300 m为间隔将老君山从山底(1 400 m)到山顶(2 900 m)划分为1 400~1 700、>1 700~2 000、>2 000~2 300、>2 300~2 600 和>2 600~2 900 m 5个海拔带。每个海拔带设置2~5条样线,长度从1.5~ 2.5 km不等,共18条样线。
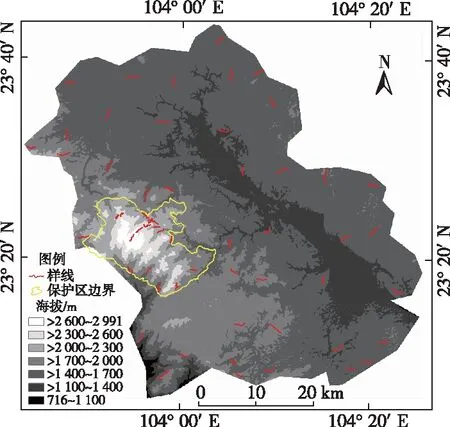
图1 云南省文山市鸟类调查样线分布
2.2 野外调查
野外调查于2019—2020年开展,在繁殖季和非繁殖季各调查2次,繁殖季调查时间为2019年7月和2020年4月,非繁殖季调查时间为2019年9—10月和2020年1月。每次开展野外调查时,保护区内的18条样线均调查。在保护区外,因所涉面积较大且存在山体滑坡、道路施工等突发情况,故每次调查根据交通可达性选取10~15条样线开展调查,直至所设41条样线均完成单次调查。在8:00—12:00 和15:30—19:00鸟类活动频繁的时间段进行调查,调查行走速度一般控制在1.5~2.5 km·h-1,借助Olympus 8×42 双筒望远镜观察,记录鸟类种类和数量,能明确辨别叫声的鸟类也予记录。调查时携带《中国鸟类野外手册》[18]、Canon7D 相机和400 mm长焦镜头辅助辨认鸟种。
2.3 数据分析
鸟类编目的分类系统、鸟类居留型依据《中国鸟类分类与分布名录》[19]。区系划分参照《中国动物地理》[20]。保护级别和濒危等级界定参考2021版《国家重点保护野生动物名录》[21]和《中国生物多样性红色名录:脊椎动物 第二卷 鸟类》[22]。
鸟类食性数据参考《中国鸟类的生活史和生态学特征数据集》[23],由于鸟类食性复杂,森林鸟类根据食性可分为食果鸟、食蜜鸟、食谷鸟、食虫鸟、食肉鸟和杂食鸟等类群[24]。在分析数据时将食果鸟、食谷鸟和食蜜鸟等均归为植食性鸟类;将食虫鸟和食肉鸟等均归为肉食性鸟类。在进行老君山鸟类垂直分布特征分析时,仅分析繁殖鸟类(即留鸟和夏候鸟),排除不能确定海拔归属的鸟类(包括猛禽、雨燕科和燕科等长时间在空中活动的类群)[16],并采用插值法研究海拔鸟类物种丰富度:即认为鸟类在它的观测最高海拔和最低海拔之间是连续分布的[13],用于制作鸟类垂直分布柱状图。在进行垂直移动分析时,为了减少结果的误差,还排除了两个季节调查中记录总次数小于5的物种。
采用R语言中 “iNEXT”包基于样本大小的稀疏和外推(R/E)方法计算文山市和老君山两个季节物种丰富度,并且计算样本覆盖率(sample coverage,SC),以衡量样本完整性[25]。
通过非度量多维尺度分析(nonmetric multidimensional scaling,NMDS)方法分析保护区内外鸟类群落组成差异,基于Bray-Curtis相似性系数构建不同季节保护区内外相似性矩阵,采用胁强系数(Stress)度量二维NMDS的拟合优度。Stress<0.05表明拟合程度极高,0.05≤Stress<0.1表明拟合程度较高,0.1≤Stress<0.2表明拟合程度一般,0.2≤Stress<0.3表明拟合程度略差,Stress≥0.3表明拟合程度较差[26]。非参数多元方差分析也称为基于置换的多元方差分析 (permutation based MANOVA,PerMANOVA)是一种利用置换方法检验多元方差分析的非参数程序,以距离测量为基础,适用于生态学群落分析[27]。笔者研究中将PerMANOVA用于检验不同季节保护区内外鸟类群落组成是否存在差异。NMDS和PerMANOVA均采用R语言中 “vegan”包完成。
对保护区内外物种数、个体数、遇见率和Shannon-Wiener指数采用非参数Mann-Whitney检验,视P≤0.05为显著。采用Excel 2019、R 4.0.5和GraphPad 8.0软件对所有数据进行处理。
采用Shannon-Wiener多样性指数(H′)和Pielou均匀度指数(J)计算群落多样性,公式[28]如下:
H′=∑PilnPi,
(1)
J=(-∑PilnPi)/lnS,
(2)
Pi=Ni/N。
(3)
式(1)~(3)中,Pi为群落第i个物种个体数占群落中总个体数的比例;Ni为第i个物种个体数;S为群落中物种数;N为群落个体总数。
采用Jaccard相似性指数(I)表示不同海拔群落相似性,计算公式[29]为
I=c/(a+b-c)。
(4)
式(4)中,c为群落A和群落B共同的物种数;a和b分别为群落A和群落B物种数。
样线鸟类遇见率计算公式为
E=N/L。
(5)
式(5)中,E为鸟类遇见率;N为全部物种总个体数量;L为样线长度,km。
3 结果与分析
繁殖季和非繁殖季保护区内外物种稀疏和外推曲线见图2,曲线趋于平缓,SC值均大于0.98,表明抽样充分,调查较为全面。
3.1 文山市鸟类组成
野外记录到鸟类199种,隶属于16目51科(附录1),其中雀形目鸟类145种,占调查总数的72.86%;非雀形目鸟类54种,占调查总数的27.14%。各科中记录种数较多的是鹟科(Muscicapidae)和柳莺科(Phylloscopidae)。
记录到国家Ⅱ级重点保护鸟类有棕背田鸡(Zaporniabicolor)、褐翅鸦鹃(Centropussinensis)、黑翅鸢(Elanuscaeruleus)、凤头蜂鹰(Pernisptilorhyncus)和凤头鹰(Accipitertrivirgatus)等24种。《中国生物多样性红色名录:脊椎动物 第二卷 鸟类》中收录白腹隼雕(Aquilafasciata)1种易危(vulnerable,VU);近危(near threatened, NT)14种,包括中华鹧鸪(Francolinuspintadeanus)、短嘴金丝燕(Aerodramusbrevirostris)和画眉(Garrulaxcanorus)等。
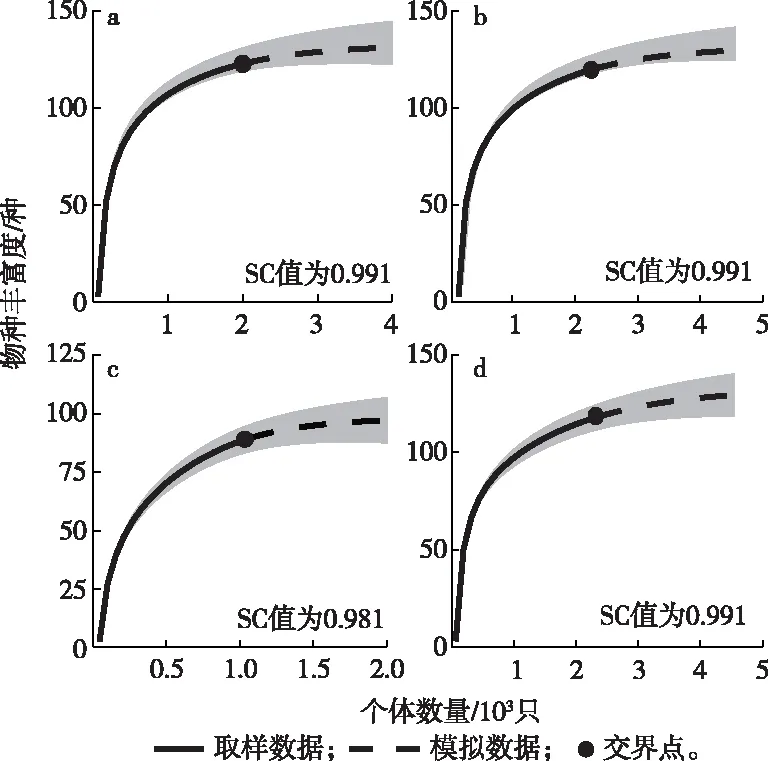
a为保护区内繁殖季;b为保护区内非繁殖季;c为保护区外繁殖季;d为保护区外非繁殖季。
从居留类型分析来看,留鸟152种,占调查总数的76.38%;夏候鸟14种,冬侯鸟26种,旅鸟7种,留鸟占绝对优势。对166种繁殖鸟(留鸟和夏候鸟)进行区系分析:东洋种占绝对优势(113种),广布种次之(50种),古北种最少(3种)。
3.2 文山市保护区内外鸟类多样性比较
保护区内调查到鸟类149种,隶属于12目45科,个体数为4 109只;记录国家Ⅱ级重点保护鸟类16种,红色名录收录近危(NT)7种;其中繁殖季调查到鸟类121种,隶属于12目43科;非繁殖季调查到鸟类118种,隶属于9目37科。在保护区外,调查到鸟类147种,隶属于16目47科,个体数3 269只;记录国家Ⅱ级重点保护鸟类15种;红色名录收录易危物种(VU)1种,收录近危(NT)物种10种;其中繁殖季调查到鸟类87种,隶属于10目36科;非繁殖季调查到鸟类115种,隶属于14目41科。文山市24种保护鸟类中,保护区内记录16种,占66.67%;濒危物种15种,其中保护区记录7种,占46.67%。
对保护区内外鸟类的物种数、遇见率、Shannon-Wiener指数和Pielou均匀度指数进行繁殖季和非繁殖季的差异性检验(图3)。在繁殖季,保护区内鸟类Pielou均匀度指数高于保护区外(P=0.028,Z=-2.191),其余各指数无显著差异。在非繁殖季,保护区外物种数(P=0.002,Z=-3.165)和Shannon-Wiener指数(P=0.005,Z=-2.776)显著高于保护区内,其余指数无显著差异。
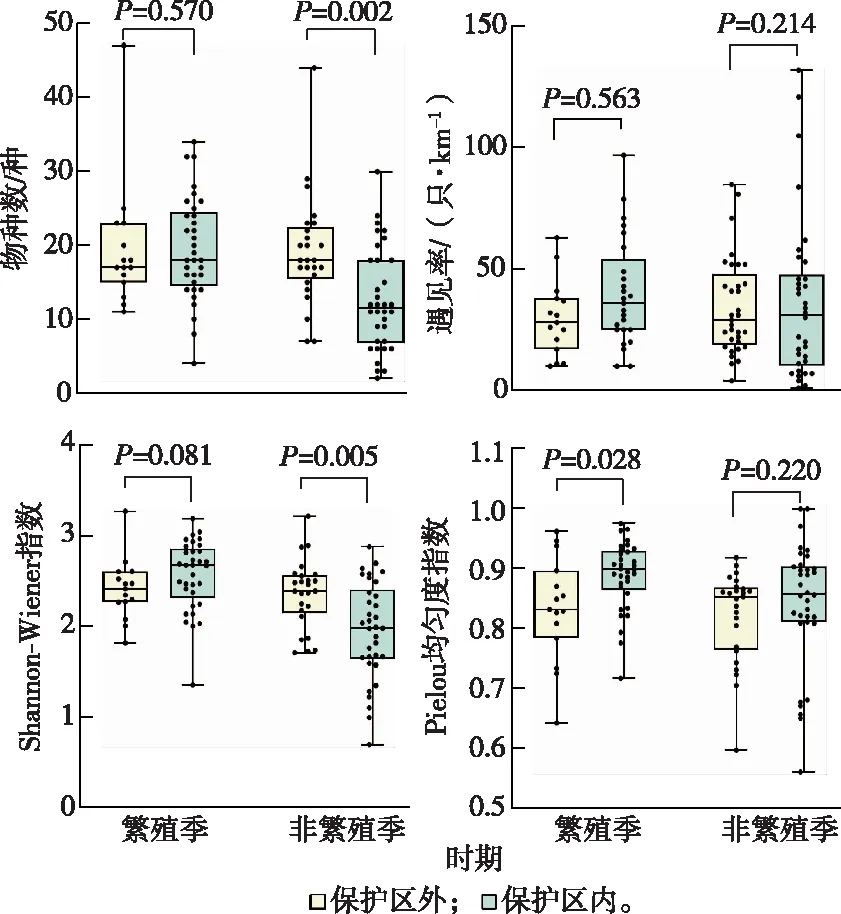
图3 保护区内外鸟类群落特征比较
通过NMDS分析,在繁殖季、非繁殖季和全年的胁强系数均小于0.2,说明NMDS分析具有一定拟合效果。由NMDS结果(图4)可知,保护区内外鸟类群落有重叠区域,重叠部分表明鸟类群落相似性高,未重叠部分表明鸟类群落相似性较低。PerMANOVA检验结果表明,老君山自然保护区内外鸟类群落水平存在显著差异(繁殖季:R2=0.120,P=0.001;非繁殖季:R2=0.052,P=0.001;全年:R2=0.120,P=0.001)。
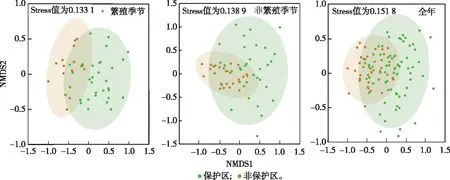
绿色的点表示保护区内每条样线的鸟类群落,橘红色的点表示保护区外每条样线的鸟类群落。点与点之间的距离越近表示群落的物种相似性越高,距离越远表示相似性越低。
3.3 文山市老君山鸟类垂直分布
3.3.1鸟类垂直分布特征
老君山繁殖季和非繁殖季鸟类丰富度都为中峰模式,即随着海拔的增加,鸟类丰富度呈现先增加后减小趋势,在>1 700~2 000 m海拔带鸟类丰富度最高,在>2 600~2 900 m海拔带鸟类丰富度最低,但繁殖季鸟类丰富度整体高于非繁殖季(图5)。

图5 老君山繁殖鸟类垂直分布
相似性分析结果表明最高海拔和最低海拔鸟类相似性最小,相邻海拔鸟类相似性较高。在繁殖季,>2 000~2 300和>2 300~2 600 m海拔带鸟类最相似;在非繁殖季,>1 700~2 000和>2 000~2 300 m海拔带鸟类最相似(表1)。
3.3.2留鸟垂直分布的季节变化
老君山繁殖季和非繁殖季共调查到留鸟85种,排除不能确定具体海拔鸟类后有73种,排除记录次数小于5的物种后有55种。这55种鸟类中,有8种鸟类的分布范围未发生季节变化,其余鸟类从繁殖季到非繁殖季均发生海拔分布的变化,有4种变化方式(图6)。第1种是分布下移,此类季节性移动的鸟类繁殖季分布上限可达到较高海拔带,在非繁殖季分布上限向下移动,共记录到31种鸟类,占55种鸟类的56.36%,比如白喉红臀鹎(Pycnonotusaurigaster)和白颊噪鹛(Garrulaxsannio)。第2种是分布上移,鸟类在繁殖季分布于较低海拔带,非繁殖季上移,记录7种,占14.55%,比如方尾鹟(Culicicapaceylonensis)和栗臀币鸟(Sittanagaensis)。第3种是分布收缩,即繁殖季分布于较广的海拔范围,非繁殖季海拔下限上移、海拔上限下移,记录7种,占12.73%,比如黑头奇鹛(Heterophasiadesgodinsi)和红尾希鹛(Minlaignotincta)。第4种是分布扩张,与分布收缩相反,非繁殖季鸟类垂直分布范围变大,记录2种,占3.64%,分别是大山雀(Paruscinereus)和红翅鵙鹛(Pteruthiusaeralatus)。
老君山鸟类季节性垂直移动主要为分布下移,所记录到31种分布下移的鸟类中,杂食性鸟类17种,占54.84%;肉食性鸟类13种,占41.94%;植食性鸟类1种,占3.23%。

表1 老君山不同海拔鸟类群落的相似性指数和共同物种数比较

图6 老君山鸟类垂直分布的季节变化
4 讨论与建议
4.1 文山市鸟类多样性
此次调查记录到文山市鸟类199种,占云南省鸟类945种[30]的21.06%,占中国鸟类1 445种[19]的13.77%,补充了西双版纳优先区文山市鸟类多样性本底数据,同时提供了老君山鸟类多样性基础数据。文山国家级自然保护区的综合考察包含了文山市老君山片区和西畴县的小桥沟片区,报告记录221种鸟类(含文献数据),但未区分物种记录的片区[9]。老君山调查到鸟类149种,在保护区原有科考名录上新增鸟类53种,其中新增国家Ⅱ级保护鸟类7种。
根据动物地理区系的划分,老君山位于东洋界华南区滇南山地亚区[16],在此次调查结果中,东洋界鸟类占绝对优势,古北界占比最小,这与文山国家级自然保护区科考报告中数据一致。物种的丰富度和特有性是评价生物多样性保护优先区保护价值的指标之一[31],近年来相关工作不断推进,西畴县距离文山市较近,也属于西双版纳生物多样性保护优先区。金吉辉等[32]利用红外相机监测技术对西畴县进行鸟兽调查,其中鸟类调查到12种,鸟类中雉类4种;文山市调查到鸟类199种,但雉类仅3种。红外相机可以更好地用于监测雉类和地栖鸟类,采用样线法则能调查到更多的小型雀形目鸟类,样线法和红外相机技术均存在局限性[33],建议文山市后期可以增加红外相机监测法,对地面活动的鸟类和兽类进行监测。
4.2 文山市老君山鸟类垂直分布
研究山地生态系统群落的组成和物种分布范围的时空变化对于了解山地生物生态和保护状况至关重要[34],对于横断山区鸟类垂直分布的研究多是针对繁殖季开展[11-13],而关于垂直分布随着季节变化的研究目前较为缺乏[16],笔者研究为山地鸟类垂直分布的季节变化增加了文山市老君山的案例。老君山鸟类丰富度峰值位于中低海拔(>1 700~2 000 m),属于中峰模式中的偏前峰模式,这与针对横断山区多个山体的研究结果[11-13]相似。老君山高海拔(>2 600~2 900 m)地区温度较低,生境多为苔藓矮林,食物相对缺乏;低海拔(1 400~1 700 m)地区多为村庄、农田和人工林,生境破坏严重,干扰较大,这可能是老君山鸟类丰富度最大值位于中低海拔的主要原因。
从繁殖季到非繁殖季,老君山56.36%的鸟类呈现分布下移现象,且主要表现为海拔分布上限的下移,这与高黎贡山鸟类在冬季分布上下限均下移有所不同[16],可能是因为在非繁殖季节老君山天气恶劣和食物供应减少导致高海拔地区不适合栖息,而低海拔地区缺少森林等适宜栖息地,鸟类难以下移,从而形成分布上限下移而下限不变的情形。老君山向下移动的鸟类主要由杂食性鸟类和食虫性鸟类组成,这可能有两个原因:一是杂食性鸟类可以充分利用不同海拔的食物资源,因此会进行季节移动[35];二是节肢动物在秋冬季呈减少趋势,尤其是在高海拔地区[36]。
4.3 文山市鸟类保护

文山国家级自然保护区老君山片区繁殖季和非繁殖季鸟类丰富度峰值都位于>1 700~ 2 000 m海拔带(图5),然而根据保护区功能区的划分[9],>1 700~2 000 m多处于实验区和缓冲区,调查过程中时常发现有采集野生菌和放牧的情况,因此建议保护区管理机构可将>1700~2 000 m海拔带作为鸟类监测保护的重点区域,减少人为干扰和破坏。在非繁殖季保护区外重点物种和区域也值得关注,例如2020年1月14日,笔者在文山市暮底河水库调查到15只鸳鸯(Aixgalericulata),它是国家Ⅱ级重点保护野生鸟类,在云南为冬候鸟[16]。鸳鸯越冬生境特征及栖息地选择研究结果表明,人为干扰会影响鸳鸯的越冬活动及停歇地的选择[40]。暮底河水库是文山市饮用水水源地,现暂不能确定该水库是否为鸳鸯的稳定越冬地,建议当地环境保护部门和水库管理部门加强对鸳鸯种群的监测和栖息地保护。
致谢:感谢云南文山国家级自然保护区管护局蓝俊麟和杨云协助调查工作;感谢李奇生、李江梅、聂军、田静和洪磊参与野外调查;感谢李斌强、李万德和梁丹在数据分析方面提供的帮助。
