植物根内皮层分化及其可塑性研究进展
2023-03-14朱莹莹孙光军刘海伟
张 标 ,朱莹莹,孙光军,刘海伟
(1. 中国农业科学院 烟草研究所/农业农村部烟草生物学与加工重点实验室,山东 青岛 266101;2. 中国农业科学院 研究生院,北京 100081;3.中国烟草总公司 贵州省公司,贵州 贵阳 550004)
植物根系可以选择性地从土壤环境中摄取水分和矿质养分,是植物从土壤中吸收养分的重要器官,在植物的生长发育过程中起着至关重要的作用。植物根系可以通过调节自身结构以适应土壤复杂多变的环境[1],如干旱、涝渍,以及养分胁迫等,甚至与微生物的相互作用密切相关[2-4]。植物根系吸收的水分和矿质养分向中柱的径向运输主要包括3 条途径,分别是通过相邻细胞细胞间隙自由扩散的质外体运输途径、通过相邻细胞胞间连丝的共质体运输途径,以及通过细胞膜上的转运蛋白介导的跨细胞运输途径[5]。植物根系吸收的水分和养分需穿过所有细胞层才能到达根中央维管系统,并通过木质部导管运输到地上部,以供植物生长发育所需[6-8]。
根内皮层是根部细胞层最内层的细胞层,是控制植物对水分和矿质养分吸收的关键细胞层。在很多物种中,内皮层被人们认为是水分和养分通过径向运输至植物根维管系统的基本屏障[9]。研究表明,植物对水分和矿质养分吸收的过程中,根内皮层在发挥着核心作用。因为根内皮层在发育过程中形成了对水分和矿质养分吸收的屏障,从而可以有选择性地从土壤中摄取水分和矿质养分[10]。内皮层的发育起始于根尖分生组织。在根分生组织中,皮层初始细胞的斜周分裂导致内皮层的形成[10],这个过程受到转录因子SHR(SHORTROOT)、SCR(SCARECROW),以及生长素等途径共同调控[11],并且内皮层细胞壁中充满疏水聚合物,因此根内皮层被认为是水和溶质从皮层向维管系统运输的主要限制因素[12]。近年来,调控内皮层分化的分子机制和内皮层屏障在植物根部物质运输中的关键作用是当前研究的热点和重点内容。因此,本文就内皮层分化、内皮层屏障作用及其在应对环境变化的可塑性展开综述,以期为该领域的科研工作者提供有益的认识和参考。
1 内皮层分化
内皮层细胞的形成靠近根分生组织的静止中心,与皮层—内皮层初始细胞的斜周分裂关系密切。这种分裂形成的内皮层作为最内层的皮质细胞层,包围着根的中央维管系统[13]。根据皮层细胞初生壁和次生壁未分化、出现不连续的凯氏带和不完整的木栓层,以及连续的凯氏带和完整的木栓层的先后顺序,将内皮层的发育分为未分化阶段、分化阶段Ⅰ和分化阶段Ⅱ(图1)。
根内皮层处于分化阶段Ⅰ时,此阶段可形成凯氏带,但不形成木栓层。随着植物根系的发育,根内皮层分化在阶段Ⅰ向阶段Ⅱ转换过程中,木栓质以零散的、类似“补丁”的形式在内皮层细胞开始沉积[14],在内皮层细胞中可以观察到“补丁状”不完整的木栓层。最后,在完全木栓化的内皮层细胞中,木栓质覆盖整个内皮层细胞的表面。因此,在完全分化的内皮层细胞中会形成由不同的聚合物组成的2 种结构,即凯氏带和木栓层(图1)。
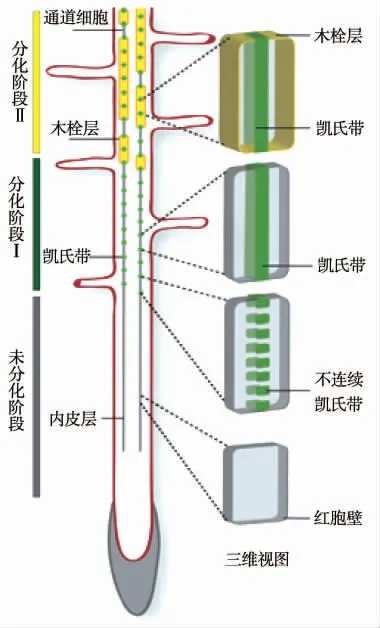
图1 内皮层分化示意图(改自Barberon and Geldner,2014)[15]
1.1 凯氏带
凯氏带主要由疏水的木质素聚合物组成,是位于相邻内皮层细胞之间细胞壁上的木质化结构[10]。这种疏水聚合物通过嵌入内皮层细胞壁从而封闭了相邻内皮层细胞间的细胞间隙,在根内皮层细胞中起着质外体扩散屏障的作用[16-17]。
在过去相当长一段时间内,人们提出木栓质是凯氏带的主要成分,但是现在这种说法已经被推翻。有研究指出,凯氏带实际上是主要以木质素为基础的结构[14]。这一见解使我们能够清楚地定义由不同聚合物组成的2 种扩散屏障结构,即基于木质素沉积形成的凯氏带以及相继形成的由木栓质沉积组成的次生壁结构木栓层[14]。
对拟南芥(Arabidopsis thaliana)的研究表明,内皮层从未分化阶段向分化阶段Ⅰ状态转变主要是由转录因子MYB36 驱动的[18],MYB36 直接调控与凯氏带形成的相关基因如凯氏带结构域蛋白(CASP)、过氧化物酶(PER64),以及类渗透蛋白(ESB1)的正向表达。在相应的MYB36 缺失突变体的研究中并未定位到凯氏带,相反,而是观察到木栓质的异位沉积,且木栓质沉积更靠近根尖分生组织以响应凯氏带的缺失[18]。对MYB36 功能缺失突变体的研究中发现,木质化的丧失导致内皮层透性增加,从而导致矿质养分的动态平衡被破坏[19]。这些证据直观表明了MYB36 在凯氏带形成过程中的决定性作用。植物根系吸收的水分和养分一旦到达内皮层,凯氏带就会封闭质外体,迫使养分运输通过共质体途径运输以保护维管系统免受营养物质的外流和病原菌的入侵[20]。
1.2 木栓层
内皮层分化阶段Ⅱ的特征是木栓层的形成,这是以木栓质为基础沉积在次生壁的次生细胞壁结构,随着植物根系的发育,最终形成完整的木栓层覆盖整个内皮层细胞表面。木栓质是由多种单体组成的复杂的生物聚合物,其沉积对环境具有高度响应性[21]。木栓质在控制水和溶质运输方面的作用主要依赖于其脂肪族结构,脂肪族木栓质结构域的典型单体是ω-羟基酸、α,ω-二元酸、脂肪酸和醇等,甘油被酯化为ω-羟基酸、α,ω-二元酸,这些单体通过质膜运输,最终组装成质外体中的木栓质聚合物[21,22]。
木栓质的沉积可能与转录因子MYB41 有关。有研究表明,MYB41 的异位表达会诱导内皮层木栓化,但是它在内皮层发育中的确切作用目前尚不清楚[23]。木栓质的生物合成涉及多种酶,主要包括3-磷酸甘油酰基转移酶(GPAT)家族、细胞色素P450单加氧酶(CYP)家族、脂肪酰基还原酶(FAR)、羟基肉桂辅酶A 转移酶(ASFT),以及β-酮酯酰-辅酶A合成酶(KCS)等[24]。虽然学者们对木栓质的单体组成研究已有几十年,但目前关于木栓质的生物合成的许多过程仍不是很清楚[25]。
2 内皮层屏障
研究指出,根内皮层细胞在发育过程中会经历两个特殊的分化阶段,分别形成凯氏带和木栓层两种重要的扩散屏障(图2),二者能够选择性的控制水分以及养分等进入根部细胞,在控制和调节水分和养分的获取和流失方面起着重要作用[7,10,16]。
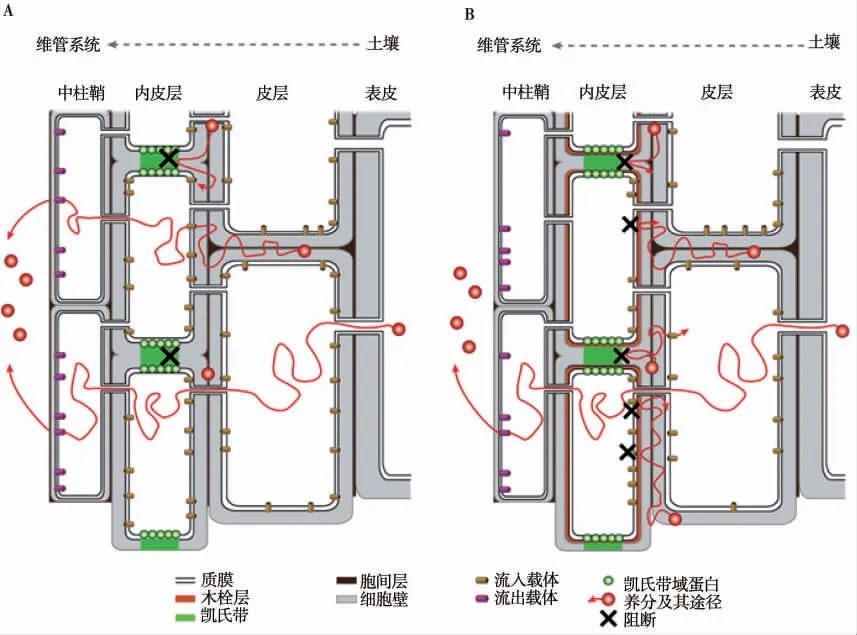
图2 凯氏带和木栓层在限制离子通过内皮层运输方面的作用示意图(改自Geldner,2013)[26]
2.1 凯氏带屏障作用
基于细胞壁的扩散屏障的形成在植物中建立有效的运输途径在植物吸收营养物质的过程中起着至关重要的作用。最典型的例子是根内皮层中作为质外体屏障的凯氏带的建立[27](图2-A)。凯氏带类似于生理“栅栏”,在大多数的根中,水分和养分从根的皮层到木质部的质外体途径运输被凯氏带阻断[28-29],同时有利于防止有毒物质进入植物体[30]。例如,Cd被认为优先通过质外体途径向木质部维管系统运输,根内皮层细胞木质化形成的凯氏带有利于限制这种重金属通过质外体途径运输,从而减少根和地上部对其的吸收和积累[31]。
在玉米(Zea mays)的一项研究中显示,ZmSTL1基因编码定位于凯氏带结构域的重要转录蛋白家族ZmESBL,它通过调节内皮层凯氏带结构域的木质素沉积来调节内皮层凯氏带的形成。ZmESBL 的缺失直接导致内皮层凯氏带结构域的木质素沉积受损,从而导致凯氏带的形成被破坏。缺乏ZmESBL的突变体表现出凯氏带结构域木质素沉积受损,导致凯氏带屏障缺陷,在盐胁迫下表现为显著增加了Na+通过质外体途径的运输,进而增加了Na+通过蒸腾流从根到地上部的运输,从而导致植株表现为盐敏感型[32]。该研究还发现ZmESBL 的同源基因ESBL也参与了拟南芥凯氏带的发育及其耐盐性的调节,揭示了基于木质素的凯氏带显著抑制了植株地上部对Na+的积累,并且显著提高了植株在高蒸腾环境条件下的耐盐性。
2.2 木栓层屏障作用
木栓层作为继凯氏带形成之后的另一个重要的屏障,阻断了养分的跨细胞运输途径,也标志着内皮层细胞完全分化,此时养分的运输只能通过共质体途径进行(图2-B)。木栓质作为内皮层屏障的疏水性成分,在非生物胁迫以及生物胁迫下减轻对植物的伤害起着关键作用[3,33-34]。在非生物胁迫的背景下,木栓质的功能得到了广泛的研究[21,35-39]。
木栓质含量的改变可能对植物的生理及生长发育等产生重大的影响。例如,在拟南芥的研究中,对木栓化增强型突变体esb1 的具体分析不仅为木栓质在水分和溶质的运输中起到的作用提供了确凿的证据,而且还表明根中更高的木栓质含量对整个植物生理功能的影响,如减少干旱胁迫下的植株失水、枯萎,以及不同的地上部离子组成等[6]。
值得一提的是,根内皮层细胞中的质外体屏障并不能完全阻断水和溶质向中柱的运输。因为内皮层中部分细胞并没有木栓化,这些细胞被人们称为“通道细胞”(图1),它们为水和溶质的运输提供了新的通道[40]。有研究指出,通道细胞的形成可能与细胞分裂素(Cytokinin,CTK)有关[41]。因此,内皮层中的质外体屏障并不是完美的屏障[5]。
植物在其生长发育过程中会进化出多种适应机制来响应环境的变化,如转运蛋白就是根系调节物质吸收的重要机制,但在细胞间非特异性的质外体运输途径中的物质吸收调节也不容忽视。根内皮层中的凯氏带和木栓层作为质外体屏障是调控质外体运输的关键机制,在控制物质的流入、流出,以及抵御病原菌入侵等方面起着重要作用[16]。
3 内皮层分化的可塑性
虽然植物在正常生长发育过程中根内皮层会经历正常的分化阶段,但从实际情况来看,植物在生长发育过程中难免会遭受各种逆境胁迫,对植物生长发育产生不利影响,导致内皮层细胞早期分化或延迟分化(图3-A),这种生理机制是植物为适应环境变化而形成的一种自我保护机制。因此,内皮层的发育进程受到广泛的非生物胁迫和生物胁迫的影响,这种影响使得内皮层细胞的分化具有高度的可塑性,并且这种可塑性可以最大限度地减轻各种胁迫对植物的伤害作用。
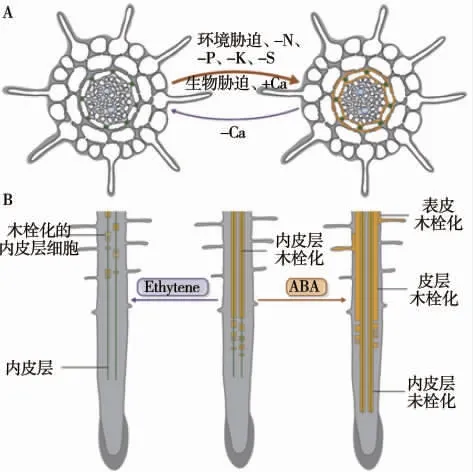
图3 内皮层分化的可塑性示意图(改自Barberon,2017)[10]
3.1 环境胁迫对内皮层分化的影响
诸多的环境胁迫,如盐分、干旱,以及缺氧等胁迫下均会诱导凯氏带的早期发育、木栓质的合成,及其在内皮层中的沉积增加[42-47]。以干旱胁迫为例,Grunhofer 等[48]对杨树(Populus)进行模拟干旱胁迫处理,结果发现其根系发育表现出高度的一致性。具体表现为典型的凯氏带早期发育和随后的内皮层木栓化程度不断增加,根尖部分的根系导水率比其他区域高得多。分析表明,根基部的木栓质沉积增加导致根系导水率显著降低[48-49]。在大麦(Hordeum vulgare)的研究中也有类似的发现,通过PEG8000 处理模拟不同渗透势的水分亏缺显著上调了根中木栓质生物合成基因的表达,导致内皮层木栓化增加,并显著降低了质外体途径的根系导水率,防止水分从根内部向外的回流散失导致失水[50]。对水稻(Oryza sativa)的研究表明,干旱胁迫下显著增加了内皮层细胞的大小和厚度,并且导致木栓质更多的在内皮层积累,内皮层木质化和木栓化增加[51]。从本质上来说,增强的木质化和木栓化的作用是在干旱期间保存根维管系统中的水分,以保护根不会在水分亏缺下死亡[51]。在葡萄(Vitis vinifera)的研究中也有同样的结论,即干旱胁迫下,木栓质生物合成相关基因MYB41 的表达上调,导致木栓质沉积增加以响应水分亏缺[34]。因此,这些环境胁迫因素可以直接影响内皮层的发育进程,表现为促进内皮层分化,而更早的形成内皮层屏障可以减轻逆境胁迫对植物的伤害。
通过对植物受盐分胁迫的深入研究发现,盐分胁迫会影响根中凯氏带的宽度,并在多种植物中诱导内皮层细胞的木栓化[52]。在水稻的几项研究中发现,盐分通过根的径向运输受到凯氏带和木栓层的限制[42-43,53];在渗透胁迫下水稻根内皮层木质化和木栓化程度增加,并且减少了根内部组织的失水[54];水稻根内皮层分化形成的质外体屏障在其适应受涝条件、防止径向氧气的损失等方面起着至关重要的作用[55],这些研究为内皮层的屏障作用提供了直接的证据。
有研究表明,拟南芥在盐分胁迫下,木栓质通过限制Na+的内流、K+的外流,以及水分向介质中的回流来增强植株的耐盐性[33]。在干旱、盐分或其他非生物胁迫下,木栓质在根内皮层细胞壁中的沉积也会发生变化,以防止水分和养分的流失、增强根的保水能力。例如,对拟南芥的研究发现,在干旱和盐分胁迫下,木栓质含量及其沉积形成的木栓层在减少水分损失和减少根对Na+的吸收方面起着关键作用[56]。木栓质还会影响植物对矿质养分的吸收和运输,进而影响到营养的有效性,起到水分和养分扩散屏障的作用[16]。木栓质在根中的产生和沉积对植物的生长发育、生理、与微生物的协调作用,以及各种胁迫适应具有显著的影响,并且在土壤有机质周转、土壤化学组分、水分,以及其他与微生物群落相关的因素方面具有潜在的生态影响[57]。
3.2 矿质元素对内皮层分化的影响
矿质元素在影响植物根内皮层分化过程中起着重要作用。一些必需营养元素如氮(N)、磷(P)、钾(K)或硫(S)的缺乏同样影响根部内皮层的分化[16]。Armand 等[58]对大麦的研究表明,低N 和低P 供应不仅会导致主根凯氏带和木栓层的形成显著增加,而且还会导致侧根的凯氏带和木栓层的形成显著增加。这与Li 等[59]研究结果一致。在低N 和低P 处理下质外体屏障形成增加,尤其是木栓层形成增加导致了根系导水率显著降低。当重新加入营养充足的基质后,低N 和低P 处理的植株根系导水率迅速恢复。这可能表明,营养胁迫下质外体屏障的形成导致根系导水率的降低在一定程度上是可逆的[16,58]。深入分析表明,增加的凯氏带和木栓层不仅能控制植物对水分和养分的吸收,而且可以阻止水分和养分的流失,水分和养分得以保留在植物体内以供植物生长发育所需,彰显出内皮层屏障作为双向屏障的重要作用[10,58]。但之前在水稻的一项研究中显示,在低浓度铵态氮肥供应条件下,木质素和木栓质的沉积较少,根系导水率和溶质渗透率与对照相比更大,而在高浓度铵态氮肥供应条件下生长的植株则相反[60]。这种差异可能与植物种类差异以及自身遗传因素有关。
另外,Siqueira-Silva 等[61]在研究去除根尖内皮层的质外体屏障对植物吸收铁(Fe)的过程中发现,植株地上部积累了更多的Fe,增加了Fe 的毒性效应,直接证明了根内皮层屏障在限制Fe 从根部到地上部分的运输方面所起的重要作用。
重金属Cd 胁迫可诱导小麦(Triticum aestivum)、水稻等作物的根内皮层质外体屏障早期发育,并且Cd 处理浓度越大,质外体屏障发育越强,凯氏带和木栓层的发育更靠近根尖。这可以减少小麦、水稻等作物的根对Cd 的吸收和向地上部的转运积累[62-63]。近几年人们发现,工程纳米颗粒物对环境的污染日益严重。对玉米的研究发现,氧化铈(CeO2)纳米颗粒物与Cd 处理均会导致根尖附近质外体屏障发育的增加,与对照相比,CeO2纳米颗粒物与Cd 处理条件下质外体屏障形成更靠近根尖[64]。相似的研究如在氧化镧(La2O3)纳米颗粒物处理下会促进与木质素合成相关基因表达上调,诱导玉米幼根质外体屏障早期发育,并降低玉米根系导水率[65]。Liska 等[66]在对玉米的根进行单侧干旱胁迫和单侧Cd 胁迫的研究中发现,单侧干旱胁迫以及Cd 胁迫会诱导根内皮层中凯氏带和木栓层的早期发育及不对称发育。其结果表明,植物受到胁迫之时内皮层屏障在保护维管系统以及在控制水和离子向地上部运输中的重要作用。
另外,一些研究表明,一些微量元素,如钙(Ca)、硅(Si)等,可能与Cd2+或Na+之间存在相互作用,在Cd 或盐胁迫下外源添加Ca 或Si 会影响内皮层的分化。例如,在研究Ca 含量对东南景天(Sedum alfredii)吸收Cd 的影响中发现Ca 作为常量元素与根内皮层中凯氏带的形成和木栓质的沉积有关。在低Ca 供应条件下会诱导根内皮层质外体屏障发育延缓,从而促进东南景天对Cd 的吸收和积累。分析表明,低Ca 供应条件下,木质素和木栓质合成相关基因的表达降低,延缓了内皮层中木质素和木栓质的形成与沉积,从而延缓了质外体屏障的形成和发育。而高Ca 供应条件下质外体屏障作用增强,说明Ca 含量与根内皮层中凯氏带和木栓层的发育呈正相关[67]。Si 可以通过诱导内皮层质外体屏障的形成增加减少了盐胁迫下水稻幼苗对Na+的吸收[68]。在重金属Cd 胁迫下,添加Si 显著增加了水稻[69]和小麦[70]根内皮层木栓化,从而降低了通过质外体途径对Cd的吸收。然而,另有研究表明,渗透胁迫可以促进大麦根内皮层木栓化,但在渗透胁迫下添加Si 并不能促进大麦根内皮层木栓化。渗透胁迫下Si 处理后植株抗性增强是由其他因素引起的[71]。因此,有学者认为Si 对内皮层分化和质外体屏障发育的潜在影响仍存在争议[71]。因此,从某种程度上来说,在重金属、盐分或其他胁迫下,内皮层发育进程被改变是因为Ca 或Si 与Cd 或Na 之间存在一定的相互作用,从而影响环境胁迫下内皮层的分化。
3.3 植物激素对内皮层分化的影响
Barberon 等[16]发现,在很多营养胁迫条件下,根内皮层木栓化是高度可塑的,这些营养胁迫通过植物激素脱落酸(abscisic acid,ABA)和乙烯(ethylene)对木栓质沉积产生影响,并且ABA 和乙烯在影响内皮层分化方面起到拮抗作用(图3-B)。ABA 通过增加苯丙氨酸解氨酶、过氧化物酶的活性,以及木栓质相关基因的表达来调控质外体屏障的发育[72]。在Cd胁迫下,非超积累生态型植物东南景天内源ABA 水平由于ABA 生物合成基因的表达上调而增加,而超积累生态型植物中内源ABA 水平并无变化。同时,非超积累生态型植物内皮层中凯氏带和木栓层的发育比超积累生态型植物更靠近根尖[73]。研究表明,乙烯是影响质外体屏障形成的另一种重要激素。高水平的外源乙烯应用会推迟超积累生态型东南景天质外体屏障的形成[74],促进其对Cd 的吸收。但一项最新的研究发现,乙烯信号对Cd 的响应促进了质外体屏障的形成,并减少了Cd 通过质外体旁流的运输[75]。研究结果不同的原因可能是植物种类的不同以及不同的原因乙烯浓度的影响。同时有研究指出,有机酸会影响根内皮层分化。Shakirova 等[76]发现,外源添加水杨酸能激活木质素合成的关键酶苯丙氨酸解氨酶,水杨酸处理的小麦幼苗促进了根内皮层细胞壁的木质化,木质素在根细胞壁的沉积增加,加强了质外体屏障性能。
另有研究发现,根系分泌的有机酸会提高与木质素和木栓质生物合成相关基因的转录水平。为验证这一观点,Colmer 等[77]研究了4 种浓度较低、不会对根系产生毒害作用的有机酸对水稻根部径向氧气损失的屏障建立的影响。结果表明,这些有机酸可以诱导根部建立阻止径向氧气损失的屏障,且这种屏障的主要成分是木栓质。
3.4 生物胁迫对内皮层分化的影响
除非生物胁迫外,在包括病原菌、线虫等在内的生物胁迫也会影响内皮层的分化,导致木栓质沉积增加,以减轻有害生物对植物的危害[78-80]。根际微生物群落与内皮层之间的联系在近2 年才开始被研究。Salas-Gonzalez 等[4]在拟南芥的研究中发现,根际微生物群落与内皮层的协调作用可以维持植物体内矿质营养的动态平衡。控制根内皮层分化的调控网络也影响植物根际微生物群落的组成;植物根际微生物群落中的个别成员控制根内皮层发育,特别是对木栓质沉积的影响,从而影响到植物地上部的离子组成以及在非生物胁迫或生物胁迫下植物的耐性。这一研究开辟了内皮层与根际微生物群落存在互作关系的先河,将会是未来的研究热点。但是目前仍然存在许多谜团是我们未解开的。例如,转录因子MYB36、MYB41 是否影响根际微生物群落的活性,根际微生物群落与内皮层之间互作的机制是怎样的,还需要学者们通过更深层次的研究来确定。
另有最新研究显示,在根际接种促进植物生长发育的细菌如枯草芽孢杆菌、曼德式假单胞菌可以提高植物的抗盐性[81]。接种细菌的植株在盐胁迫下根内皮层中的木质素和木栓层的沉积加速。枯草芽孢杆菌产生细胞分裂素(cytokinin,CTK),曼德式假单胞菌产生生长素(auxin,IAA),在小麦根际接种这2 种细菌后,观察到根内皮层中凯氏带的形成加快,并且在盐胁迫下,植株体内Na 的积累量显著降低。接种此类根际细菌可以促进内皮层的分化,并且提高植物的抗盐性。值得考虑的是,在植物根际接种细菌影响了内皮层的发育进程,究其本质,是细菌本身对内皮层分化的影响,还是其产生的激素对内皮层分化的影响还需更深层次的研究来确定。
4 展望
虽然我们现在对于根内皮层细胞的了解一直在稳步增加,但是不得不承认,我们对其发展和功能的了解仍十分有限。例如,凯氏带被认为是主要以木质素为基础的木质化结构,但到现在为止,凯氏带的详细组分仍存在争议。而木栓层作为继凯氏带之后的另一个内皮层屏障正受到越来越多的关注,但是对于木栓质的生物合成、单体运输及组装等方面的研究仍存在一些空白之处。在拟南芥凯氏带缺陷突变体如sgn1 的研究中发现了木栓质的异位沉积,并且这种异位木栓化是植物自主发生的。异位木栓化可能是植物遭受逆境胁迫时的一种自我保护机制,但是这种异位木栓化的具体调控机制目前尚不清楚。随着遗传学、分析化学,以及分子生物学等学科的不断进步发展,我们对根内皮层分化的分子机制的认识不断提高,学者们已经鉴定出多种与内皮层分化相关的基因,并且研究对象也从之前的模式植物拟南芥逐渐向水稻、玉米等作物转变。在过去的几十年里,人们研究了广泛的非生物胁迫对根内皮层分化的影响,其研究成果将为育种工作者培育耐盐、耐旱等新品种作物提供理论支撑。
前人的研究大多集中于非生物胁迫对根内皮层分化的影响,即以非生物胁迫为手段观察内皮层的发育情况。但是对植物生长发育必需的大量营养元素如N、P、K 对内皮层分化的影响研究较少。而针对我国肥料利用率不高、过量施肥带来的各种环境问题,根内皮层是一个很好的切入点。内皮层分化在营养胁迫下的可塑性这一重大发现为我们研究植物对养分的高效吸收利用提供了全新的思路。因此,在未来对于内皮层的研究重点应该放在大量营养元素与内皮层之间的互作关系上,利用激素诱导或信号转导等途径,充分开发内皮层的高度可塑性,实现植物对水分和养分的高效吸收和利用应是我们重点研究的目标。
