Siglec-15抗体在非小细胞肺癌中的调控机制研究
2023-03-13李延康贺钢枫
李延康贺钢枫
(包头市中心医院(内蒙古医科大学包头临床医学院)心胸外科,内蒙古 包头 014000)
2020年全球统计新发肺癌患者近220万例,死亡人数占比达18%。肺癌被认为是危害人类健康的主要恶性肿瘤之一[1]。手术行肺段、肺叶乃至全肺切除是IA期至IIIA期可耐受手术患者的第一选择。而针对无法耐受手术的患者或临床分期在IIIB期至IV期的患者,如果同时具备表皮因子生长受体(epidermal growth factor receptor, EGFR)、细胞间质上皮转换因子(cellular-mesenchymal epithelial transition factor, c-Met)或Kirsten大鼠肉瘤病毒癌基因同源物(kirsten rat sarcoma viral oncogene homolog, KRAS)等基因位点的阳性突变,可以根据突变结果选择针对性靶向治疗药物[2-3]。但靶向治疗药物的获益群体仅限于非小细胞肺癌(non-small cell lung cancer, NSCLC)驱动基因突变且有对应药 物的患者,对于没有驱动基因获益的NSCLC患者群体和小细胞肺癌(small cell lung cancer, SCLC)患者群体而言,免疫检查点抑制剂(immune checkpoint inhibitors, ICIs)是除去传统放疗、化疗治疗外,又一可选治疗策略(图1)。肿瘤微环境(tumor microenvironment, TME)中的程序性死亡受体-1(programmed cell death 1, PD-1)和程序性死亡受体-配体1(programmed cell death-ligand 1, PD-L1)分子过度表达是部分实体肿瘤患者的主要免疫逃逸机制,抗体阻断PD-1/PD-L1的相互作用可以使受损的免疫功能恢复正常,且不会产生其他过多的副作用。部分PD-1/PD-L1已经获批用于NSCLC和SCLC的一线使用[4-6],是目前ICIs治疗NSCLC的主要策略。但需要引起注意的是,多中心、多样本的统计回顾表明,PD-1/PD-L1在NSCLC中的获益占比仅约为20%[7]。针对PD-1/PD-L1无应答或已产生耐药性的现状,后续抗肿瘤治疗策略如何选择陷入困境之中,目前已有众多相关试验开展,以寻求抗肿瘤治疗新方向。本文将就唾液酸结合免疫球蛋白样凝集素 -15(sialic acid-binding immunoglobulin-like lectin, Siglec-15)以及其相关抗体在小鼠与人源NSCLC中的调控机制和临床应用的研究进展作一综述。
1 Siglec-15的生物学特征
Siglec家族是一类免疫球蛋白样凝集素,其成员多含有免疫受体酪氨酸抑制基序(immunoreceptor tyrosine-based inhibitory motif,ITIM),通过识别含有唾液酸(sialic acid, SA)的糖链结构,介导抑制性受体信号,并在人类癌细胞和肿瘤浸润髓系细胞中广泛表达,在免疫调节动态平衡方面起着重要作用。Siglec家族总体分为两类,一类是序列可变的,如Siglec-7、Siglec-9;另一类是序列不可变的,如Siglec-15[8]。Siglec家族作为I型跨膜蛋白,其胞外由两种免疫球蛋白样结构域组成,分别为数个2型免疫球蛋白恒定区(immunoglobulin cinstant 2, IgC2)和1个免疫球蛋白可变区(immunoglobulin variable, IgV),其间由跨膜结构域连接[9]。Siglec-15作为Siglecs家族的成员是非常独特的,仅包含1个IgC2和1个IgV;而且其胞质内并没有ITIM,在免疫细胞及组织中普遍缺失,仅选择性在髓系细胞和破骨细胞中表达[10]。研究指出,当Siglec-15发挥免疫作用时,其跨膜区赖氨酸274位点可以与DNAX激活蛋白-12(DNAX activation protein-12, DAP-12)结合;而DAP-12自身拥有一个ITIM,可以募集酪氨酸激酶并使其磷酸化,进而向下游传递信号[11]。研究表明,Siglec-15的IgV结构域中含保守的精氨酸(R143)基序,在与SA结合的过程中起到关键性作用[12],其可以作为巨噬细胞受体,识别在肿瘤中广泛表达的SATn抗原。而SA作为一种人体上表达的糖类,可以用来区分自体和非自体,同时介导细胞与病原体之间的相互作用[13]。另有研究表示,一些病原体包括恶性肿瘤细胞,可以通过抑制Siglec家族来抑制自体免疫反应,延长自身的存活周期[14]。
2 Siglec-15参与免疫调节
Siglec家族成员作为抑制性受体在多种淋巴和髓系来源的免疫细胞上表达,通过在抗原呈递细胞上的相关实验证明,Siglec-15的表达抑制了小鼠T细胞的增殖,同时削弱了细胞因子的分泌以及杀伤力[15],而这些动物实验表明,Siglec-15可能通过某种机制尚不完全明确的受体,达到抑制小鼠T细胞,进而可能影响人类的T细胞的工作(图1)。由此进一步实验,建立实验性自身免疫性脑脊髓炎(experimental autoimmune encephalomyelitis, EAE)小鼠模型,验证Siglec-15对T细胞的体内作用。与对照组相比,Siglec-15基因敲除(knock out, KO)小鼠和注射Siglec-15胞外区融合蛋白的小鼠中,EAE程度加重明显,T细胞反映显著增强。在另一方面,与PD-L1 KO小鼠表达类似的是:Siglec-15 KO小鼠在鸡卵白蛋白多肽刺激下,在血液中表现出比野生型(wild type, WT)小鼠更多的I型T特异性识别鸡卵白蛋白(ovalbumin T-I, OT-I)细胞增殖。探究其作用机制,白介素-10(interleukin-10, IL-10)在其中可能起到关键的作用,因为Siglec15 KO小鼠的血清IL-10水平低于WT小鼠,而IL-10单抗消除了WT小鼠和Siglec-15 KO小鼠在OT-I型T细胞增殖方面的差异[16](图1)。分析细胞增殖以及利用流式细胞术检测细胞凋亡的实验可证实Siglec-15通过调节细胞生长抑制T细胞作用[17]。另有研究指出,Siglec-15的抗体可由核因子κB受体活化因子配体和巨噬细胞集落刺激因子共同诱导的多核破骨细胞生成[18]。因肿瘤坏死因子(tumor necrosis factor, TNF)中的TNF-α和TNF-β可由多种上皮细胞、基质细胞、干细胞释放,有理由推测Siglec-15上调巨噬细胞表达可能是自然免疫以及获得性免疫的负反馈调节途径。值得一提的是,一些病原体,例如乙型链球菌,甚至已经衍生出相关的蛋白质,与髓系细胞上的抑制性Siglec家族成员结合,从而逃避免疫介导的杀伤[19]。
3 Siglec-15参与肿瘤相关表达
不可手术的NSCLC的治疗策略一直是困扰临床医生抉择的难题。不同病理类型的NSCLC、不同患者的自身个体差异等诸多因素,使得肿瘤治疗策略的个体化制定尤为重要。尽管PD-1/PD-L1为主要代表的免疫治疗在全世界范围内取得了优良的治疗效果,但并不是所有患者都具备PD-1/PD-L1阳性获益,而且免疫治疗后期的耐药现象也不可避免[20]。在一项入组291例患者的胰腺导管腺癌的队列研究中表明,Siglec-15和PD-L1的单独阳性表达率分别为18.6%和30.3%,Siglec15和PD-L1共同阳性表达率为6.1%,而PD-L1阴性表达同时Siglec-15阳性表达率则为18%[21]。Siglec-15作为近年来新发现的肿瘤免疫逃逸途径,有研究指出,PD-L1的主要诱导剂干扰素-γ(interferon-γ, IFNγ)显著抑制了巨噬细胞中Siglec-15的表达,使得Siglec-15与PD-L1的表达互斥[22](图1)。换而言之,Siglec-15会出现与PD-L1互补的表达谱,使其可能成为解决免疫抑制的新靶点。与各自正常组织相比,Siglec-15mRNA在肺癌、结肠癌以及肾癌等多种肿瘤中表达显著上调,通过定量免疫荧光对200余例NSCLC标本进行免疫组化分析的进一步细化研究表明,共有25.7%的样本表达Siglec-15蛋白[15]。而一项多中心的针对超过600余例NSCLC标本的检测中发现,与KRAS突变相比,EGFR突变的肿瘤中Siglec-15表达更高,且与其它病理学特征无关[23]。具体到NSCLC中不同的病理分型而言,腺癌中的Siglec-15表达量显著高于鳞癌,而鳞癌中的PD-L1表达量却又高于腺癌[24]。不同病理类型肿瘤中TME的免疫逃逸机制高度不同,其中TME由肿瘤细胞、成纤维细胞、血管细胞、肿瘤浸润性淋巴细胞(tumor-infiltrating lymphocytes, TILs)等组成。而TILs含有不同的细胞亚群,并在TME中发挥不同的作用,拥有正向免疫调节的细胞中则包含树突细胞、自然杀伤细胞(natural killer cell, NK)和CD8+T细胞(cluster of differentiation 8 positive cell, CD8+T)等。这类细胞可以识别肿瘤细胞,并达到杀伤和清理的目的。Hao等[24]实验证明了Siglec-15在肺腺癌的表达与TILs中CD8+T细胞的密度呈正相关,但值得注意的是,Siglec-15表达含量多寡并不能用来判断NSCLC预后,与TME中的TILs关系也尚不完全明了(图1)。与人患肿瘤相对的是,常见的小鼠肺癌、结肠癌和黑色素瘤的模型中,均未检测到Siglec-15 mRNA的表达。但在含有丰富的肿瘤相关巨噬细胞(tumor-associated macrophage, TAM)浸润的B16-粒细胞巨噬细胞集落刺激因子(B16-granulocyte macrophagecolony stimulating factor, B16-GMCSF)模型(PD-1不敏感)中,可以检测到Siglec-15mRNA的表达[25]。Siglec-15 KO小鼠在移植了B16-GMCSF后,利用质谱流式细胞仪分析提示TILs中CD8+T细胞、NK细胞和部分髓系细胞显著增加,而TAM减少;对比WT小鼠,KO小鼠的肿瘤细胞生长速度减缓,其生存时间延长。上述相关实验结果表明,巨噬细胞以及Siglec-15在肿瘤免疫中具有重要的抑制作用。虽然在功能上与PD-1/PDL1相似,但Siglec-15介导的免疫抑制机制却是与PD-1/PD-L1途径不同,这两条途径代表了各自独立的免疫调节机制,因此,阻断Siglec-15通路可以为PD-1/PD-L1途径失败的患者提供一个新的免疫治疗方向。
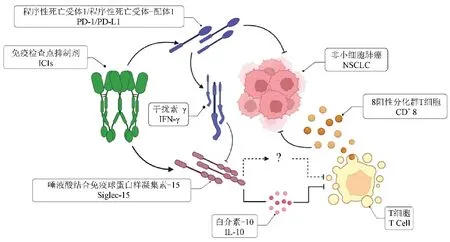
图1 Siglec-15对非小细胞肺癌的调控作用Figure 1 Regulatory effect of Siglec-15 in non-small cell lung cancer
4 Siglec-15作为免疫抑制分子的意义
Siglec-15通路的发现在部分程度上解释了现阶段免疫治疗的困境:为什么PD-1/PD-L1治疗只对约20%NSCLC患者有效。这些研究从侧面证实PD-1/PD-L1是肿瘤免疫逃逸机制之一,而并非唯一通路。对于PD-1/PD-L1治疗失败或无应答的患者而言,抗Siglec-15可以作为一个新的治疗靶点:作为单药应用在临床抗实体肿瘤当中,亦或是寻求与PD-1/PD-L1联合使用,寻求双单抗联用的协同效应。Siglec-15在体内的高表达提示在部分患者的TME中,Siglec-15免疫抑制通路发挥主要作用[16]。因为其与PD-1/PD-L1天然的表达互斥,意味着此时PD-1/PD-L1通路并不是免疫抑制的主要因素,进而可以用Siglec-15的表达作为生物标记物,来预估PD-1/PD-L1的治疗效果。继PD-1/PD-L1信号通路获得成功之后,针对Siglec-15通路的发现及目前临床应用反馈而言,肿瘤免疫逃逸的潜在机制值得更深入的研究。利用免疫组化定量分析为进一步研究TME提供工具基础,积极寻找TME免疫调控的新方向,用以指导NSCLC的精准免疫治疗。
5 Siglec-15抗体治疗NSCLC的探索
基于对Siglec-15的通路特征和表达模式的临床前研究,一款靶向Siglec-15的单克隆抗体NC318被投入到I/II期临床试验(NCT03665285)当中。NC318通过B淋巴骨髓细胞杂交瘤制备,可以与人/鼠的Siglec-15特异性结合。此单抗可以在体外环境下与Siglec-15结合并破坏TAM介导的活动,例如促进肿瘤细胞的生长与转移、抑制T细胞作用以及刺激肿瘤血管生成,进而直接影响疾病的进展[26]。对于人实体肿瘤而言,它可以阻断Siglec-15信号通路,使免疫环境正常化,恢复免疫系统对恶性肿瘤细胞的杀伤能力[26]。在非随机、开放标签、多中心的I/II期临床试验中,主要以NSCLC、乳腺癌、卵巢癌、结直肠癌以及黑色素瘤患者为入组对象,而针对于PD-1耐药的NSCLC患者群体,则单独分组治疗观察。根据公布的前期给药方式及剂量显示[27],NC318采用静脉注射的方式应用给所有入组的晚期实体肿瘤的患者,剂量为8~1600 mg,每周2次(quaque 2 weekly, q2w)。值得注意的是,所有入组患者均未检测体内PD-1或Siglec-15的表达水平,而所有入组的NSCLC患者均曾接受过PD-1/PD-L1的单药或联合治疗。相关数据显示在NSCLC患者群体中应用NC318后,有1例患者完全缓解,1例患者部分缓解、3例患者疾病稳定、7例对PD-1/PD-L1治疗无效的受试者中,有5例观察到单药的抗肿瘤活性;统计分析其客观缓解率(objective response rate, ORR)为29%,疾病控制率为71%。在药物毒副作用方面,NC318在多种剂量水平中表现出了良好的耐受性,没有涉及剂量限制性毒性的相关问题。常见的治疗不良事件包括了腹泻、淀粉酶升高、脂肪酶升高和瘙痒等;而免疫相关不良事件包括了葡萄膜炎、肺炎和白癜风等。在第2阶段临床试验中,调整NC318的应用剂量为400 mg,q2w。2021年8月,对NC318 II期临床试验做出调整,更改治疗方案为800 mg,q1w。目前临床患者招募仍在进行中,更多的相关试验结果有待进一步统计分析。
2022年,美国癌症研究协会年会上报告了有关Siglec-15抗体的最新研究,表明另一款Siglec-15的单克隆抗体BSI-060T在体外及小鼠实验中获得了良好的数据,作为一款全人源的单克隆抗体,BSI-060T与NC-318相比其在亲和力、生物活性和半衰期方面具有良好的数据回报[27]。临床前研究指出,BSI-060T与Siglec-15结合,在体外阻断了Siglec-15与其受体之间的相互作用;在体外T细胞反应测定中,BSI-060T可减轻Siglec-15介导的CD8+T细胞的增殖抑制和IFNγ的释放;在人源化Siglec-15小鼠肿瘤模型中,BSI-060T表现出对肿瘤细胞生长的显著抑制作用且耐受性良好;在非人灵长类动物的研究中,BSI-060T展现出良好的药代动力学[27]。目前BSI-060T正在申请开始I期临床试验,更多的相关试验结果仍需进一步回报。期待Siglec-15抗体在NSCLC治疗中获得突破性进展。
6 Siglec-15抗体治疗NSCLC的展望
确定哪些患者可以从Siglec-15单抗的治疗中获益最大,是目前临床相关研究中争论最多的一环。结合没有采用任何生物标志物检测的NC318 I期临床试验结果分析,在全部入组的实体瘤患者当中,NSCLC的治疗效果最为明显[28],而具体的反应机制尚未明确,初步分析可能与肿瘤内部Siglec-15表达水平有关。在对NSCLC的Siglec-15检测表明,有25%的样本呈现Siglec-15阳性表达[15],而这一数据又与NC318在NSCLC中的ORR相近,结合Siglec-15与PD-1/PD-L1相反的表达模式特性,确定哪些生物标志物可以指导患者的治疗选择,以及预测Siglec-15单抗的疗效,仍需更多的临床数据和统计分析。Siglec-15作为具有与PD-1/PD-L1不同表达及功能的新一代肿瘤相关免疫抑制因子,可能在未来为NSCLC免疫治疗提供新的选择方向。进一步深入研究Siglec-15抗体的作用机制,探讨可能的治疗策略,将有助于设计双特异性抗体以及探索联合治疗方案,提高肿瘤患者的自身免疫力,完善对PD-1/PD-L1治疗不敏感的肿瘤免疫治疗。
