全反式维A酸通过AKT和ERK1/2通路影响IL-22诱导HaCaT细胞中的Cx43和GJIC功能
2023-03-09梁晓冬谈桂其李春红邓新华梁景耀张锡宝
梁晓冬,谈桂其,李春红,邓新华 ,梁景耀,张锡宝
细胞间隙连接通讯(gap junctional intercellular communication,GJIC)是多细胞生物中常见的通讯方式。GJIC通过连接蛋白 (connexins,Cx) 组成的间隙连接 (gap junction,GJ) 通道,以实现连接相邻细胞作用[1]。最近的研究表明,Cx43 参与了生物体内由细胞外囊泡、纳米管或 GJ 介导的细胞交流过程[2]。
全反式维A酸(all-trans retinoic acid,RA)的作用包括调节细胞分化和免疫系统、抗皮脂分泌、抗炎等,对银屑病患者具有重要作用[3]。有学者[4]发现维A酸类药物作为抗肿瘤药物起抑制细胞增殖和诱导细胞凋亡的能力,可以增强GJIC的功能, 但RA 作为GJIC 的一种典型增强剂对银屑病的作用和机制并未引起太多关注。本课题组最近的研究表明,IL-22 通过激活 JNK 信号通路,显著下调 Cx43 表达并降低 HaCaT 细胞的 GJIC 功能[5]。本研究旨在通过探讨 RA 是否影响 HaCaT 细胞中 IL-22 诱导的 Cx43 表达下调和 GJIC 降低过程,以及是否通过丝裂原活化蛋白激酶 (MAPK) 信号通路介导,从而更好地了解 RA 对银屑病的分子机制过程。
1 材料与方法
1.1HaCaT 细胞培养和处理 HaCaT 细胞(Sigma,Shanghai,China)用 Dulbecco′s 改良 Eagle′s 培养基培养,培养环境为 37°C 和 5% CO2的加湿培养箱。第一部分:对于每个实验条件HaCaT 细胞,用 50 μg/L IL-22 (PeproTech Inc., Rocky Hill, NJ, USA) 诱导处理,再用 10 μmol/L RA(Sigma-Aldrich,St. Louis,MO,USA)处理 24 h[5]。第二部分: 分别通过用 5 μmol/L MK-2206(Selleckchem, Houston, TX, USA)和10 μmol/L RA,或 1 μmol/L SCH772984(MedChemExpress, Monmouth Junction, NJ, USA)和10 μmol/L RA处理细胞24 h,来抑制 AKT 或 ERK1/2 信号通路 。
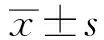
1.3蛋白印迹法(Western blot) 经各组处理后,从 HaCaT 细胞中提取总蛋白后裂解,并测量每个裂解物样品中总蛋白的浓度。将等量的蛋白质加载到 10% SDS-PAGE 凝胶中,并在电泳后转移到聚偏二氟乙烯膜 (Invitrogen, CA, USA),室温封闭 2 h,然后与以下一抗, Cx43、JNK、p-JNK、AKT、p-AKT、ERK1/2、p-ERK1/2(Cell Signaling Technology,MA,USA)、p38(Boster Biological Technology,武汉,中国)、p-p38(Santa Cruz,CA,USA)、 GAPDH(Transgene,Paris,France)孵育过夜。GAPDH 作为对照控制。将膜与 HRP 偶联的二抗(Santa Cruz,CA,USA)孵育。使用 ECL 化学发光系统(Amersham Pharmacia Biotech,NJ,USA)开发免疫反应性印迹。所有结果代表3个独立实验。最后通过 ImageJ 软件 (NIH, MD, USA) 进行定量分析。
2 结果
2.1RA 可抑制IL-22 诱导的 HaCaT 细胞GJIC功能降低过程 在用不同浓度的 RA(0、1、5、10、20 μmol/L)处理 24 h后, SLDT测定显示 RA 以浓度依赖的方式逐渐增强 HaCaT 细胞中的 GJIC功能(图1)。用 10 μmol/L RA 预处理 24 h,有效地抑制了 IL-22 诱导的 HaCaT 细胞中 Cx43 表达降低过程,这表明 GJIC 可能受到相同趋势的影响(图1)。
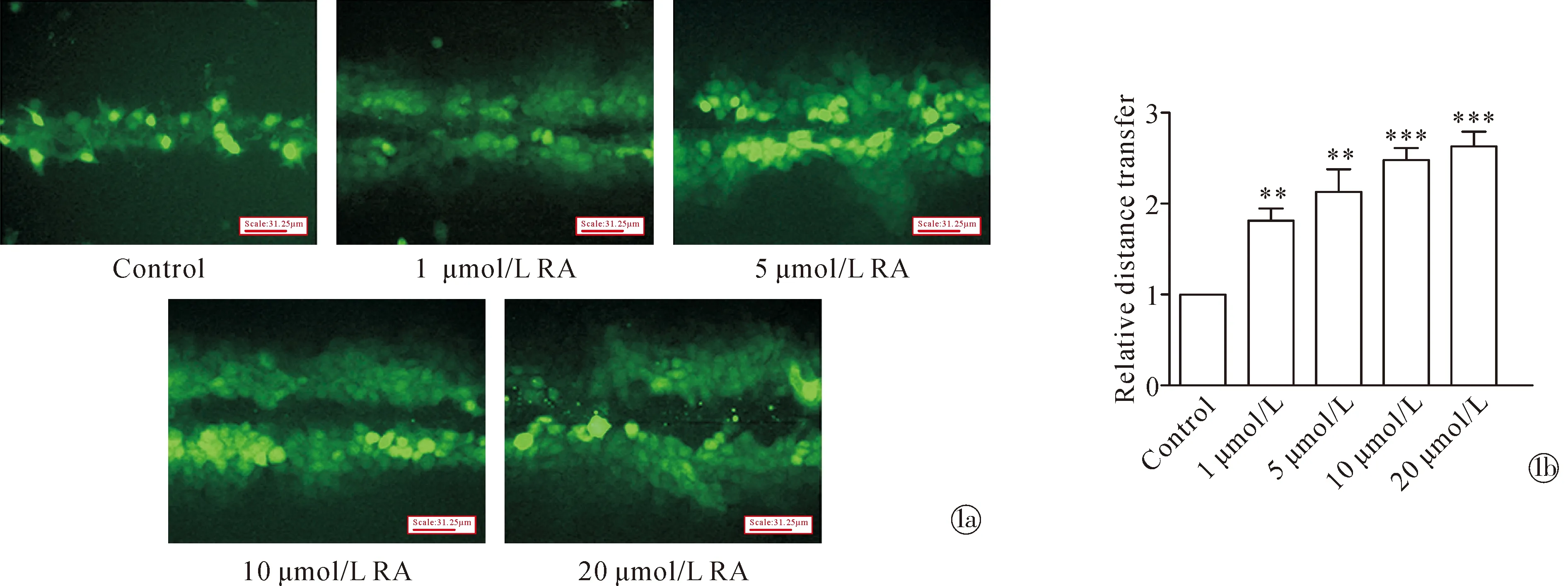
Nore:**P <0.01, ***P <0.001, compared with untreated HaCaT cells, Scale bar= 31.25 mm.Images show GJIC by the scrape-loading dye-transfer method in HaCaT cells treated with various concentrations of RA (0, 1, 5, 10, or 20 μmol/L) for 24 h; The histograms show a quantitative analysis of the distance in Fig.1a图1 RA 对 HaCaT 细胞中 GJIC 的影响Fig.1 The effect of RA on GJIC in HaCaT cells
2.2RA可恢复 IL-22 诱导的 HaCaT 细胞中 Cx43 表达的降低,如图2所示,Western blot显示RA有效地恢复了HaCaT中IL-22诱导的Cx43弱表达水平。然而,RA 并未降低 由IL-22 诱导的 p-JNK 表达增加。这些结果表明,RA 可减少IL-22 诱导的 HaCaT 细胞中 Cx43 表达的降低,但与抑制 IL-22 激活的 JNK 信号通路无关。
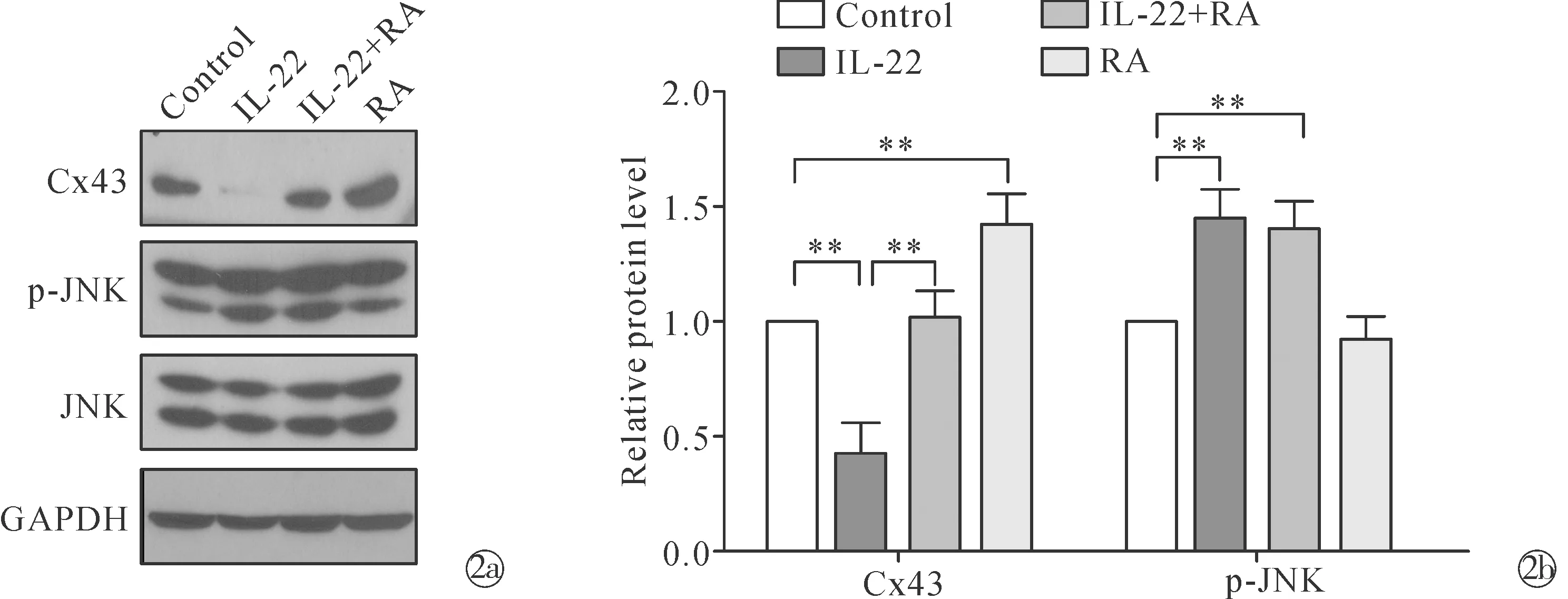
Note:**P <0.01. Western blot shows the protein expression levels of Cx43 and p-JNK in HaCaT cells pretreated with 10 μmol/L RA 24 h before stimulation with 50 μg/L IL-22; The histograms show a quantitative analysis of protein expression levels in Fig.2a图2 RA 对IL-22 诱导的HaCaT 细胞中Cx43 和 p-JNK 表达的影响Fig.2 Effects of RA on expression of Cx43 and p-JNK in IL-22-induced HaCaT cells
如图3所示,在用 IL-22 处理后,Cx43、p-AKT 和 p-ERK1/2的表达显著降低, p-p38的表达无显著改变。RA 预处理部分恢复了 IL-22 诱导的 Cx43、p-AKT 和 p-ERK1/2 表达的降低,表明 RA 可能通过激活 AKT 和 ERK1/2 信号传导途径来恢复 IL-22 诱导的 Cx43 表达下调。
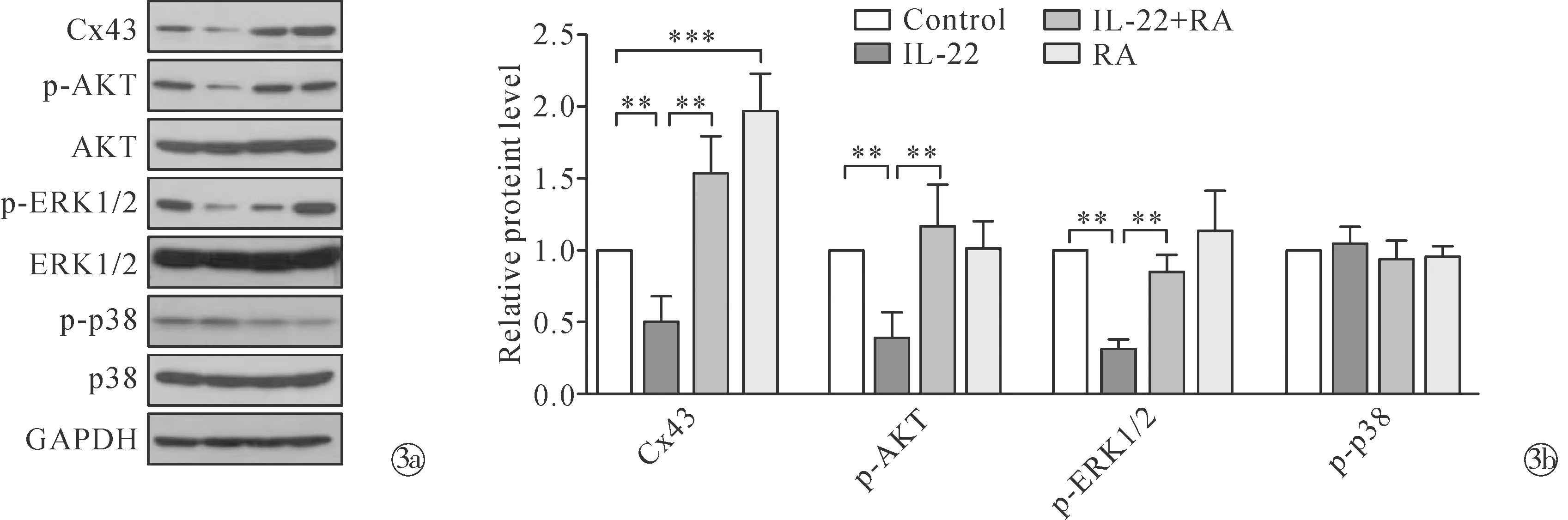
Note:**P<0.01, ***P<0.001.Western blot shows the protein expression levels of Cx43, p-AKT, p-ERK1/2 and p-p38 are shown in HaCaT cells treated with 10 μmol/L RA 24 h and/or before stimulation with 50 μg/L IL-22;The histograms show a quantitative analysis of protein expression levels in Fig.3a图3 RA对IL-22诱导的HaCaT细胞AKT、ERK1/2和p38信号通路的影响Fig.3 Effects of RA on the IL-22-induced AKT, ERK1/2 and p38 signaling pathways in HaCaT cells
2.3RA 通过激活 AKT 和 ERK1/2 信号通路恢复 IL-22 诱导的 Cx43 表达下调和 GJIC 功能降低 用AKT 特异性抑制剂MK-2206预处理细胞后,显著阻断了RA恢复IL-22诱导的HaCaT细胞中p-AKT和Cx43表达上调的影响。类似地,用ERK1/2 特异性抑制剂SCH772984预处理细胞后,显著阻断了RA抑制IL-22诱导的HaCaT细胞中p-ERK1/2和Cx43表达上调的影响。此外,用 MK-2206 和 SCH772984 预处理分别有效地降低了 RA 恢复 IL-22 诱导的 HaCaT 细胞中 GJIC 增加的影响。这些结果表明RA可以通过激活AKT和ERK1 / 2信号通路,以抑制IL-22诱导引起的Cx43表达下调和GJIC功能降低过程,见图4~5。
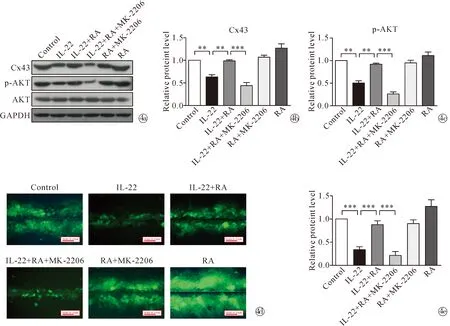
Note:**P<0.01, ***P<0.001, Scale bar=31.25 mm.Western blot shows the protein expression levels of Cx43 and p-AKT in HaCaT cells treated with 10 μmol/L RA, or RA and 5 μmol/L MK-2206 before stimulation with 50 μg/L IL-22;~ The histograms show a quantitative analysis of protein expression levels in Fig.4a; Images show GJIC in HaCaT cells treated with 10 μmol/L RA, or RA and 5 μmol/L MK-2206 before stimulation with 50 μg/L IL-22;The histograms show a quantitative analysis of the distance in Fig.4d图4 RA 对IL-22 诱导的HaCaT 细胞中Cx43、p-AKT 表达和 GJIC 的影响Fig.4 Effects of RA on IL-22-induced Cx43, p-AKT expression and GJIC in HaCaT cells
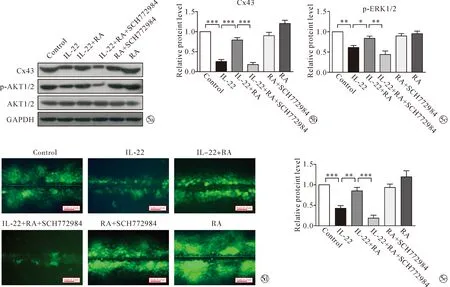
Note:*P<0.05, **P<0.01, ***P<0.001, Scale bar= 31.25 mm. Western blot shows the protein expression levels of Cx43 and p-ERK1/2 in HaCaT cells treated with 10 μmol/L RA, or RA and 1 μmol/L SCH772984 before stimulation with 50 μg/L IL-22;~ The histograms show a quantitative analysis of protein expression levels in 5a; Images show GJIC in HaCaT cells treated with 10 μmol/L RA, or RA and 1 μmol/L SCH772984 before stimulation with 50 μg/L IL-22; The histograms show a quantitative analysis of the distance in Fig.5d图5 RA 对 IL-22 诱导的HaCaT 细胞中Cx43、p-ERK1/2 表达和 GJIC 的影响Fig.5 Effects of RA on IL-22-induced Cx43, p-ERK1/2 expression and GJIC in HaCaT cells
3 讨论
银屑病是一种常见的慢性炎症性皮肤病[6]。角化细胞的分化和免疫系统的激活是银屑病的特征[7]。IL-22可被先天或后天免疫系统激活,IL-22通常在T淋巴细胞介导的疾病中表达升高,如银屑病、类风湿性关节炎和间质性肺病等[8]。IL-22参与多种信号通路,在银屑病的发病、发展和转归中发挥重要作用[9-10]。
IL-22 传统上被认为是 Th17 的细胞因子。然而,淋巴细胞能够分泌 IL-22,例如 Th22、Th1、γδ T和自然杀伤细胞[11]。IL-22是一种重要的细胞因子,介导白细胞和上皮细胞之间的联系,并参与多种免疫介导的炎症性疾病的病理生理过程,如银屑病、肠道炎症和癌症[8]。有研究[12]表明,银屑病患者皮损中IL-22的表达水平明显高于健康个体。在同一银屑病患者中,从病变皮损中分离的 T 细胞产生的 IL-22 水平高于从外周血中分离T 细胞产生的水平[13]。血浆中 IL-22 浓度与斑块型银屑病的疾病活动度呈显著正相关。 IL-22 主要作用于角质形成细胞,以介导角质形成细胞的增殖并抑制其终末分化,导致棘层肥厚、颗粒不足和角化过度特征的银屑病样外观[14]。笔者先前的研究表明[5],IL-22 在 HaCaT 细胞中下调 Cx43表达并降低 GJIC功能,并通过激活 JNK 信号通路在小鼠模型中诱导银屑病样变化。本研究中进一步发现RA 通过激活 AKT 和 ERK1/2 信号通路来恢复 IL-22 诱导的 HaCaT 细胞中 Cx43 表达下调和 GJIC降低,而与 JNK 和 p38 信号通路无关。
RA 已被证明可以上调多种不同细胞类型[15-16]中的 Cx43 表达和 GJIC功能,与笔者在 HaCaT 细胞中的研究一致,尽管在 NTera2/克隆 D1 细胞[17]和 p19 胚胎癌细胞中存在一些矛盾的结果[18]。除了增加 Cx43 表达水平外,RA 通过增强蛋白磷酸酶 2A 和 Cx43 之间的相互作用促进 Cx43 的去磷酸化,从而导致原代人子宫内膜基质细胞中 GJIC 功能的增加[4]。特定位点的磷酸化[16]以及 Cx43 的泛素化[19]已被发现可降低 GJIC功能。尽管RA 对 Cx43 泛素化的直接影响尚未被报道,但在原发性急性早幼粒细胞白血病中,RA 治疗会增加泛素样修饰激活酶 3 的降解,该酶控制着细胞周期进程中许多重要蛋白质的降解[20]。目前RA已被充分证明不仅能抑制细胞增殖和迁移,而且在通过 Cx43 介导的 GJIC 增强细胞对抗癌治疗的敏感性方面发挥重要作用[21-22]。
众所周知,IL-22 通过磷酸化 AKT、p38、JNK、ERK1/2、STAT1、STAT 3 和 STAT 5 中的丝氨酸和酪氨酸残基并与 IL-22 受体 (IL-22R,由 IL-10Rβ1 和 IL-22R1 链组成) 结合来触发多个细胞内途径[23]。RA 强烈诱导树突状细胞亚群中IL -22 结合蛋白 (IL-22BP) 的表达,其可在体外和体内特异性抑制和结合 IL-22[8]。之前与 IL-22 的类似研究表明,RA 的治疗可激活多种信号通路,包括 AKT 和 MAPK[24]。RA通过RARα介导的PI3K/AKT和ERK信号通路,显著降低血管平滑肌细胞的增殖和迁移[25]。在本研究中,IL-22 通过激活HaCaT 细胞中的JNK 和抑制AKT 和 ERK 信号传导来下调 Cx43 的表达并降低 GJIC功能。相反,RA 通过激活 AKT 和 ERK 信号通路而不抑制 JNK 信号通路来逆转这种调节。Tacheau等[26]发现 p38 和 PI3K/AKT 通路的激活协同介导 TGF-β1,可诱导的正常鼠乳腺上皮细胞中 Cx43 表达和 GJIC 的增加。激活的 PI3K/AKT信号通路增加了 β-连环蛋白的核积累和 β-连环蛋白与 Cx43 启动子的结合活性,进而上调了骨细胞中 Cx43 的表达和 GJIC功能[27]。PI3K 催化亚基的组成型活性形式的表达,增加了大鼠肝上皮 T51B 细胞中的 GJIC功能,而成骨细胞系中PI3K 信号传导的药理学抑制,不仅消除了细胞通讯[28]并显著降低了 Cx43 mRNA 和蛋白质的稳态表达[29]。此外,振荡引起的 GJIC 增加取决于骨细胞中AKT 信号的激活,但与 Cx43 mRNA 或蛋白质水平的改变无关[30]。然而Zhou等[31]的一项研究表明,激活 PI3K/AKT 信号可显著降低大鼠海马脑缺血再灌注损伤后的 Cx43蛋白表达和 GJIC功能。AKT信号 激活对于破坏 TNF-α 刺激细胞中的 GJIC 至关重要[32]。同样一些研究表明,ERK 信号通路是否参与 GJIC 的调节是有争议的。先前的结果表明,通过用没食子儿茶素没食子酸酯治疗可以防止人内皮细胞中血清剥夺导致的Cx43间隙连接和GJIC的下调,它激活ERK MAP激酶,但与PI3K/AKT、p38和JNK信号传导的激活无关[33]。在血管平滑肌细胞中,ERK介导血管紧张素Ⅱ诱导的Cx43表达增加和Ser262和GJIC磷酸化,而ERK-siRNA消除了血管紧张素Ⅱ的作用[34]。相反地, ERK信号通路的激活与大鼠肝上皮细胞中过氧化氢引起的Cx43过度磷酸化和GJIC抑制有关[35-36]。总之,以上这些结果表明 PI3k/AKT 和 ERK 信号通路可能对不同细胞和疾病模型中的 Cx43 蛋白表达和 GJIC 功能产生不同的影响。
总之,这项研究有力表明了 RA 通过激活 AKT 和 ERK1/2 信号通路来抑制 IL-22 处理的 HaCaT 细胞中 Cx43 表达和 GJIC 的下调。 这些发现增强了对维A酸在 GJIC 中作用的理解,并可能为银屑病的治疗提供新的靶点。
