苏氨酸对猪生理作用的研究进展
2023-03-06张海泽胡冰艳庞卫军
张海泽 胡冰艳 庞卫军
(西北农林科技大学动物科技学院,杨凌712100)
苏氨酸,也称为β-羟基-α-氨基丁酸,是由Rose于1935年在纤维蛋白质水解物中分离和鉴定出来的。苏氨酸具有4种异构体,但只有L-苏氨酸是在自然界本身存在并具有生物活性,其具有促进蛋白质合成和沉积、调节脂质代谢、促进磷脂合成和脂肪酸氧化以及参与机体免疫功能等作用[1-3]。近年来,合成赖氨酸和蛋氨酸在配合饲料中的应用使苏氨酸逐渐成为影响动物生产性能的限制性因素,苏氨酸已成为以玉米-豆粕型饲粮为主的育肥猪的第二限制性氨基酸[2]。研究发现,在饲粮中添加苏氨酸可以调整饲粮氨基酸平衡、促进猪肠道发育和维持肠道健康等作用[4-5]。近年来,苏氨酸在猪生产领域的研究取得了新进展,本文总结了苏氨酸的代谢途径、作用机制以及苏氨酸在猪生产中的应用,并对潜在的研究方向进行展望,以期能为苏氨酸在猪生产中的实际应用提供思路和方向。
1 苏氨酸的代谢途径和作用机制
苏氨酸是构成机体蛋白质的重要组成部分,特别是肠道黏蛋白[6]。苏氨酸在机体内的代谢途径和其他氨基酸不同,是唯一不经过脱氢酶作用和转氨基作用的氨基酸,而是通过苏氨酸醛缩酶(threonine aldolase,TA)、苏氨酸脱氢酶(threonine dehydrogenase,TDG)以及苏氨酸脱水酶(threonine dehydratase,TDH)催化而转变为其他物质。苏氨酸的代谢途径见图1[7-9],主要有以下3条:1)通过TA代谢为甘氨酸和乙醛,但仅有少部分苏氨酸通过此途径代谢;2)通过TDG代谢为氨基丙酸、甘氨酸和乙酰辅酶A,正常生理条件下,苏氨酸主要通过TDG氧化生成2-氨基-3-氧基丁酸酯,2-氨基-3-氧基丁酸酯在体内进一步降解为甘氨酸、氨基丙酮及乙酰辅酶A;3)通过TDH代谢为丙酸和α-氨基丁酸,动物处于饥饿、空腹或饲喂高水平蛋白质饲粮时,可通过TDH途径使苏氨酸参与肝脏糖异生过程,以维持体内葡萄糖稳态。苏氨酸代谢生成各种重要的产物,包括甘氨酸、乙酰辅酶A和丙酮酸盐等,这些产物在动物的代谢中起着至关重要的作用。

图1 苏氨酸的代谢途径
苏氨酸的作用机制见图2,现有文献表明,苏氨酸有增加蛋白质合成、调节脂质代谢和沉积的能力,以及可以对肠道免疫功能产生积极影响,苏氨酸在体内的作用机制可能通过以下几种方式实现:
1)苏氨酸通过胰岛素样生长因子-Ⅰ(insulin like growth factor-Ⅰ,IGF-Ⅰ)激活磷脂酰肌醇-3-羟激酶(phosphatidylinositol-3-hydroxykinase,PI3K)/蛋白激酶B(protein kinase B,Akt)/雷帕霉素靶蛋白(target of rapamycin,TOR)信号级联反应来促进肌肉蛋白质合成。Zhao等[10]研究证明,饲粮补充苏氨酸导致PI3K和AktmRNA表达的上调,并提高磷酸化Akt与总Akt比率,表明苏氨酸通过PI3K/Akt信号通路增加了肌肉蛋白质的合成。TOR是PI3K/Akt途径的下游成分,可刺激蛋白质合成并促进细胞生长。此外,苏氨酸被TDH降解生成甘氨酸后,可通过PI3K/Akt信号通路以依赖性方式激活哺乳动物雷帕霉素靶蛋白复合体1(mammalian target of rapamycin complex 1,mTORC1),并通过抑制蛋白质水解来调节蛋白质合成,Wang等[11]在猪小肠上皮细胞上的研究证明了这一观点。
2)高脂肪饮食中补充苏氨酸可能通过调节过氧化物酶体增殖物活化受体γ(peroxisome proliferator activated receptor γ,PPARγ)信号通路来调节脂质沉积。Ma等[12]研究表明,高脂饮食诱导的小鼠通过饲喂苏氨酸可以抑制肥胖和降低血清甘油三酯含量,其可能通过PPARγ信号级联与脂肪生成/脂肪分解开关控制合并来调节脂肪沉积。在Jiang等[13]的研究中也发现,苏氨酸缺乏会通过增加脂肪酸和甘油三酯的合成以及减少脂肪酸氧化和甘油三酯的运输来增加肝脏甘油三酯和脂肪酸的积累。
3)苏氨酸通过激活核因子-κB(nuclear factor-kappa B,NF-κB)信号通路调节机体免疫能力。Wang等[14]研究发现,苏氨酸以剂量和时间依赖性方式显着增加猪肠上皮细胞的β-防御素分泌,其通过抑制沉默信息调节因子1(silent information regulator 1,SIRT1)的表达来增强β-防御素的表达,并激活NF-κB信号通路。Zhang等[15]研究也发现,苏氨酸通过激活NF-κB信号通路来调节聚合免疫球蛋白受体(polymeric immunoglobulin receptor,pIgR)水平,从而影响应激条件下肠道黏蛋白以及免疫球蛋白A(immunoglobulin,IgA)的产生。
此外,有研究表明,苏氨酸在胚胎干细胞的分化中作为信号因子发挥重要作用,苏氨酸是小鼠胚胎干细胞(embryonic stem cell,ESC)多能性关键的必需氨基酸,培养基中添加L-苏氨酸可激活ESC的多条信号通路,包括PI3K/Akt、丝裂原活化蛋白激酶(mitogen-activated protein kinases,MAPKs)、哺乳动物雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)、核糖体蛋白S6激酶beta-1(ribosomal protein S6 kinase beta-1,p70S6k)和真核翻译起始因子4E结合蛋白1(eukaryotic translation initiation factor 4E-binding protein 1,4E-BP1),以刺激通过DNA合成前期到DNA合成期(G1/S阶段)的ESC过渡[16]。
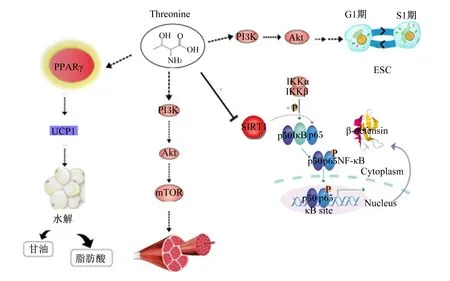
PPARγ:过氧化物酶体增殖物活化受体γ peroxisome proliferator activated receptor γ;UCP1:解偶联蛋白1 uncoupling protein 1;Threonine:苏氨酸;PI3K:磷脂酰肌醇-3-羟激酶 phosphatidylinositol-3-hydroxykinase;Akt:蛋白激酶B protein kinase B;mTOR:哺乳动物雷帕霉素靶蛋白 mammalian target of rapamycin;ESC:胚胎干细胞 embryonic stem cell;SIRT1:沉默信息调节因子1 silent information regulator 1;IKK:核因子-κB抑制物激酶 nuclear factor-κB inhibitor kinase;IκB:核因子-κB抑制物 nuclear factor-κB inhibitor;NF-κB:核因子-κB nuclear factor-κB;β-defensin:β-防御素;Cytoplasm:细胞质;Nucleus:细胞核;κB site:κB位点。
2 苏氨酸在猪生理上的作用
目前苏氨酸在猪生产中的应用研究多集中在断奶仔猪方向,因苏氨酸对肠道的作用,使其在仔猪肠道发育方面的研究备受关注,近年来关于苏氨酸应用于猪生产的部分研究[14,17-24]情况见表1。
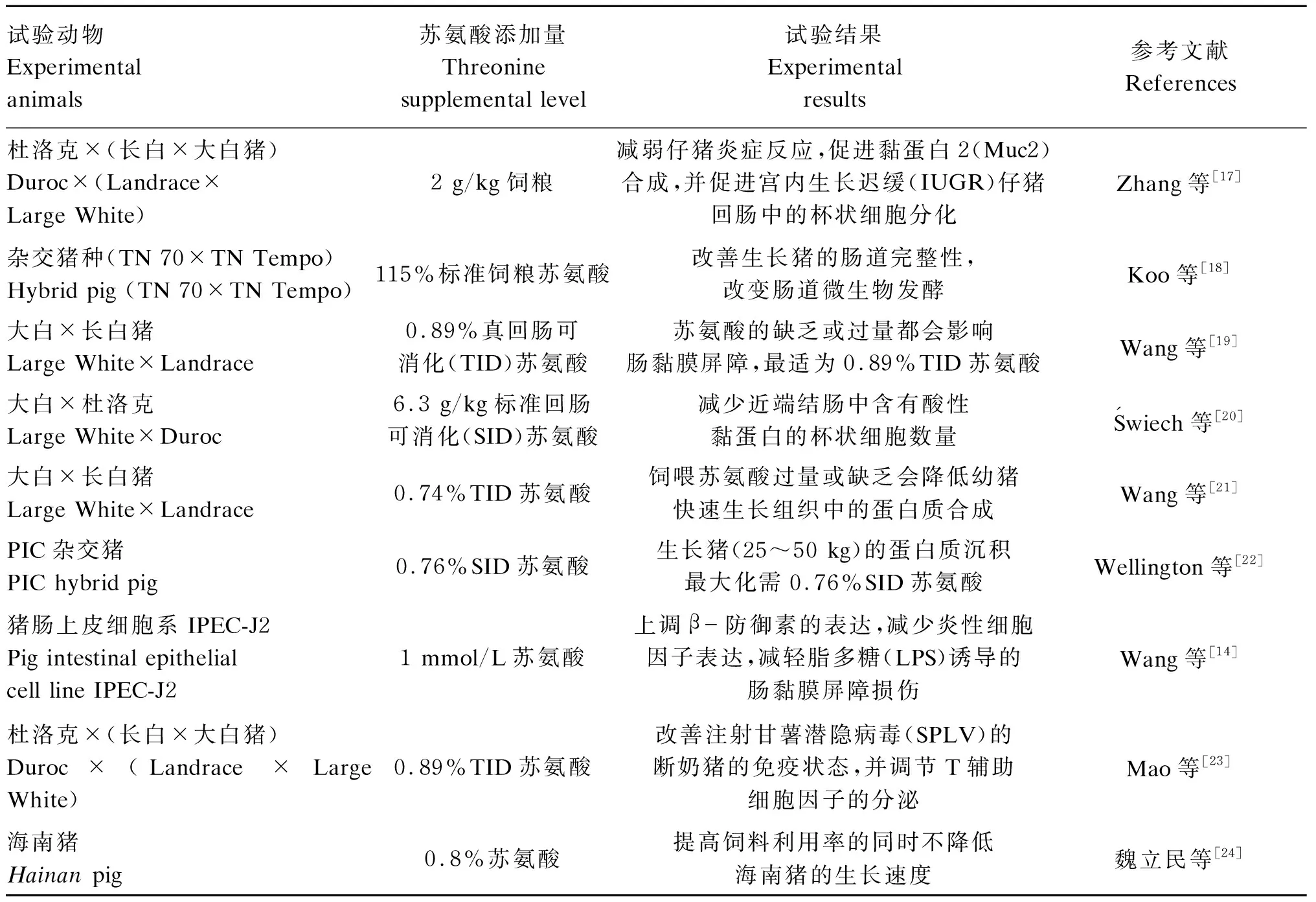
表1 苏氨酸在猪生产中的应用研究
2.1 苏氨酸对肠道结构的影响
肠道是生物体中最重要的器官之一,在保护其他器官免受有害物质侵害方面发挥着重要作用,并在消化和吸收膳食营养物质方面起关键作用[25]。营养物质从外部通过肠道被吸收以支持动物的生命活动,各种有害的病原体和毒素也可能通过肠道攻击宿主[26]。肠道屏障在生命活动中起着重要作用,能保护宿主免受胃肠道中存在的肠道微生物、食物抗原和毒素的侵害[27]。肠道结构的完整性是发挥其营养消化和吸收的功能的基础。研究表明,饲粮苏氨酸限制会显著损害小肠所有部分的黏蛋白合成,其中十二指肠的最大减少量为40%[28]。

总之,苏氨酸对于仔猪肠道的发育、肠道黏膜屏障的形成以及减少肠道炎症发生具有积极的影响,不过,关于仔猪发育所需苏氨酸最适需要量,还需要更多、更细致的研究。苏氨酸过量或缺乏都不利于仔猪发育,后续的研究中可以探究仔猪在不同生长阶段对苏氨酸的需求量的变化情况,这对降低成本、提高生产效率具有重要意义。
2.2 苏氨酸对肠道微生物的影响
出生到30日龄是仔猪从母乳到固体饲粮过渡的转变时期,这个时期的仔猪肠道发育不健全,肠道菌群定殖未成熟,易受到病原微生物的入侵,是仔猪极为脆弱的时刻[29]。哺乳仔猪在此期间更容易发生体重减轻、食欲不振和消化功能障碍等情况[30],而仔猪肠道感染是断奶前后发病的主要原因。越来越多的证据表明,饲粮苏氨酸对仔猪肠道微生物群有积极影响,能够促进有益菌群的增殖。
在保育猪中,Koo等[18]研究表明,在NRC(2012)的标准基础上增加15%的苏氨酸,可通过与饲粮成分相互作用的方式改变肠道微生物发酵;与复杂饲粮(含有多种动物蛋白质来源)相比,简单饲粮(含有豆粕作为蛋白质来源)中的苏氨酸补充剂降低了肠道氨的含量,但增加了肠道挥发性脂肪酸的含量。其他动物中的研究也表明,在饲粮中添加苏氨酸能有效地改变家禽和大鼠的肠道微生物群组成。饲喂大鼠含有L-苏氨酸的氨基酸混合物,增加了被右旋糖酐硫酸钠影响的大鼠中肠杆菌、乳酸杆菌、拟杆菌和肠球菌等有益菌群的频率[31]。Dong等[32]研究发现,饲粮粗蛋白质的减少会降低蛋鸡肠道细菌多样性,而低粗蛋白饲粮中添加苏氨酸可恢复细菌多样性并显著提高潜在有益细菌的丰度,对这种效应的一种解释可能是黏蛋白基因表达的上调,因为黏蛋白不能在小肠中被消化,因此会到达盲肠,充当细菌降解的底物。
最新的研究发现,人母乳中的游离苏氨酸可影响婴儿肠道微生物的组成,Riederer等[33]研究表明,苏氨酸对母乳喂养婴儿肠杆菌科成员具有潜在保护作用;该研究提出了母乳游离氨基酸与婴儿肠道微生物群组成在早期哺乳期间的潜在相互作用,而此前已有研究表明母乳喂养可降低初生重对早产胎儿肠道微生物的影响[34]。因此,母猪饲粮中添加苏氨酸是否会对IUGR仔猪产生有益的影响是值得探究的内容,能否提高仔猪存活率以及断奶重对于猪生产具有重要意义。
2.3 苏氨酸对免疫功能的影响
生产中通常向未断奶的仔猪提供教槽料,以改善仔猪断奶后的生长性能,并提高仔猪断奶前对固体饲粮的适应能力,但由于教槽料和母乳之间存在显著差异,从母乳逐渐过渡到固体饲粮的过程对仔猪来说是产生应激的重要原因[35]。不仅如此,仔猪还需要面临来自固体饲粮中病原体的风险,此时期的仔猪肠道疾病多发,尤其是腹泻问题。此外,饲粮的转变通常会导致消化功能紊乱,进而导致生长性能低下[36]。研究表明,苏氨酸对肠道免疫具有重要作用,苏氨酸可促进淋巴细胞的增殖以及抗体的分泌,同时调节炎症因子的表达。
Wellington等[22]研究表明,生长猪在免疫系统刺激(immune system stimulation,ISS)[注射脂多糖(LPS)]期间对苏氨酸的利用率提高,且生长猪(25~50 kg)蛋白质沉积最大时需要0.76%SID苏氨酸。在另项研究中,生长猪在ISS(注射LPS)下导致对苏氨酸需求的增加,可能在很大程度上归因于被吸收的苏氨酸对合成免疫系统代谢物的利用能力的增强,特别是黏蛋白[37]。Wang等[14]研究表明,L-苏氨酸可以上调β-防御素的表达,并降低猪肠上皮细胞系IPEC-J2细胞中炎性细胞因子的表达;同时,L-苏氨酸能够缓解LPS诱导的猪肠上皮细胞中的肠黏膜屏障损伤。由此可见,L-苏氨酸介导的内源性防御素表达可能是减少抗生素使用,增强动物抗病性和肠道健康的有效方法。在猪伪狂犬病活疫苗激发下,饲粮TID苏氨酸水平的提高减弱了血清干扰素-γ(IFN-γ)含量的提高,以及不同组织中Toll样受体(TLR)3、TLR7和TLR9的mRNA相对表达量的提高,这表明饲粮添加适宜水平的苏氨酸可以通过下调TLR3、TLR7和TLR9在组织中的表达来改善注射SPLV断奶仔猪的免疫状态,从而调节T辅助细胞因子的分泌[23]。
此外,在其他动物中的研究也表明了苏氨酸对免疫功能的积极影响,包括肉鸡[38-39]、鱼类[40]和北京鸭[41]等。在当今全面禁抗的大背景下,提升畜禽本身的免疫能力对于维持生产十分重要。苏氨酸作为家畜必需氨基酸,在机体免疫过程中发挥重要功能,因此,能否使用苏氨酸替代部分抗生素功能的研究将在今后猪生产中发挥重要作用。
3 苏氨酸在猪生产中的特殊意义
3.1 苏氨酸对母猪繁殖性能的影响
在妊娠期间,母猪摄入的氨基酸除了用于自身代谢需要外,还要用于乳腺组织的生长、胎儿的生长发育,母猪妊娠90 d后,由于胎儿重量增加,蛋白质合成急剧上升,对于氨基酸的需求也大大增加,因此母猪妊娠后期对于氨基酸的需求量相比于妊娠前期更高。Levesque等[42]研究表明,在整个妊娠期饲喂单一的氨基酸会导致母猪妊娠早期过量摄入氨基酸,而在妊娠晚期氨基酸摄入不足,且在妊娠最后1/3的时间苏氨酸需求量提高了2倍,这说明妊娠期分期饲喂母猪将更能满足其对营养的需求。
在母猪哺乳期,Cooper等[43]研究表明,哺乳期母猪苏氨酸的需要量为37~40 g/d,而维持仔猪生长需要的苏氨酸为36~39 g/d。此外,Schneider等[44]研究表明,母猪哺乳期SID苏氨酸需求量为0.50%(28 g/d),母猪哺乳期苏氨酸∶赖氨酸为57%。母猪繁殖或仔猪生长速率的改善与每日SID苏氨酸摄入量接近27 g相对应;当SID苏氨酸∶SID赖氨酸为68%或SID苏氨酸为27.6 g/d时,对母猪的繁殖性能具有优化作用;上述结果表明,母猪实际的每日苏氨酸需求量低于NRC(2012)对每日SID苏氨酸的建议量,但需要更高的SID苏氨酸∶SID赖氨酸[45]。Cuaron等[46]研究表明,采食高粱为基础的饲粮补饲苏氨酸,可防止初产母猪血浆免疫球蛋白G含量的减少,试验组母猪自身合成的抗牛血清蛋白的抗体含量也高于对照组。Leonard等[47]研究表明,随着苏氨酸摄入量的增加,妊娠母猪血浆苏氨酸含量呈二次增加,这种增加伴随着血浆赖氨酸含量的二次减少;随着苏氨酸水平的提高,母猪体重也呈二次增加,但是母猪产子数、泌乳量、乳蛋白含量、仔猪初生重和平均日增重均不受苏氨酸水平的影响。
综上可知,妊娠母猪对于苏氨酸的需求随着妊娠期的发展而发生改变,应分别在妊娠早期和晚期重新评估妊娠母猪对苏氨酸的需求。
3.2 苏氨酸对公猪繁殖性能的影响
随着人工授精技术的广泛应用以及非洲猪瘟的影响,增加种公猪的精液产量及精液品质对于节省生产成本具有重要意义。生猪产业对于公猪的营养需要越来越重视,然而苏氨酸在公猪营养方面的应用未见研究。此前李红[48]研究表明,在犬饲粮中添加赖氨酸和苏氨酸能明显提高公犬的精液品质,显著提高公犬的射精量、精液密度和精子活力,并降低精子畸形率。林燕[49]通过对雄鼠的研究发现,提高饲粮中的苏氨酸水平有利于缓解病毒对雄鼠繁殖性能造成的损害,并且苏氨酸通过提高被攻毒雄鼠的免疫力、降低睾丸组织的炎症因子等提高雄鼠的繁殖性能。苏氨酸对于公猪精液质量和繁殖能力的影响值得探究,且因为公猪在实际生产中的特点决定了公猪对于氨基酸的需求较大,因此,探究适宜的饲粮氨基酸水平对于生猪产业具有重要的研究和应用意义。
3.3 苏氨酸应用于低蛋白质饲粮
多年来的研究发现,适当降低饲粮蛋白质水平可以提高饲料利用率,降低蛋白质饲料原料用量和生产成本。许多研究报道,低蛋白质饲粮有效地降低了猪断奶后腹泻的发生率,维持了肠道健康,并对肠道形态和微生物群有明显的影响[50-51]。魏立民等[24]研究发现,海南猪饲粮粗蛋白质水平降低至14%,参考对照组平衡赖氨酸、色氨酸、缬氨酸和精氨酸水平,并补充添加苏氨酸至0.8%,可以在提高饲料利用率的同时保持海南猪的生长速度。江勇等[52]在北京鸭的研究中发现,低蛋白质饲粮中添加苏氨酸可以提高北京鸭体重、平均日增重、平均日采食量和胸肌率,降低料重比。但也有研究表明,降低饲粮蛋白质水平会导致猪生长性能的降低[53-56]。因此,低蛋白质饲粮中苏氨酸的补充剂量以及低蛋白质饲粮中苏氨酸与其他氨基酸之间的平衡还需要更为全面细致的研究。
3.4 苏氨酸与其他氨基酸的联合使用
当体内蛋氨酸和赖氨酸过量时,苏氨酸的需求量也增加,补充适量的苏氨酸可消除因蛋氨酸和赖氨酸过剩引起的体重下降,减轻因色氨酸过剩引起的生长抑制;苏氨酸在TDH的作用下代谢生成甘氨酸,还参与合成丝氨酸、乙酰辅酶A和丙酮酸盐等[7-8],可以起到平衡体内氨基酸水平的作用。此外,苏氨酸与赖氨酸之间的“氨基酸信号”可调节机体脂质代谢,Goda等[57]研究发现,与对照大鼠相比,饲喂低赖氨酸饲粮的大鼠血清赖氨酸含量降低,而血清苏氨酸含量显着升高;当低赖氨酸饲粮中苏氨酸含量受到限制时,饲喂低赖氨酸饲粮的大鼠血清苏氨酸含量提高会导致脂质掺入骨骼肌,从而导致脂肪肌肉组织的形成。
苏氨酸虽具有平衡饲粮氨基酸的作用,但有研究表明,苏氨酸限制有利于延缓肥胖相关代谢功能障碍的发展[58]。同时,也有研究表明,限制食物中必需和非必需氨基酸可以通过多种途径改善机体葡萄糖稳态和维持代谢健康[59-60]。因此,探究饲粮中适宜的氨基酸水平以及苏氨酸的最适添加量,对于发挥出猪的最佳生产性能具有重要意义。
4 小 结
苏氨酸是动物生长所必需的氨基酸,合成赖氨酸和蛋氨酸在配合饲料中的应用使苏氨酸逐渐成为影响动物生产性能的限制性因素。目前关于苏氨酸的研究仍存在一些不足:1)已有许多研究表明其对仔猪肠道健康的重要性,然而,还需要进一步的试验来确定苏氨酸如何调节肠道微生物群、免疫反应和屏障之间的动态平衡;2)在未来养猪生产的应用中,苏氨酸的使用剂量以及与其他氨基酸联合使用的比例值得探究,在低蛋白质饲粮日益成为主流饲养方式的情况下,苏氨酸的合理使用既可以减少总氮排放,又可以平衡饲粮氨基酸,维持猪的需要;3)苏氨酸涉及的营养代谢方面的调节,使得其在公猪营养方面具有一定的研究意义,未来需要填补苏氨酸在这一领域的研究空白。总之,随着苏氨酸研究的深入开展,其在猪生产领域将具有更为重大的应用前景。
