6个大豆GmBES1基因干旱诱导表达及启动子分析
2023-03-02张军陈炯辛尹珺伊王欢刘秋瑾田秋丰白长胜翟莹
张军,陈炯辛,尹珺伊,王欢,刘秋瑾,田秋丰,白长胜,翟莹
6个大豆基因干旱诱导表达及启动子分析
张军1,陈炯辛2,尹珺伊1,王欢1,刘秋瑾1,田秋丰1,白长胜1,翟莹2
(1.黑龙江省农业科学院畜牧兽医分院,黑龙江 齐齐哈尔 161005;2.齐齐哈尔大学 生命科学与农林学院,黑龙江 齐齐哈尔 161006)
BES1/BZR1转录因子在油菜素内酯信号通路中发挥重要作用,该通路调控植物的生长发育及对各种胁迫的抗性。本研究对6个大豆基因在干旱胁迫下的表达量进行检测,同时对6个基因启动子中的顺式作用元件进行预测分析。实时荧光定量PCR结果显示,6个基因在干旱胁迫下的表达量随着时间存在不同程度的变化,其中的表达量升高最明显。启动子序列预测分析结果显示,/////启动子序列中分别含有3、6、4、3、4和7种激素及逆境响应相关顺式作用元件。这些结果为大豆基因的抗旱机制研究及应用提供理论依据。
大豆;油菜素内酯;干旱胁迫;启动子;顺式作用元件
油菜素内酯(Brassinosteroid, BR)是一类植物特有的类固醇激素,在植物生长发育过程中起着广泛的作用,包括细胞伸长和分裂、形态建成、维管分化、开花、育性、种子萌发、衰老和抵抗各种生物和非生物胁迫等[1-2]。BES1(BRI1-EMS-Suppressor)及其同源物BZR1(Brassinazole-Resistant 1)是介导BR信号通路的关键转录因子,该信号通路的开放与关闭取决于BES1/BZR1的活性。当BR信号存在时,BES1/BZR1被蛋白磷酸酯酶快速去磷酸化并激活,与靶基因的E-box或BRRE元件结合,调控其表达,进而启动一系列BR应答事件[3-4]。在缺乏BR的情况下,BES1/BZR1则被激酶磷酸化并失活[5-6]。BES1/BZR1调控的靶基因数量众多。例如,在拟南芥中共鉴定出受BR调控的BZR1靶基因953个,BES1靶基因1609个[7]。能够直接与、、、和等基因启动子区结合,通过调控这些基因的表达,影响拟南芥花粉管发育和植株育性[8]。香蕉通过抑制、和等乙烯合成相关基因的表达,减少乙烯的合成,进而抑制果实的成熟[9]。
研究已表明,外源施用BR能够提高作物的抗逆性,进而增加作物的产量。作为BR合成途径中的重要调控因子,BES1/BZR1转录因子家族基因抗逆性的研究也逐渐开展。油菜的15个BZR基因均可以被外源脱落酸(Abscisic acid, ABA)诱导表达,且它们在低温胁迫下均表现差异表达。其中6个BZR基因响应盐胁迫,11个BZR基因响应干旱胁迫[10]。棉花基因可以快速应答干旱胁迫,外源喷施BR有望提高棉花的抗旱性[11]。小麦通过激活基因的表达提高转基因植株抗旱性[12]。桦树中的5个BZR基因可以响应盐和干旱胁迫,在桦树中过表达可以提高对盐胁迫的耐受性[13]。胡杨在烟草中异源过表达能够提高转基因烟草对盐胁迫的耐受性[14]。前人在大豆中鉴定了16个基因[15]。本研究将对其中6个基因对干旱胁迫的应答及它们启动子中逆境相关作用元件进行分析,探索大豆基因与抗旱性的关系,以期为大豆基因的抗旱机制研究及应用提供理论依据。
1 实验部分
1.1 实验材料
实验所用的‘北豆9号’大豆种子来自齐齐哈尔大学植物分子育种研究室。
1.2 水培大豆幼苗及干旱处理
将草炭土与细沙按1∶1比例混合,播种经挑选后的饱满大豆种子。一周后,将生根的大豆移入Hoagland营养液中水培至第一片三出复叶叶片展开。在Hoagland营养液中溶解20%的PEG8000,将幼苗移入进行模拟干旱胁迫处理。在处理的0, 1, 2, 5, 10和24h分别取样,均取第一片三出复叶,迅速置于液氮中并转移至-80℃超低温冰箱中储存。
1.3 RNA提取及反转录cDNA
使用Takara公司的RNAiso Plus试剂提取各时间点样本总RNA,第一链cDNA的合成使用Innovagene公司的cDNA反转录试剂盒。
1.4 实时荧光定量PCR(qPCR)
登陆NCBI数据库(http://www.ncbi.nlm.nih.gov)搜索并下载大豆基因的mRNA序列。利用Primer 5软件,根据6个基因的开放阅读框序列,分别设计qPCR引物。选择大豆基因作为qPCR的内参基因[16]。基因的引物序列均列于表1中。使用Innovagene公司的SYBR Green qPCR Premix试剂盒,在BIO-RAD CFX96 Real-Time PCR仪上进行qPCR,体系和参数的设置均参照邱爽等[16]。

表1 实时荧光定量PCR引物序列
1.5 启动子预测分析
从Phytozome数据库(http://phytozome-next.jgi.doe.gov)下载6个基因的启动子序列(基因起始密码子ATG上游2000bp序列)。分别将6个基因的启动子序列输入PlantCARE在线软件(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/),预测启动子中顺式作用元件的数量和功能。
1.6 数据处理
利用SPSS软件中Duncan's multiple range tests法分析数据差异显著性。
2 结果与讨论
2.1 6个GmBES1基因在干旱胁迫下的表达分析
图1为qPCR结果,显示6个基因在干旱胁迫下的表达量随着时间存在不同程度的变化。表达量在处理后高于对照(0h),最大值出现在处理后24h,约为对照5倍左右;表达量在处理后先下降后升高,最大值出现在处理后24h,约为对照3倍左右;表达量在处理后出现两次峰值,均为对照4倍左右;表达量在处理后的1, 2, 10 h显著高于对照,最大值出现在处理后10h,约为对照14倍左右;表达量在处理后升高,最大值出现在处理后24h,约为对照2.5倍左右;表达量在处理后的2 h和24h显著高于对照,约为对照1.5和2倍左右。综上,在干旱诱导程度上,的表达量升高最明显。
由以上结果推测,6个基因应该与干旱胁迫相关,它们在大豆幼苗应对干旱胁迫时可能发挥作用。相关研究已发现,拟南芥通过直接负调控干旱应答基因的表达,从而介导BR和干旱途径之间的信号交叉[17]。对干旱胁迫下的棉花喷洒BR能够引起的快速表达,外源喷施BR有助于提高棉花的抗旱能力[11]。但6个基因在干旱胁迫下的表达方式存在不同,也间接预示基因家族在大豆抵御干旱胁迫中的功能存在多样性[18]。基因的表达模式与功能密切相关,干旱应答的BES1/BZR1转录因子基因过表达后提高转基因植株抗旱性已有报道[12],这也为后续大豆基因在抗旱育种中的应用提供了依据。
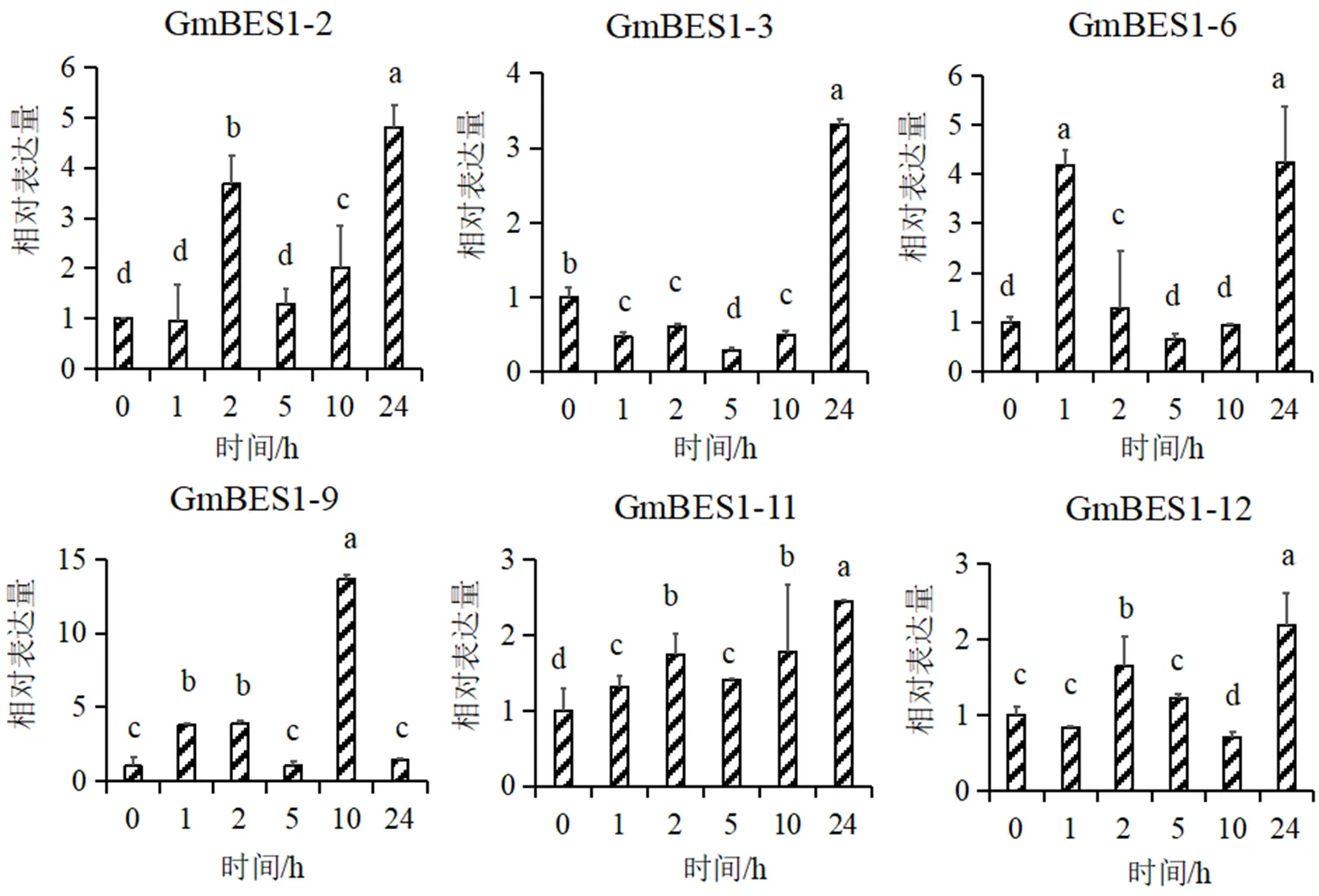
图1 6个GmBES1基因在干旱胁迫下的表达不同字母表示差异显著(p小于0.05)
2.2 6个GmBES1基因启动子顺式作用元件预测分析
通过在线软件对基因上游启动子区域的顺式作用元件进行预测分析。结果如表2所示,6个基因启动子中除了含有启动子的特定元件TATA-box和CAAT-box外,还含有不同种类和数量的激素及逆境相关顺式作用元件。5个启动子(除外)均含有2~3个ABA响应元件(ABRE);和的启动子中还含有茉莉酸甲酯、赤霉素及水杨酸响应元件;此外,和启动子中也含有水杨酸响应元件。
ABA、茉莉酸、赤霉素和水杨酸等植物激素及其信号转导通路在环境胁迫过程中起重要作用[19],这类顺式作用元件的存在预示着基因可能参与了激素的信号途径,尤其和的启动子可能受多种激素信号的调节。、、和的启动子中均含有MYB转录因子结合位点。MYB转录因子家族基因通常与植物逆境相关[20],推测这4个基因可能是MYB转录因子调控的靶基因。BES1/BZR1基因能够应答低温胁迫在之前已被报道[10,21-22]。、和的启动子中也含有低温响应元件(LTR),这些基因的表达能否被低温胁迫所诱导还需要进一步验证。此外,5个启动子(除外)还含有1~2个厌氧诱导元件(ARE),启动子还含有1个防御和胁迫响应元件(TC-rich repeats)。以上这些预测结果与6个基因在大豆幼苗能够应答干旱胁迫的结果是相符的。qPCR的结果显示,在6个基因中,对干旱胁迫的响应最明显,但在启动子中预测到的逆境相关顺式作用元件最少,推测其启动子中可能尚存在未被鉴定的逆境相关元件。
3 结论
实时荧光定量PCR结果显示,6个基因在干旱胁迫下的表达量随着时间存在不同程度的变化,其中的表达量升高最明显。启动子序列预测分析结果显示,/////启动子序列中分别含有3、6、4、3、4和7种激素及逆境响应相关顺式作用元件。上述结果为大豆基因的抗旱机制研究及应用提供理论依据。
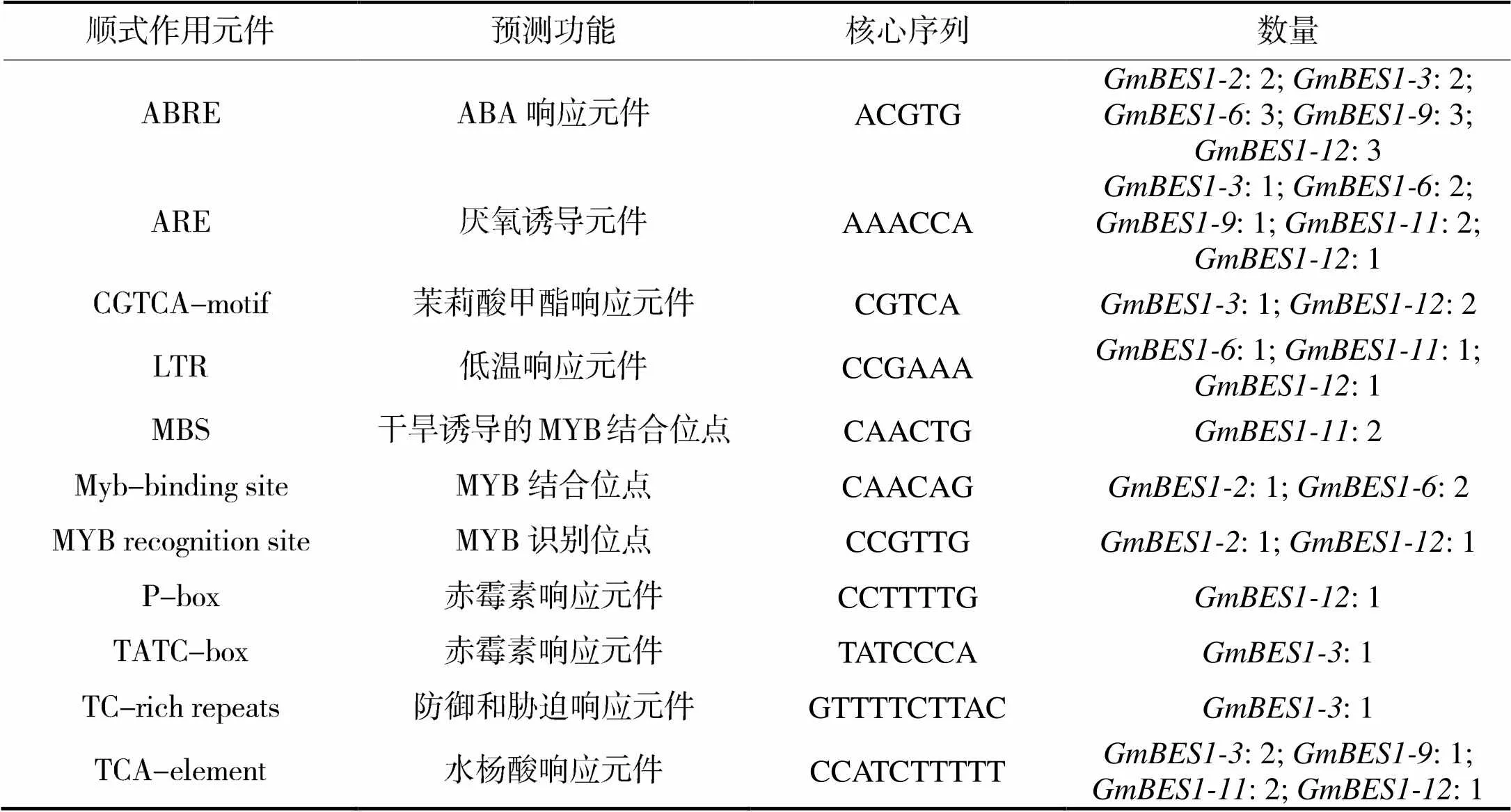
表2 6个GmBES1基因启动子中的顺式作用元件
[1]LIU X, YANG Q, WANG Y, et al. Brassinosteroids regulate pavement cell growth by mediating BIN2-induced microtubule stabilization[J]. Journal of Experimental Botany, 2018, 69(5): 1037-1049.
[2] NAWAZ F, NAEEM M, ZULFIQAR B, et al. Understanding brassinosteroid-regulated mechanisms to improve stress tolerance in plants: a critical review[J]. Environmental Science and Pollution Research, 2017, 24(19): 15959-15975.
[3] TANG W, YUAN M, WANG R, et al. PP2A activates brassinosteroid-responsive gene expression and plant growth by dephosphorylating BZR1[J]. Nature Cell Biology, 2011, 13(2): 124-131.
[4] YIN Y, VAFEADOS D, TAO Y, et al. A new class of transcription factors mediates brassinosteroid-regulated gene expression in[J]. Cell, 2005, 120: 249-259.
[5] HE J X, GENDRON J M, YANG Y L, et al. The GSK3-like kinase BIN2 phosphorylates and destabilizes BZR1, a positive regulator of the brassinosteroid signaling pathway in[J]. Proceedings of the National Academy of Sciences of the United States of America, 2002, 99(15): 10185-10190.
[6] LI J, NAM K H, VAFEADOS D , et al. BIN2, a new brassinosteroid-insensitive locus in[J]. Plant Physiology, 2001, 127(1): 14-22.
[7] YU X, LI L, ZOLA J, et al. A brassinosteroid transcriptional network revealed by genome-wide identification of BES1 target genes in[J]. The Plant Journal, 2011, 65(4): 634-646.
[8] YE Q Q, ZHU W J, LI L, et al. Brassinosteroids control male fertility by regulating the expression of key genes involved inanther and pollen development[J]. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(13): 6100-6105.
[9] GUO Y F, SHAN W, LIANG S M, et al. MaBZR1/2 act as transcriptional repressors of ethylene biosynthetic genes in banana fruit[J]. Physiologia Plantarum, 2019, 165(3): 555-568.
[10] SAHA G, PARK J I, JUNG H J, et al. Molecular characterization of BZR transcription factor family and abiotic stress induced expression profiling in[J]. Plant Physiology and Biochemistry, 2015, 92: 92-104.
[11] 安汶铠,常丹,张富春. 干旱胁迫下棉花幼苗转录因子BES1/BZR1对外源油菜素内酯的响应表达特征[J]. 西北植物学报,2015, 35(7): 1311-1316.
[12] CUI X Y, GAO Y, GUO J, et al. BES/BZR transcription factor TaBZR2 positively regulates drought responses by activation of[J]. Plant physiology, 2019, 180(1): 605-620.
[13] LV J, LI Y, LIU Z, et al. Response ofgenes to abiotic stress and hormone treatment in[J]. Plant Physiology and Biochemistry, 2020, 151(17): 157-165.
[14] 张国燕,陈子豪,李文君,等. 异源过表达胡杨基因提高烟草对盐胁迫的耐受性[J]. 基因组学与应用生物学,2020, 39(8): 3593-3599.
[15] LI Q, GUO L, WANG H, et al. In silico genome-wide identification and comprehensive characterization of thegene family in soybean[J]. Heliyon, 2019, 5(6): e01868.
[16] 邱爽,张军,何佳琦,等. 大豆肌醇半乳糖苷合成酶基因克隆及非生物胁迫表达分析[J]. 西南农业学报,2021, 34(5): 945-949.
[17] YE H, LIU S, TANG B, et al. RD26 mediates crosstalk between drought and brassinosteroid signalling pathways[J]. Nature Communications, 2017, 8(1): 14573.
[18] 陈旭,沈春洋,莫福磊,等. 番茄基因家族鉴定及非生物胁迫下表达模式分析[J]. 东北农业大学学报,2021, 52(11): 9-17.
[19] 杨瑞瑞,李小双,梁玉青,等. 植物激素在苔藓生长发育与逆境响应过程中的作用机制研究进展[J]. 西北植物学报,2022, 42(3): 527-540.
[20] 唐宁,陈信波. 植物MYB转录因子与非生物胁迫响应研究[J]. 生物学杂志,2014, 31(3): 74-78.
[21] 郭新磊,路普,王园园,等. 棉花基因家族的全基因组鉴定及表达分析[J]. 棉花学报,2017, 29(5): 415-427.
[22] LI H, YE K Y, SHI Y T, et al. BZR1 positively regulates freezing tolerance via CBF-dependent and CBF-independent pathways in[J]. Molecular Plant, 2017, 10(4): 545-559.
Bioinformatics and drought stress expression analysis of sixgenes in soybean
ZHANG Jun1,CHEN Jiong-xin2,YIN Jun-yi1,WANG Huan1,LIU Qiu-jin1,TIAN Qiu-feng1,BAI Chang-sheng1,ZHAI Ying2
(1.Branch of Animal Husbandry and Veterinary of Heilongjiang Academy of Agricultural Sciences, Heilongjiang Qiqihar 161005, China; 2.College of Life Science and Agroforestry, Qiqihar University, Heilongjiang Qiqihar 161006, China)
BES1/BZR1 transcription factors play an important role in brassinosteroid signaling pathway, which regulates plant growth and development and resistance to various stresses. In this study, the expression levels of sixin soybean were detected under drought stress, and the-acting elements in the promoters of sixwere predicted and analyzed. Real-time fluorescent quantitative PCR results showed that the expression levels of sixchanged in different degrees with time under drought stress, among which the expression level ofincreased most significantly. The results of promoter sequence prediction analysis showed thatpromoter sequences contained 3, 6, 4, 3, 4 and 7 types of hormones and stress-related-acting elements, respectively. These results provided a theoretical basis for the study of drought resistance mechanism and application of soybean.
soybean;brassinosteroid;drought stress;promoter;-actingelement
2022-09-04
齐齐哈尔市科技计划创新激励项目(CNYGG-2022021);黑龙江省省属高等学校基本科研业务费科研项目(145109506)
张军(1982-),男,黑龙江双鸭山人,助理研究员,硕士,主要从事分子遗传学研究,307906439@qq.com。
翟莹(1982-),女,吉林吉林人,教授,博士,主要从事植物分子遗传育种研究,fairy39809079@126.com。
S565.1
A
1007-984X(2023)01-0070-05
