黔北上奥陶统宝塔组头足类的聚爆深度分析及其地质学意义
2023-02-27彭庭祖杨宇宁王冬梅池祥日
彭庭祖,杨宇宁,王冬梅,池祥日
贵州大学资源与环境工程学院, 喀斯特地质资源与环境教育部重点实验室, 贵阳,550025
内容提要: 上奥陶统宝塔组是扬子板块分布广泛的标志性地层单元, 因富含类型多样的头足类化石及发育特殊的网纹状构造而闻名。目前, 有关宝塔组沉积时期水体深度的分析都是基于沉积学和古生物学特征的定性推测, 而且争议较大。笔者等在黔北尧龙山地区宝塔组采集到大量原域埋藏的头足类化石, 根据它们的隔壁强度系数 (Septal Strength Index, SSI) 与聚爆深度的耦合关系, 首次对这些头足类生活时的古海水深度进行定量化分析。通过选取符合SSI分析条件的16块Sinoceras和12块Michelinoceras标本, 系统分析了SSI分别与外部特征和内部构造的相关性。结果显示, SSI随壳体长度及隔壁间距的增大而增加, 随顶角角度的增加而减小, 但与内部构造单一参数的相关性不大, 可能受到多个内部构造参数的共同制约。定量化分析结果显示, 尧龙山地区宝塔组中共埋出现的Sinoceras和Michelinoceras的SSI分别为5.564 ~ 10.829和11.432 ~ 19.532, 对应的生态深度大约分别为128~ 169 m和228 ~ 280 m, 表明两属头足动物生活时可能已经出现明显的生态分层现象, 古生物地理区系也出现一定的差异。不过, 两个属内部不同种的生态深度没有明显差别。头足类的生态深度分析结果表明, 研究区宝塔组沉积时期的水体深度可能大于280 m, 远比前人分析的结果深得多。
奥陶纪温和的气候及广布的浅海为海洋无脊椎动物的繁盛和演替提供了良好的条件 (詹仁斌等, 2013), 包括处于食物链顶端的头足类在内的大量海洋生物都在这一时期实现快速的生态扩张。奥陶纪时期的头足类多样性极高、演化速率快、生态类型多样并且出现在各个生态分层空间 (Crick, 1980, 1993; Kröger and Zhang Yunbai, 2009; 张元动等, 2009), 大幅增加了海洋生态系统营养结构的复杂性 (Teichert et al., 1964; Zhang Yuandong et al., 2010; Kröger et al., 2011; Fang Xiang et al., 2019)。奥陶纪大辐射时期, 全球头足类的多样性分别在弗洛早期、达瑞威尔末期和桑比期—凯迪期之交出现3次峰值, 并且总体呈增长趋势, 直至凯迪中晚期后多样性急剧减少 (Kröger and Zhang Yunbai, 2009; 詹仁斌等, 2013)。相比之下, 扬子台地奥陶纪的头足类先后则经历了达瑞威尔期的扩张、凯迪期的复苏以及凯迪晚期至赫南特期的衰落 (Fang Xiang et al., 2019)。
中国奥陶纪的头足类广泛分布在西藏 (赖才根, 1982; 陈挺恩, 1984; 程立人等, 2005; 罗凯等, 2012)、甘肃 (张日东, 1962)、湖北 (方翔等, 2015a)、吉林 (彭玉鲸, 1991)、江苏 (潘正勤, 1986)、辽宁 (王敏成, 1986)、湖南 (赖才根和齐敦伦, 1977)、安徽 (应中锷, 1989)、陕西 (赖才根, 1981) 等地的奥陶纪地层中。其中, 扬子板块保存在碳酸盐岩地层单元中的头足类最为丰富, 比如下奥陶统红花园组 (徐光洪和徐安武, 1988) 和本研究的目标地层上奥陶统宝塔组 (彭庭祖等, 2021) 都发现大量的头足类化石。
头足类在海水中主要是通过体内气室排吸水形成的反作用力调节沉浮和运动 (Flower and Teichert, 1957), 因此其壳壁厚度、壳型、壳体大小、壳面纹饰和内部结构在一定程度上反映头足类的生活方式、游泳能力及生存环境 (赵金科等, 1965;徐光洪和徐安武, 1988; 张永辂等, 1988; 肖传桃等, 2006)。例如, 壳壁厚度越大表明能承受静水压力的强度越大, 生活时的水体较深; 直壳形的头足类相较于平旋壳型更适应深水区域, 且扩大缓慢的直锥形壳能减小对水的阻力从而快速运动; 气室排列稀疏且壳体细小的体管能增大气室体积比率从而更利于运动; 壳面纹饰清晰的化石表明当时应处于低能环境 (张永辂等, 1988; 肖传桃等, 2006)。此外, 头足类在奥陶纪海洋生态系统中处于中心位置 (Kröger and Zhang Yunbai, 2009), 分析这一时期头足类的生活水深有助于了解当时它们的生态分层结构、空间分异以及古水深信息。
现生鹦鹉螺类头足动物的壳体实验结果表明, 当内部气压小于1 bar (压强单位, 10 bar=1 MPa) 时壳体所能承受的静水压力约为70 ~ 90 bar, 对应的水深约在700 ~ 900 m (Bidder, 1962; Mutvei, 1964; Denton and Gilpin, 1966)。相比之下, Westermann (1973) 通过对7个目47个头足动物化石属进行实验分析, 认为直角石目 (Orthocerida) 头足类所能承受的极限深度为150 ~ 500 m。Chen Junyuan和Teichert (1987) 对近3000块头足类化石标本的古生态学研究表明, 头足动物在奥陶纪不同时期逐渐适应不同水深, 且最深可达500 m。上奥陶统宝塔组作为扬子台地的标志性沉积单元之一, 以发育奇特的网纹状灰岩和产出大量头足类动物化石为特征 (戎嘉余和陈旭, 1987; Zhan Renbin et al., 2016), 而宝塔组的成因机制及水体深度长期以来都是学术界争议的焦点 (姬再良, 1985; Chen Junyuan et al., 1991; Lindström et al., 1991; 陈旭等, 1993; Rong Jiayu et al., 1999; 周传明和薛耀松, 2000; Zhan Renbin et al., 2016)。然而, 这些研究绝大部分都是基于沉积学、地球化学或岩相古地理的定性化分析, 却忽视了宝塔组中大量极具特征的头足类化石及其所能揭示的水深条件信息。据此, 笔者等根据在黔北尧龙山地区宝塔组中采集到大量保存完整、分异度和丰度较高以及内部结构清晰的头足类化石, 重点分析了外部形态、内部构造与头足类动物的聚爆深度 (头足类因静水压力过大而发生内爆时, 其下潜所处的临界深度被称为聚爆深度; Denton and Brown, 1973) 和生态深度 (常规生活所处的水体深度; 陈均远, 1988) 的耦合关系, 为探讨华南晚奥陶世头足类的生态分层结构以及宝塔组沉积时期的海水深度提供新信息。
1 地质背景
晚奥陶世时期, 贵州沉积区由于受到因都匀运动抬升的黔中隆起的影响 (崔金栋, 2013; 陈超等, 2014), 仅在黔北、黔东和黔南局部区域发育有完整的奥陶纪地层 (周名魁等, 1993; 周明辉, 2005; 尹琼, 2013; 牟传龙等, 2014)。这一时期海平面的相对上升致使水体自黔中向北逐渐加深, 在黔北地区形成了宽缓的碳酸盐岩沉积环境 (谢尚克等, 2011; 牟传龙等, 2014)。研究区位于贵州省遵义市桐梓县尧龙山镇 (图1a), 该区出露的上奥陶统地层主要为宝塔组、涧草沟组及五峰组。其中, 宝塔组是扬子台地岩性最稳定、分布最广的碳酸盐岩地层单元之一 (周传明和薛耀松, 2000; 许效松等, 2001), 与上覆地层涧草沟组为整合接触、与下伏地层十字铺组可能为不整合接触 (Munnecke et al., 2011; 樊茹等, 2013)。宝塔组岩性为一套灰色、深灰色中—厚层具网纹状构造的泥晶灰岩或生物碎屑泥晶灰岩, 层理极为发育 (图1b), 以产出大量特征性的中华震旦角石Sinoceraschinense而得名。此外, 三叶虫、腕足类、腹足类、牙形石和棘皮类等生物也比较常见, 但缺乏内栖生物化石, 可能是深水环境的营养物质匮乏 (陈旭和丘金玉, 1986; 江茂生, 1998)、沉积环境处于底栖生物生活的透光带之下 (戎嘉余和陈旭, 1987) 或底部氧气不足 (Zhan Renbin and Jin Jisuo, 2007) 等原因所致。牙形石生物地层研究表明, 宝塔组自下而上可识别出2个牙形石带:Hamaroduseuropaeus带和Protopanderodusinsculptus带 (王钢, 1986; 安太庠, 1987; 曾庆銮等, 1987; 王钢, 1990), 其地质时代为晚奥陶世桑比中期—凯迪早期 (Chen Xu et al., 1995; Zhan Renbin et al., 2016)。

图1 黔北尧龙山地区交通位置图 (a); 酒店垭剖面宝塔组地层呈水平状产出(b); 宝塔组内原域埋藏的头足类化石(c) 以及酒店垭剖面宝塔组的岩石地层和生物地层柱状图(d)
扬子台地内宝塔组的沉积厚度变化较大, 厚度通常介于20 ~ 35 m之间 (禚喜准等, 2017), 一般西部和南部近岸浅水环境区域的厚度较大, 而靠近台地边缘的远岸区域其厚度较小 (Zhan Renbin et al., 2016)。黔北尧龙山地区酒店垭剖面的宝塔组发育有101层网纹灰岩, 总厚25.68 m (图1d)。笔者从研究区天域极地、酒店垭和集镇采坑3个剖面的宝塔组采集到近千余件原域埋藏的头足类化石 (营游泳或底栖—游泳的头足类在死亡后由于气室大量充水致使其沉降在同一生态域的海底并被掩埋石化, 未经历异地搬运; 图1c), 共有8属11种及少量属种未定类型 (彭庭祖等, 2021), 以震旦角石属Sinoceras和米契林角石属Michelinoceras为优势分子, 盘角石属Discoceras为常见的特征性分子。尧龙山宝塔组头足类的组合特征与宜昌三峡地区宝塔组的头足类 (Chen Tingen and Zou Xiping, 1984; 徐光洪和徐安武, 1988) 比较相似, 总体上符合深水陆棚Sinoceras—Michelinoceras—Discoceras生物相 (肖传桃等, 2006) 的特征, 不过三峡地区头足类的化石产出层位可能略高于尧龙山地区 (彭庭祖等, 2021)。
2 研究材料
黔北桐梓尧龙山地区的上奥陶统宝塔组中产出丰度和分异度都很高的长锥型头足类化石 (彭庭祖等, 2021), 是进行隔壁强度系数分析的理想材料。本研究在尧龙山地区的酒店垭、天域极地和集镇采坑等3个剖面采集到原域埋藏的近千块头足类化石, 并从中遴选出28件符合SSI分析要求的长锥型标本。其中, 喇叭角石目喇叭角石科震旦角石属Sinoceras标本16块 (图2a—f), 直角石目直角石科米契林角石属Michelinoceras标本12块 (图2g—k)。研究标本均为立体保存, 从平行于壳体的长轴方向切割揭示纵切面。根据SSI的分析要求, 所有研究标本度量或测算9个特征参数, 分别为最小隔壁厚度 (σ)、隔壁半径 (R)、壳长 (L)、顶角大小 (α)、隔壁间距 (△)、体管直径/壳体直径 (g/r2)、隔壁颈长/气室高 (n/d)、隔壁下凹度 (s/d) 和气室高/壳径 (d/r1), 并计算出每个个体的聚爆深度和生态深度 (表1)。由于外壳型头足类至少有14种不同的体管结构, 在静水压力下所承受的机械强度也各不相同, 有学者提出现生鹦鹉螺类隔壁强度系数的推导公式不适用于化石类型 (Mutvei, 2017)。据此, 笔者等只选取具有相同体管结构的Sinoceras和Michelinoceras化石标本作为研究对象, 并从中选择隔壁的保存状态符合SSI分析要求的个体进行分析。需要提及的是, Westermann (1973) 的实验数据不包括生长阶段个体的信息, 为避免测量数据影响隔壁强度的分析结果, 本研究的所有测量参数全部来自成年个体的化石标本 (成年个体都发育有一个基本完整的住室; Hewitt and Westermann,1996)。

图2 黔北尧龙山地区宝塔组的头足类Sinoceras和Michelinoceras

表1 黔北尧龙山地区宝塔组中应用于SSI分析的Sinoceras和Michelinoceras的特征参数度量
3 研究方法
3.1 隔壁强度分析
头足类作为海生软体动物, 最大栖息深度主要由壳体的力学性能决定 (Ward and Boletzky, 1984)。头足类壳体在水中受到四周静水压力的作用且壳体内部压力通常小于101.325 kPa (1 atm), 当外部静水压力超过生物体所能承受的极限而发生内爆时, 其下潜所处的临界深度被称为聚爆深度 (又称极限深度) (Denton and Brown,1973; 陈均远, 1988)。头足类的内爆现象主要与生物体抗压强度最弱的部分有关 (Westermann, 1973; 陈均远, 1988), 而承受海水静水压力的构造主要是壳壁、隔壁和体管壁。
对于外壳型头足类而言, 钙质壳壁 (calcareous shell wall) 包裹所有内部软体组织, 可抵抗四周的静水压力、支撑身体弯曲和抵抗其他生物的意外攻击, 因而是整个身体中最坚硬的部分 (Denton and Brown, 1973; Westermann and Hamilton, 1985)。体管壁 (siphuncle wall) 除了承受周围的静水压力之外, 还是气室内部和体管之间物质交换的场所, 并能固定软体组织, 因此其机械强度往往大于产生内爆现象时的临界强度 (陈均远, 1988)。因此, 体管壁强度可能只对生存环境起到粗略的限制 (Hewitt and Westermann, 2010)。相比于壳壁和体管壁, 隔壁 (septum)是头足类壳体中最脆弱的部分 (Hewitt and Westermann, 2010), 因为大多数浅水生活的头足类的隔壁都比较薄, 当下潜接近最大生态深度时容易受到局部张力而发生内爆现象 (Hewitt and Westermann,1996)。据此, 隔壁是头足类最适合开展聚爆深度分析的内部结构, 其强度系数由隔壁厚度和曲率半径的函数关系所决定 (Westermann, 1973; 陈均远, 1988)。
头足类隔壁的成分为珍珠质, 向口前方凹陷形成半球形。这种弯曲方式可以较大程度上增加住室的空间, 但也会降低隔壁在水中的抗压强度 (陈均远, 1988; Hewitt and Westermann, 1996)。珍珠层的抗张强度无法直接确定, 但在薄壳力学原理中, 半球形隔壁所受到的机械强度与隔壁厚度成正比, 与曲率半径成反比 (陈均远, 1988)。因此, 根据现生乌贼和鹦鹉螺的仿生实验数据, Westermann(1973) 将头足类的抗张强度与聚爆深度相结合, 推导出隔壁强度系数 (Septal Strenth Index,SSI) 的函数公式;SSI=σ/R×1000, 其中σ表示最小隔壁厚度,R表示相应的曲率半径。现生头足类的仿生试验和数值模拟分析显示, 1个单位SSI所代表的聚爆深度为30 m, 对应的生态深度约为聚爆深度的2/3或3/4 (Westermann and Hamilton, 1985)。头足类为了保持隔壁强度的稳定, 曲率半径需随最小隔壁厚度的增加而成比例增加 (Westermann, 1973)。另外, 由于头足类的隔壁通常不是一个标准的半球形, 微小差异的累积会导致抗张强度有所减小。为此, Hewitt (1993) 细化并改进了头足类的隔壁强度指数, 提出当内部气压为1 atm时, 1个单位的SSI对应的水体深度为26.2 m。
对直壳型头足类 (图3a) 而言, 理想的隔壁是一个标准的圆弧型 (图3b), 但有些成年个体的隔壁并不接近这种理想形状 (Westermann, 1977), 进而会降低曲率半径的测量精度。因此, 隔壁强度系数公式不能适用于所有头足类材料, 而且参数的测量精度对分析结果往往有重要影响。因此, 开展SSI分析的先决条件是, 尽量选择隔壁接近标准圆弧且隔壁厚度无异常变化的长锥型壳体, 也不宜选择隔壁因浮力而容易变形的短锥型壳体 (Westermann, 1977)。此外, 成年头足类个体的隔壁厚度有时会出现异常变化, 导致隔壁强度系数存在一定误差 (Hewitt and Westermann, 2010)。为了提高隔壁厚度的测量精度, 本研究选取很少受重结晶或泥晶化作用影响的隔壁 (图3c, d) 进行多位点测量, 最后采纳SSI计算值最小的数值。

图3 直角石类形态复原图 (a); 直角石内部结构参数解析图(b) (改自Westermann and Hamilton, 1985; Fang Xiang et al., 2017); 宝塔组Michelinoceras的隔壁放大图(c)、 (d) , 周围强烈方解石化, 但隔壁的形态和厚度稳定
3.2 数理分析
笔者等选取符合SSI分析条件的头足类化石进行研究, 综合利用标尺实测、Image J和Origin等软件对化石标本进行形态度量和数理计算统计, 数据精度保留至千分位。为了判断这些特征参数与隔壁强度系数SSI的相关程度, 笔者等采用person相关系数 (常用于表征两个变量之间的线性相关性; Stigler and Stephen, 1989) 进行研究。当相关系数值越接近1或-1时, 表明相关性越强; 相关系数越接近于0, 相关性越弱。相关系数的正、负值分别表示两变量之间的正相关和负相关性。
4 宝塔组头足类的聚爆深度分析
4.1 隔壁强度系数 (SSI) 的影响因素
从头足类隔壁强度系数的函数关系 (SSI=σ/R×1000) 可知,SSI的数值取决于最小隔壁厚度 (σ) 和曲率半径 (R) 的变化。现有研究表明, 现生鹦鹉螺的外壳形状与始新世鹦鹉螺类群的外壳形状相似, 但隔壁强度系数却大大降低 (Hewitt and Westermann, 2010), 说明SSI可能与内部结构相关。
此外, 体管形态、连接环形状和隔壁颈类型等内部结构是头足类的主要鉴定特征 (赵金科等, 1965; 徐光洪和徐安武, 1988; 张永辂等, 1988; 童金南和殷鸿福, 2007; 方翔等, 2015b)。为了揭示黔北宝塔组中影响头足类化石SSI的潜在因素, 在形态度量统计的基础上, 本研究分别对外部特征和内部构造与SSI的相关性进行了分析。其中, 外部形态参数包括壳长 (L)、隔壁间距 (△) 和顶角大小 (α), 内部构造包括体管/壳径 (g/r2)、隔壁颈/气室高 (n/d)、隔壁下凹度 (s/d) 和气室高/壳径 (d/r1) 等。
4.2 外部特征 (壳长、隔壁间距和顶角大小) 与SSI的相关性
黔北宝塔组中Sinoceras和Michelinoceras的壳长 (L)、隔壁厚度 (σ) 和顶角大小 (α) 分别对于SSI的相关性分析 (图4) 显示,Sinoceras和Michelinoceras的壳长 (L) 与SSI均具有一定正相关性 (图4a)。壳体越长则SSI值越大, 说明壳体大小可能是影响聚爆深度的关键性因素之一。对于死亡后未经受强烈变化、完整保存生长记录的头足类化石而言, 壳长 (L) 在一定程度上反映了个体的纵向发育情况 (方翔, 2015), 因此大个体的隔壁厚度 (σ) 越大, 可以承受更深的聚爆深度所带来的静水压力。这与现代头足类的仿生实验结果相似, 即大型或成年头足类的聚爆深度远大于小型或未成熟的个体 (Ward, 1982; Westermann and Hamilton, 1985)。相应地, 当壳体长度增加时, 隔壁厚度也随之增加, 而且需要更宽的隔壁间距 (△) 来减少相应的累积重量, 所以SSI值常常随隔壁间距的增大而增大 (图4b)。此外, 在直壳型头足类中, 顶角大小 (α) 与壳体扩大率都反映头足类的生长速率, 当顶角角度越大说明壳体发育越快, 在水中运动时受到的阻力也越大 (徐光洪和徐安武, 1988; 肖传桃等, 2006), 因此SSI值与顶角角度呈明显的负相关 (图4c)。当头足类的气室容量总体积、体积增量、隔壁强度保持不变时, 顶角大小 (α) 常与隔壁间距 (△) 和壳体长度 (L) 呈负相关关系 (Westermann, 1973; 图4d~e)。
综上, 黔北宝塔组头足类Sinoceras和Michelinoceras的外部形态要素中,SSI值均与壳体长度 (L) 和隔壁间距 (△) 为正相关, 与顶角大小 (α) 呈负相关。这一结果与前人关于现生头足类及头足类化石的聚爆深度模拟实验 (Westermann, 1973; Westermann and Hamilton, 1985) 所观测到的现象相同。
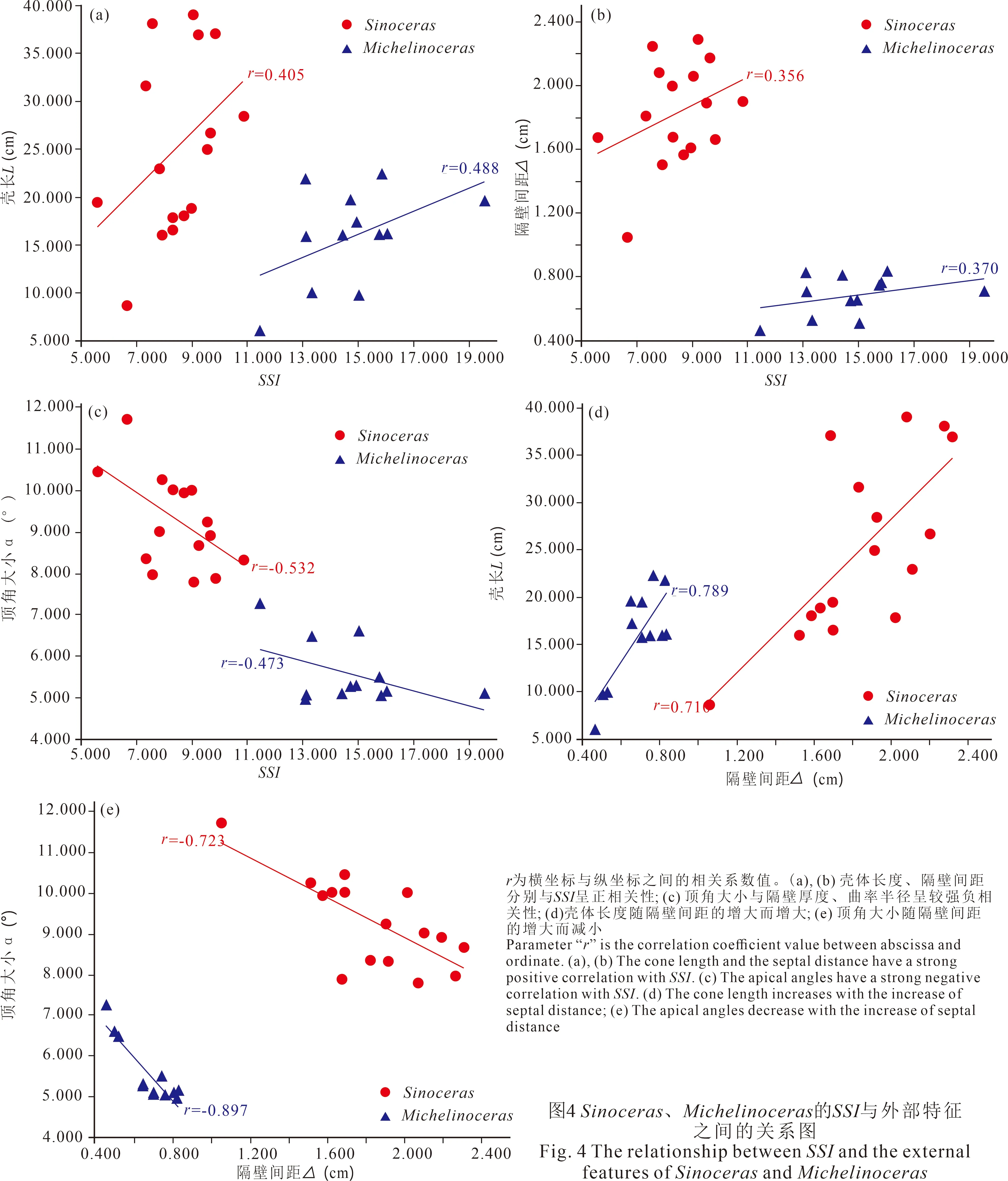
4.3 内部结构 (体管/壳径、颈长/气室高、隔壁下凹度和气室高/壳径) 与SSI的相关性
头足类的内部结构显示, 细小的体管、较短的隔壁颈、较小的隔壁下凹度及较大的气室高/壳径既能减轻生物体的累积重量, 也能增加气室容积比率, 从而提升躯体的游泳能力 (Westermann, 1977)。Sinoceras和Michelinoceras的体管都位于壳体中央, 直径均占壳径的1/10左右; 其中Sinoceras隔壁颈长约为气室高的1/2。相比之下Michelinoceras存在更短的隔壁颈长, 仅为气室高的1/10 ~ 1/5。Sinoceras作为游泳能力较强的长直锥形类型, 隔壁下凹度基本为标准的半圆形, 但Michelinoceras的隔壁下凹度稍大一些。气室高/壳径 (d/r1) 小于0.3表示头足类具有密集的隔壁 (Westermann and Hamilton, 1985),Sinoceras和Michelinoceras的气室高/壳径 (d/r1) 均大于0.3, 表明两者气室都比较稀疏, 游泳能力较强。
Sinoceras和Michelinoceras的内部结构与SSI的相关性分析结果 (图5) 显示,SSI与气室高/壳径呈弱正相关 (0.2 图5 Sinoceras、Michelinoceras的SSI与内部结构参数之间的关系图 头足类作为奥陶纪时期的一类营游泳, 部分营底栖或漂游方式生活 (戎嘉余和陈旭, 1987; 陈均远, 1988; 徐光洪和徐安武, 1988; 张永辂等, 1988; 肖传桃等, 2006) 的海生无脊椎动物, 常具有分布范围广泛和演化速度快的特点。海生的头足类在死亡后数小时内, 内部气室会被快速充满海水而沉落于海底, 因此基本属于原域埋藏 (Collins and Minton,1967; Hewitt and Westermann, 1996; Kröger, 2013; Fang Xiang et al., 2017, 2019)。头足类作为奥陶纪处于食物链顶端的重要标准化石, 它的生态空间和在地层中的分布不仅受到环境条件、生活方式和埋藏条件的影响 (萧传桃和潘云唐, 1997; 肖传桃等, 2006), 也会受到自身壳体所能承受聚爆深度的大小所控制 (Westermann, 1973; Sanders and Landman, 1987; 陈均远, 1988)。 图6显示, 黔北尧龙山地区Sinoceras的SSI值为5.564 ~ 10.829, 中位数为8.985,Michelinoceras的SSI值可达11.432 ~ 19.532, 中位数为14.96, 表明后者的抗聚爆强度明显高于前者。根据头足类1个单位的SSI值约等于26.2 m的聚爆深度 (Hewitt, 1993), 而生态深度大约为聚爆深度的2/3 (Westermann and Hamilton, 1985), 测算出Sinoceras的聚爆深度为146 ~ 284 m, 对应的生态深度约为97 ~ 189 m;Michelinoceras的聚爆深度为300 ~ 512 m, 对应的生态深度约为200 ~ 341 m ( 表1) 。这一结果充分说明, 尽管扬子地区宝塔组中的Sinoceras和Michelinoceras经常共埋于同一个层面之上, 但二者活着时具有明显不同的生态分层空间, 即大多数Michelinoceras可能为了降低生存竞争压力, 在Sinoceras生活水域之下更深的水体中生活。需要提及的是, 研究区宝塔组的S.chinense和S.eccentrica具有相似的生态深度,Michelinoceras的两个种 (M.huangnigangense,M.xuanxianense) 也没有显示出生态分层差异 (表1)。此外, Chen Junyuan (1988) 对湖北宜昌地区宝塔组中Sinoceras和Michelinoceras的聚爆深度分析显示, 两个属的SSI值都在15以上, 平均值为18.2, 明显高于黔北尧龙山地区相同属的SSI值。据此, 尽管有学者提出在相同地形条件下头足类SSI值与属种因素和采样地点无关 (陈均远, 1988; Chen Junyuan, 1988), 但本研究结果显示, 华南上奥陶统宝塔组中的Sinoceras和Michelinoceras存在明显的生态分层现象, 而且在不同海域中其生活的水深可能有所不同, 不过属内不同种之间的生态深度没有明显差异。 图6 Sinoceras和Michelinoceras的最小隔壁厚度 (σ) 与曲率半径 (R) 的双对数函数 (据Westermann, 1973) 关系图, 两个属的SSI区间值分别为5.564 ~ 10.829和11.432 ~ 19.532 研究区与湖北宜昌地区宝塔组中的头足类同属于扬子台地晚奥陶世深水陆棚Sinoceras—Michelinoceras—Disoceras生物相区 (彭庭祖等, 2021)。该生物相区分布范围极广,Sinoceras和Michelinoceras作为其中的代表性分子, 具有扩大相对缓慢、体管细小且近于壳体中央、横截面为圆形且气室稀疏等深水游泳型生态特征 (肖传桃等, 2006)。化石记录显示,Sinoceras和Michelinoceras作为宝塔组沉积时期头足动物的优势类群, 二者在扬子板块内部的陆棚环境和东南缘的深水陆棚—半深水环境中经常呈共生状态出现 (图7)。此外, 一些Sinoceras会分布在南缘的陆棚环境中, 而少数Michelinoceras则单独出现在东南缘的前陆盆地西缘 (图7)。这表明, 在生态竞争压力下, 华南宝塔组中外形比较相似的Sinoceras和Michelinoceras虽然生活时经常共享相同的陆棚海域, 但二者已经出现明显的生态分层, 而且Sinoceras和Michelinoceras开始分别零星向台地南缘和东南缘的前陆盆地迁移扩张 ( 图7) 。 图7 华南中奥陶世—晚奥陶世早期岩相古地理图 ( 改自马永生等, 2009)及Sinoceras与Michelinoceras的生物地理区系图 (仅限宝塔组沉积期; 化石记录来源于张日东, 1964; 赖才根, 1965,1980; 赖才根和齐敦伦, 1977; Chen Tingen and Zou Xiping, 1984; 齐敦伦, 1984; 沈建伟, 1988; 应中锷, 1989; 萧传桃和潘云唐, 1997; 方翔等, 2015a) 奥陶纪时期是显生宙以来全球海平面最高的时期之一 (Nielsen, 2004; Miller et al., 2005; Munnecke et al., 2010), 而宝塔组沉积于扬子台地水深最大的时期 (奥陶纪桑比中期—凯迪早期; 陈均远, 1988; 江茂生, 1998)。然而迄今为止, 学术界关于宝塔组沉积时的水体深度基本都是定性推测而且争议较大。比如, 戎嘉余和陈旭 (1987) 根据宝塔组富含浮游型三叶虫Cyclopygidae、游泳型头足类Sinoceras和Michelinoceras以及推测营假漂浮生活的腕足类等化石, 认为宝塔组的沉积水体深度可能超过浪基面 (约100 m)。Fortey (1997) 根据宝塔组中产出的底栖三叶虫眼睛较大的特征, 判断沉积环境位于透光带内, 深度可能不足200 m。陈均远等 (1991) 和Lindström等 (1991) 根据宝塔组的沉积特征和生物化石, 认为水体深度可能大于200 m。Rong Jiayu等 (1999) 认为宝塔组中的Leangella—Foliomena腕足动物组合常居住在150 ~ 200 m之间。Zhan Renbin等 (2016) 认为华南上奥陶统宝塔组沉积于赤道附近没有显著构造活动的环境中, 宝塔组大型瘤状灰岩沉积时的海水深度不会超过120 m。需要提及的是, 李志明等 (1997) 等曾测算出湘西北地区宝塔组中Sinoceras的SSI值为29 ~ 45, 并判断水体深度在100 ~ 200 m之间。但是, 即使李志明等 (1997) 关于Sinoceras的SSI值的测算结果无误, 那么对应的生态深度至少为600 m, 而不是其所推测的小于200 m。 本研究通过对研究区宝塔组中Sinoceras和Michelinoceras的聚爆深度和生态深度进行量化分析。研究结果显示,Sinoceras和Michelinoceras的生态深度分别为97 ~ 189 m (中位数为148.5 m) 和200~341 m (中位数为259 m)。这一结果大致处于前人关于直角石类化石古水深分析的合理区间 (150 ~ 500 m; Westermann, 1973; Chen Junyuan and Teichert, 1987 ) 。为了消除由于取样偏差导致的极端值, 分别去除2个属生态深度的最小值和最大值之后, 则Sinoceras和Michelinoceras的生态深度范围分别为128~169 m和228~280 m。据此, 尽管同一种头足类在不同海域的生活水深可能不尽相同, 鉴于沉积水深一般不小于游泳生物的最大生态深度, 表明黔北尧龙山地区宝塔组沉积时期的海水深度可能比之前大多数学者预想的更深 (≥280 m), 支持陈均远等 (1991) 和Lindström等 (1991) 关于宝塔组沉积水深大于200 m的推测结果。 宝塔组缺乏任何指示动荡水体的沉积构造、风暴岩或生物遗迹 (Zhan Renbin et al., 2016) , 无眼的圆尾虫类三叶虫 (Cyclopyge) 和叶月贝腕足类(Foliomena)的出现 (Rong Jiayu et al., 1999) 也指示一个相对稳定的深水环境。不过, 从岩性组合来看, 整个扬子台地的宝塔组都是一套大型瘤状灰岩, 其下伏和上覆的岩组均为正常陆棚环境沉积的碳酸盐岩。因此, Zhan Renbin等 (2016) 认为宝塔组是稳定碳酸盐沉积盆地的产物, 沉积水深总体应该位于浪基面之上 (小于120 m) , 远小于本文的分析结果。可见, 由于分析角度、研究材料和证据的差异, 在缺乏定量化数据的前提下, 有关宝塔组沉积水深的争议一直很大。此外, 扬子台地内不同区域宝塔组的沉积厚度、沉积特征和生物组合面貌都有所不同 (Zhan Renbin et al., 2016; 禚喜准等, 2017) , 黔北尧龙山地区宝塔组的沉积水深大于280 m的定量化分析结果, 可能指示研究区位于靠近黔中古陆北缘一个比较特殊的深水环境。 致谢:审稿人提出非常重要的修改意见, 编辑帮助提升论文的写作水平; 贵州大学硕士研究生罗雪绘制了直角石形态复原图, 在此一并表示衷心感谢!
5 讨论和结论
5.1 宝塔组头足类聚爆深度分析的古生态学意义


5.2 对于分析宝塔组沉积时期古水深的启示
