桃果肉组织异常木栓化现象浅析
2023-02-23颜少宾周平郭瑞金光
颜少宾 周平 郭瑞 金光
DOI:10.20023/j.cnki.2095-5774.2023.05.003
收稿日期:2023-01-16
基金项目:福建省属公益类科研院所基本科研专项(2021R1028008、2021R10280011、2022R1028009);财政部和农业农村部国家现代农业产业技术体系资助项目(CARS-30);福建省农业科学院省级種质资源保护单位建设专项;福建省农业科学院引导性科技创新项目(YDXM2023001);福建省农业科学院科技创新团队项目(CXTD2021009-2);福建省人民政府与中国农业科学院农业高质量发展超越“5511”协同创新工程项目(XTCXGC2021019-GSS01)
作者简介:*为通讯作者:金光(1965-),男,副研究员,硕士,主要从事果树栽培研究工作,E-mail:jinguang0591@163.com。颜少宾(1988-),女,助理研究员,硕士,主要从事落叶果树种质资源和果实品质研究工作,E-mail:ysb2010_good@163.com
摘要要:【目的】近年来,在福建产区发现桃果肉组织出现异常木栓化的现象,对此现象进行观测研究,为该症状的诊断及其防控技术的研发提供理论参考。【方法】采集果肉组织异常木栓化的晚熟‘红花鹰嘴桃果实进行症状观察、病原菌培养、矿质营养元素检测和组织切片染色。【结果】桃果肉组织异常木栓化早期果皮无明显症状,去皮后可见果肉白斑;晚期果皮可见黄斑、凹陷,果肉组织呈海绵状且褐化加剧。果肉异常木栓化组织经PDA培养基培养,未产生菌丝亦无明显菌落形态且无异味;果肉异常木栓化组织的Ca、Zn、N等矿质营养元素含量显著低于健康组织,而P、B、Mo等矿质营养元素含量显著高于健康组织;荧光检测技术诊断显示果肉异常木栓化组织的木栓质累积加强。【结论】桃果肉组织异常木栓化可能是一种果实生理性病害,其果肉异常木栓化组织的Ca等矿质营养元素含量降低,而木栓质累积增加。
关键词:桃;果肉;矿质营养元素;木栓质
中图分类号:S662.1 文献标识码:A 文章编号:2095-5774(2023)05-0334-06
Analysis of Abnormal Suberification in Peach Fruit Tissue
Yan Shaobin,Zhou Ping,Guo Rui,Jin Guang*
(Fruit Research Institute,Fujian Academy of Agricultural Sciences,Research Centre for Engineering Technology of Fujian Deciduous Fruits,Fuzhou,Fujian 350013,China)
Abstract:【Objective】 In recent years,the phenomenon of abnormal suberification in peach mesocarp was found in Fujian production. The observation and study of this phenomenon provided theoretical references for the diagnosis of this symptom and the research and development of its prevention and control technology. 【Method】 Late maturing red-flower peach‘Yingzuifruits with abnormal suberification were collected for symptom observation,pathogen cultivation,mineral nutrient detection,and tissue sectioning staining. 【Result】 There were no obvious symptoms in the exocarp at the early stage of suberification,and white spots could be seen in the mesocarp after peeling. In the late stage,the exocarp was macular and sunken,and the mesocarp tissue was spongy and browning was intensified. After the abnormal suberification tissue was cultured on PDA medium,there were no mycelia,no obvious colony morphology and no odor. The content of Ca,Zn and N in suberification tissue was significantly lower than that in healthy tissue,while the content of P,B and Mo was significantly higher than that in healthy tissue. The fluorescence detection technique showed the accumulation of suberin in the suberification tissue. 【Conclusion】 Abnormal suberification tissue of peach fruit may be a physiological disease,the content of Ca and other mineral nutrients in abnormal suberification tissue of peach mesocarp decreased,while the accumulation of suberin increased.
Key words:Peach;Fruit mesocarp;Mineral nutrient element;Suberin
桃原产于中国,栽培历史悠久,其果实外观艳丽、果肉滋味香甜且营养丰富,深受大众的喜爱。桃树适应力强,经济效益高,在我国各生态区均有栽培,栽培面积和产量均居世界首位。至2021年,福建省桃种植总面积1.21万hm2,总产量1.70万t,是重要的经济果树[1]。在南方地区,桃树病害种类多、数量大、难以控制,严重影响桃的产量、质量和经济效益[2],加上桃园的管理水平参差不齐,在栽培过程中病害的发生也日益凸显。
近年来,在福建桃产区发现桃果肉组织呈异常木栓化的现象。据果农反映,该果肉组织异常木栓化(亦称白斑症)发生率高达60%~70%,在晚熟桃上表现最为严重,尤其是‘红花鹰嘴桃。由于果肉组织异常木栓化在早期未有明显症状,待发展到后期,果实完全失去商品价值,造成果农重大经济损失,严重影响了桃产业的健康发展。经资料查阅发现,桃果肉组织异常木栓化症状与芒果、梨等果实生理性病害有相似之处。据国内外研究报道,果实生理性病害的发生与矿质营养元素有关,如果实中K/Ca或N/Ca比值失调会诱发芒果海绵组织生理性病[3];Ca等矿质营养元素含量的减少易诱发酥梨果肉木栓化生理性病害[4]。此次发现的桃果肉组织异常木栓化现象的发生是否与矿质营养元素失调有关尚不明确,导致生产上缺乏科学、有效的防控技术措施,极大地影响了桃产业持续良好的发展。亟需对桃果肉组织异常木栓化开展调研分析,旨在摸清果肉组织异常木栓化的发生规律,探明发生因素,为研发防控技术和解决产业问题提供参考。
1材料与方法
1.1试验材料
于2022年8月10日,在福建省宁德市古田县平湖镇桃示范基地(118.80°E,26.75°N),选取树体健壮、长势较一致的6 a生红花鹰嘴桃,采摘无明显病虫害、无机械损伤、大小均匀、色泽基本一致、八成熟的果实带回实验室,去果皮后对果肉组织异常木栓化果(果肉呈白斑或组织海绵状褐化)和健康果(果肉无明显症状)进行分类筛选备用。
1.2试验方法
1.2.1病原菌培养
取果肉组织异常木栓化果的木栓化与非木栓化交界处的组织置于PDA培养基上培养,每果取3块长宽厚分别为1.0、0.2、0.2 cm的长条形组织,置于同一培养皿中,3个生物学重复,于28℃暗培养一周,待分离物长出后,经光学显微镜镜检和形态学观察进行初步判断。
1.2.2矿质营养元素检测
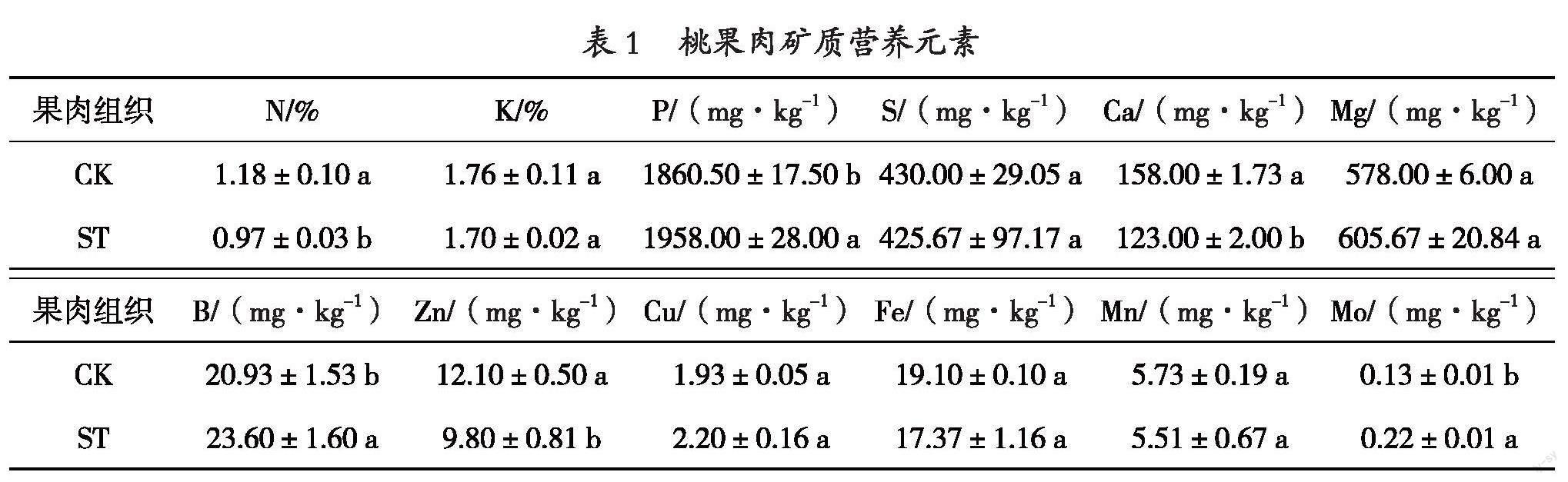
从去果皮的1.1试验材料中,分别选取10个健康果的健康組织(CK)、10个果肉组织异常木栓化程度较一致的果实的木栓化组织(ST),参照GB 5009.268-2016[5]、DZ/T 0253.1-2014[6]等标准,采用电感耦合等离子体质谱仪、电感耦合等离子体发射光谱仪和自动凯氏定氮仪等仪器进行桃果肉矿质营养元素(B、Ca、Cu、Fe、K、Mg、Mn、Mo、N、P、S、Zn)的检测。3个生物学重复。
1.2.3组织切片染色
采用1.2.2试验材料,分别切块,并用FAA固定液固定24 h以上,参考冯丽云等[7]方法进行脱水-浸蜡-包埋-切片-烤片脱蜡复水,再进行组化-FY染色-苯胺蓝染色-封片。由于环芳烃有机染料荧光黄(FY)是亲脂性荧光素,可与木栓质结合后呈绿色荧光,在采集图片过程中进行多通道荧光拍照,最后叠加多通道荧光信号,能使桃果肉的木栓质显示黄色荧光。
1.3数据处理
采用SPSS 24.0和Excel 2007等软件进行统计分析。
2结果与分析
2.1桃果肉组织异常木栓化症状
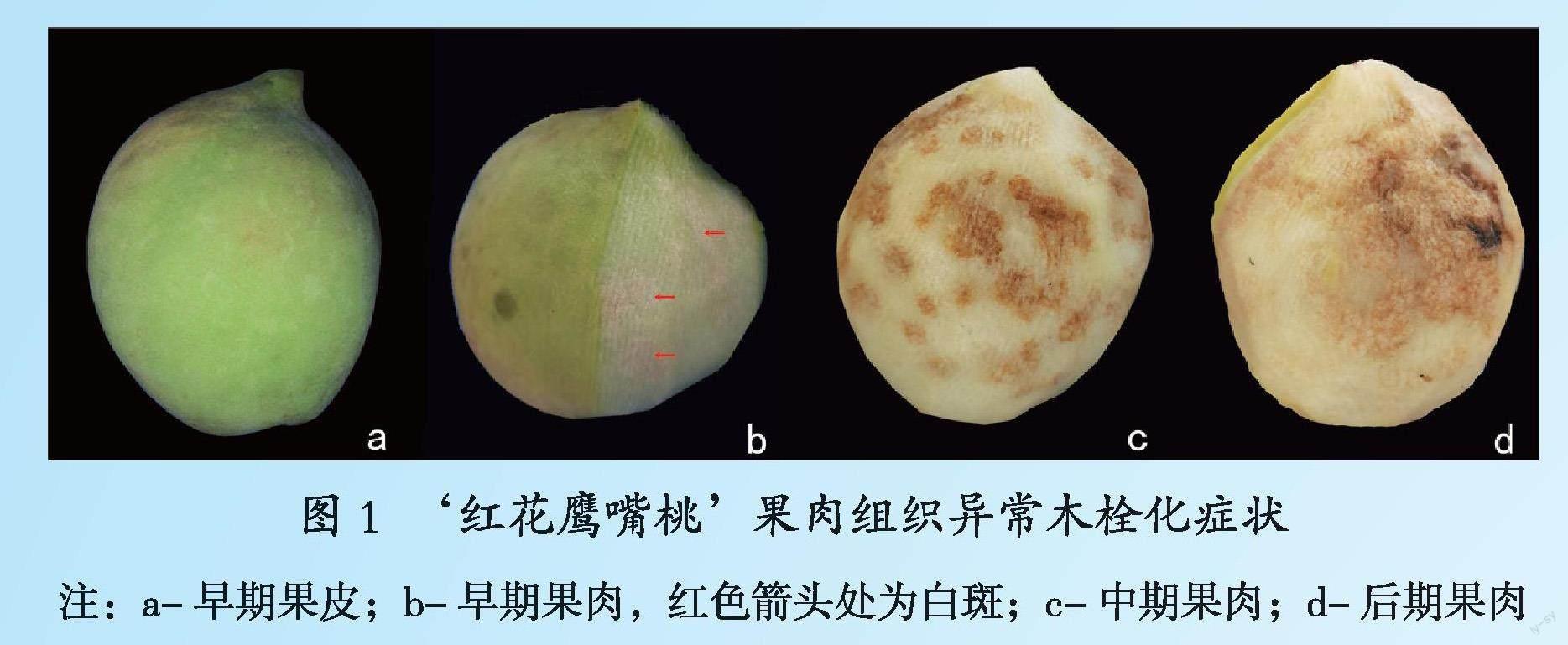
对桃果肉组织异常木栓化调查,结果发现:此症状多发于7、8月份,于果实成熟前15 d左右才明显显现,在晚熟桃上表现最为严重。发生早期果皮无明显症状(封二 图1a),去果皮后果肉可见白斑(封二 图1b);中期果皮出现异常黄斑,果肉组织转为海绵状且褐化(封二 图1c);晚期果皮的黄斑明显凹陷,果肉组织海绵状与褐化加剧,多个斑点黏连呈不规则大斑块,甚至可布满整个果实(封二 图1d)。该症状在早期从外观无法进行诊断,而晚期完全失去经济价值。
2.2桃果肉组织异常木栓化的病原菌培养
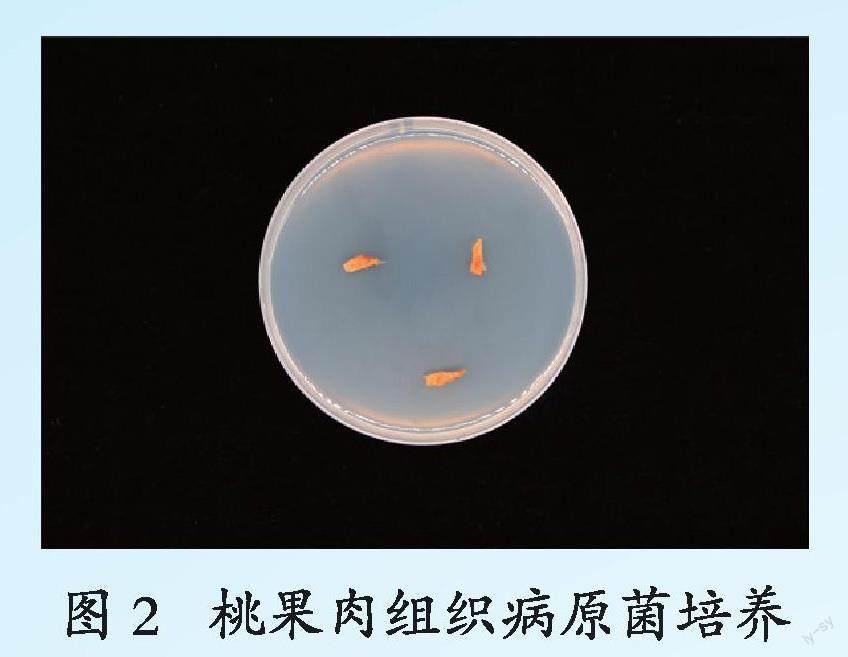
从红花鹰嘴桃果肉组织异常木栓化果实中分离出木栓化与非木栓化交界处组织,置于PDA培养基上培养,该组织不会腐烂且无异味,不会进一步扩散或侵染周边健康组织;在PDA培养基上也未产生菌丝亦无明显菌落形态(封二 图2),说明桃果肉组织异常木栓化可能与微生物致病菌无关,推测可能是一种果实生理性病害。
2.3桃果肉组织异常木栓化的矿质营养元素含量
桃果肉组织异常木栓化的症状与梨的木栓果、芒果的海绵组织[8-11]等生理性病害有类似之处,又和已报道的桃裂果、裂核、异常早熟[12]等生理性病害不同,有研究认为此类果实生理性病害的发生与矿质营养元素有关[13,14]。本研究对桃果肉组织进行矿质营养元素检测,结果发现异常木栓化组织(ST)的Ca、Zn、N等矿质营养元素含量显著低于健康组织(CK);但其P、B、Mo等矿质营养元素含量显著高于健康组织;其它矿质营养元素含量差异不显著(表1)。说明桃果肉组织异常木栓化的发生可能与矿质营养元素的失调有关。
2.4桃果肉组织异常木栓化的组织切片
果实生理性病害的主要特征是细胞解体,导致果肉絮化或产生空洞。木栓质作为细胞壁特异性的杂聚物,为植物细胞提供保护屏障功能[15]。当植物细胞受到外界环境胁迫刺激时,可诱导木栓质的沉积[16]。本研究采集桃果肉异常木栓化组织进行木栓质染色,荧光检测表明(封二 图3),与CK相比,果肉异常木栓化组织(ST)的木栓质可见明显绿色荧光;多通道荧光信号叠加后,与CK相比,果肉异常木栓化组织(ST)的木栓质呈更强的黄色荧光(封二 图4)。这说明果肉异常木栓化组织木栓质的累积显著高于健康组织,有可能是受外界环境胁迫诱导所致。使用木栓质荧光检测技术可以进一步优化,作为桃果肉异常木栓化的诊断方法。
3小结与讨论
本研究结果表明,桃果肉组织异常木栓化是在桃产区新发现的果肉组织异常化现象,取木栓化组织在PDA培养基上培养未产生菌丝、无明显菌落形态且不产生异味,推测桃果肉组织异常木栓化的发生可能与微生物致病菌无关,可能是一种果实生理性病害,但目前证据尚不足,还有待今后进一步验证。此外,在PDA培养基上无法产生菌丝或菌落,也有可能是培养基配方不适、培养环境或培养方式不佳所致,有待今后对致病菌分离的方式方法进行优化验证。田间调查发现,红花鹰嘴桃果肉组织异常木栓化发生的严重程度与其在树体上的坐果位置无直接相关,尤其与是否阳面果、裂果无直接相关,与廖永林等[17]报道的‘鹰嘴桃果实海绵组织生理性病害不同。
此次发现的桃果肉异常木栓化可能与矿质营养元素失调有关。果实生理性病害的发生是由非生物因素作用引起生理代谢失调而诱发的果实病害,施肥不当导致矿质营养元素过量、不足或失调就是非生物因素的一种[18]。前人研究表明,果肉N/Ca高的芒果果实易发生理性病害[13],B、P双缺会促进苍溪梨褐斑病的发生[4];Ca、Mg、B等含量的减少易诱发龙泉酥梨果肉木栓化生理性病害[19],说明果实生理性病害的发生与矿质营养元素有关。施用石灰或0.3%氯化钙与0.3%硼砂混合液能减轻梨、芒果生理性病害的发病率[14,20],进一步证实果实生理性病害的发生与矿质营养元素的失调有关,尤其是Ca含量的缺失。在果实成熟期,若蒸腾作用减弱、代谢生理活动减缓,果实吸收的矿质营养元素会部分回流到其他器官中,进一步减少了果实内Ca含量[21,22]。本研究结果也表明,桃果肉异常木栓化组织的Ca、Zn、N等矿质营养元素显著低于健康组织,推测桃果肉组织异常木栓化的发生可能与矿质营养元素有关。也有学者持不同观点,认为果实病害组织Ca元素缺乏,但在土壤和叶片外源施用Ca等矿质营养元素并不能降低病害的发病率[3,19],可能病害组织的Ca含量与发病无显著相关[22]。而桃果肉异常木栓化的发生是因施肥不当土壤含Ca量低导致桃果实Ca等矿质营养元素吸收不足引起的,还是因品种特性果实蒸腾速率更低、更易感病,又或是因夏季环境胁迫诱发导致的,目前尚无充分证据。此外,若桃果肉组织异常木栓化的发生与矿质营养元素直接相关,是单一Ca元素失调还是多种元素相互作用引起的也尚未可知,有待进一步研究。
桃果肉组织异常木栓化的发生促进了果肉组织木栓质累积。当植物受到外界环境胁迫时,存在于植物质膜和细胞壁之间的木栓质的生物合成被激发大量累積,通过沉积木栓酚类化合物来增强细胞壁结构形成保护屏障[23-25],从而发挥其防御作用。当植物出现抗性性状时,与防御反应相关的基因表达水平未见明显变化,但木栓质含量随之增加,且与木栓质合成相关的基因表达水平也大幅提高,进一步证实木栓质在植物抵御外来胁迫方面起着重要的作用[26]。植物木栓质的生物合成主要有木栓质脂肪族和芳香单体的生物合成等两个途径,已鉴定出β-酮脂酰辅酶A合酶(KCS),细胞色素P450单加氧酶(CYP)家族的脂肪酸氧化酶,脂肪酸酰基还原酶(FAR),甘油-3-磷酸酰基转移酶家族的酰基转移酶(GPAT)和羟基肉桂酰辅酶A转移酶(ASFT或HHT)等参与了该过程[15];AtKCS2/20基因参与木栓质前体的延伸过程[27],AtCYP86A1基因在促进短链脂肪酸的羟基化方面起着重要作用[28],FAR1/4/5基因参与木栓质相关脂肪醇的合成[29],GPAT5/7基因会影响木栓质的形成,进而降低植物的抗逆能力[30]。此外,PpyMYB144可通过正向调控结构基因PpyCYP86B1进而导致木栓质的大量积累形成果锈[31]。而桃果肉组织异常木栓化的生物合成机制目前尚不明确,具体是何种因素诱导其大量生物合成还有待进一步研究。
参考文献:
[1]福建省统计局.2022年福建省统计年鉴[M]. 北京:中国统计出版社,2022:263,270.
[2]李红松,黄婕.南方桃树主要病虫害发生与综合防治[J].广西植保,2007(4):26-29.
[3]Wainwrigth H,Burbage M B. Physiological Disorders in Mango (Mangifera indica L.) Fruit[J]. Journal of Horticultural Science,1989,64 (2):125-135.
[4]张力田,刘先琴,李育民,等.苍溪梨果实褐斑病研究[J].果树科学,1999(1):47-50.
[5]中华人民共和国国家卫生和计划生育委员会,国家食品药品监督管理总局.GB 5009.268-2016食品安全国家标准食品中多元素的测定[S].北京:中国质检出版社,2016.
[6]中华人民共和国国土资源部. DZ/T 0253.1-2014 生态地球化学评价动植物样品分析方法 第1部分:锂、硼、钒等19个元素量的测定[S].北京:中国质检出版社,2014.
[7]冯丽云,张春芬,聂园军,等.苹果花药石蜡切片制片技术改良及其解剖学观察[J].中国农学通报,2016,32(31):62-67.
[8]董肖昌,王宏伟,魏树伟,等.山东省中西部梨果实果肉木栓化褐变的成因及防治[J].果树学报,2018,35(增刊):139-142.
[9]刘家琪,程寅胜,张绍铃,等.早酥梨果实木栓化褐变与矿质营养的关系[J].山西果树,2018(2):1-4.
[10]唐志鹏,武英霞,黄晓容.几种矿质元素对紫花杧果实海绵组织的影响[J]. 果树学报,2005,22(5):505-509.
[11]唐志鹏,王惠,李明富,等.紫花杧果实生理病害海绵组织发生规律研究[J].安徽农业科学,2012,40(14):8107-8110.
[12]刘进,丁志明,李永章,等.桃果实3种生理性病害的发生及防治[J].果树实用技术与信息,2020(3):31-32.
[13]Murthy S K. Chemical Studies on Internal Breakdown in Alphonso Mango (Mangifera indica Linn.) [J].Journal of Horticultural Science,1981,56(3):247-250.
[14]王璐.早酥梨果实木栓化褐变发生规律及防治试验[J].中国果树,1990(1):35-36.
[15]Vishwanath S J,Delude C,Domergue F,et al. Suberin: Biosynthesis,Regulation,and Polymer Assembly of a Protective Extracellular Barrier[J]. Plant Cell Reports,2015,34(4):573-586.
[16]Kolattukudy P E. Polyesters in Higher Plants [M]. In: Babel W,Steinbüchel A. (eds) Biopolyesters. Advances in Biochemical Engineering/Biotechnology,Springer,Berlin,Heidelberg,2001,71:1-49.
[17]廖永林,王龙江,章玉苹,等.鹰嘴蜜桃果肉海绵组织病害调查[J].安徽农业科学,2016,44(33):123-124.
[18]魏森林,张柯,丁向阳,等.常见果树生理性病害与防治[J].河南林业科技,2015,35(2):49-51.
[19]王强,杨佳文,李根,等.龙泉酥梨果肉木栓化褐变的成因及预防措施[J].南方农业,2022,16(13):72-75.
[20]Limaye V P,Gungate R T,Joshi G D. Studies on Occurrence of Spongy Tissue in Some Varieties of Mango[J]. Dapoli Agricultural College Magazine,1975,10:36-37.
[21]张卿,邓西民.不同发育期京白梨果实蒸腾速率的测定[J].园艺学报,2006(2):360-362.
[22]高启明.扁桃幼果生长发育规律及其矿质元素和内源激素含量变化的研究[D].乌鲁木齐:新疆农业大学,2006.
[23]张妍,葛颜锐,赵冉,等.木栓质的结构组分、生物合成及其功能的研究进展[J].科学通报,2022,67(9):822-833.
[24]Lulai E C,Suttle J C,Pederson S M. Regulatory Involvement of Abscisic Acid in Potato Tuber Wound-healing[J]. Journal of Experimental Botany,2008,59(6):1175-1186.
[25]Ranathunge K,Schreiber L,Franke R. Suberin Research in the Genomics Era-New Interest for an Old Polymer [J].Plant Science,2011,180(3):399-413.
[26]Thangavel T,Tegg R S,Wilson C R. Toughing It Out-disease-resistant Potato Mutants Have Enhanced Tuber Skin Defenses[J]. Phytopathology,2016,106(5):474-483.
[27]Lee S B,Jung S J,Go Y S,et al. Two Arabidopsis 3-ketoacyl CoA Synthase Genes,KCS20 and KCS2/DAISY,are Functionally Redundant in Cuticular Wax and Root Suberin Biosynthesis,but Differentially Controlled by Osmotic Stress[J]. The Plant Journal,2009,60(3):462-475.
[28]Benveniste I,Tijet N,Adas F,et al. CYP86A1 from Arabidopsis thaliana Encodes a Cytochrome P450-dependent Fatty Acid Omega-hydroxylase[J]. Biochemical and Biophysical Research Communications,1998,243(3):688-693.
[29]Domergue F,Vishwanath S J,Joubes J,et al. Three Arabidopsis Fatty Acyl-coenzyme a Reductases,FAR1,FAR4,and FAR5,Generate Primary Fatty Alcohols Associated with Suberin Deposition[J]. Plant Physiology,2010,153(4):1539-1554.
[30]Li Y H,Beisson F,Koo A J K ,et al. Identification of Acyltransferases Required for Tutin Biosynthesis and Production of Cutin with Suberin-like Monomers[J]. Proceedings of the National Academy of Sciences of the United States of America,2007,104(46):18339-18344.
[31]張伊凡.PpyMYB144控梨果锈木栓质形成的机制研究[D]. 扬州:扬州大学,2022.
(责任编辑:许 玲)
