塞罕坝地区褐柳莺巢址选择及其对繁殖成功率的影响
2023-02-22汪挥胜祃来坤王姣姣侯建华
汪挥胜,祃来坤,2,王姣姣,侯建华
(1.河北大学,保定,071000;2.河北民族师范学院,承德,067000)
巢址选择是鸟类栖息地选择的重要组成部分,是决定鸟类种群生存繁衍的主要因素之一[1]。巢址选择受到环境中不同生物和非生物因素的综合影响,好的巢址能最大限度地提高亲鸟适合度和巢存活率[2-4]。目前的研究表明,影响巢址选择的主要因素包括微生境、气候因子、食物资源、种内种间竞争、寄生风险、巢捕食和人为干扰等[5-8],其中巢捕食可以直接影响卵、雏鸟和成鸟的生存,被认为是导致鸟类繁殖失败的主要原因[9-11]。巢的可见性和巢捕食的风险紧密相关,因此鸟类多选择隐蔽的地点筑巢,以实现较高的繁殖成功率[12-14]。
了解繁殖策略变异的原因和后果是鸟类生活史研究的焦点[15-16],对于广布种来说,繁殖策略存在地理变异已被人们所熟知[17-19]。另外即使较小的纬度差异,森林类型、鸟类的群落和捕食者的物种群落构成不同,也会导致鸟类面临不同的捕食压力,最终也会影响鸟类的繁殖策略[20-23]。
褐柳莺(Phylloscopus fuscatus)为古北种,在塞罕坝地区属于夏候鸟,常栖息于滦河岸边不同盖度和高度的灌木丛中,种群密度很大[24]。目前,有关褐柳莺繁殖生态的研究很少,仅有1 篇关于俄罗斯远东地区褐柳莺应对花鼠(Tamias sibiricus)捕食的反捕食策略研究,Forstmeier 等[25]研究发现褐柳莺面对不同的捕食风险时能对巢址选择进行调整,表明褐柳莺巢址选择行为具有可塑性。为了增加对低纬度地区以及不同森林类型中褐柳莺巢址选择的了解,进一步探讨褐柳莺巢址选择的可塑性和适应性变异,本研究对河北塞罕坝国家森林公园中的褐柳莺巢址选择及其对繁殖成功率的影响进行研究。
1 研究区概况
塞罕坝国家森林公园位于河北省最北端(42°2'—42°36' N,116°51'—117°39' E),海拔1 350~1 650 m,西北面与内蒙古接壤,是河北主要的天然次生林和人工林区,有高原、山区、森林和草原。植被群系主要包括落叶针叶林、常绿针叶林、针阔混交林、阔叶林、灌丛或灌草丛、草原与草甸等。该地区属寒温带大陆性季风气候,年平均气温-1.4 ℃,每年60 d 无霜,有雪可达7 个月[26]。研究区人工林地主要由针叶林和苗圃地组成,其中针叶林疏密均匀,郁闭度大,林下灌木层和草本层植物稀少;苗圃地疏密不均,郁闭度大,主要由云杉(Picea asperata)和樟子松(Pinus sylvestrisvar.mongolica)幼苗组成,苗高0.5~3.0 m,二者呈镶嵌分布,形成具有郁闭度差异的生境。天然植被主要为北沙柳(Salix psammophila),高3~4 m,下端树叶较少,植被间距大。
2 研究方法
2.1 搜寻巢与监测
2021 年5—8 月,在研究区内通过跟踪处于繁殖阶段的成鸟和“地毯式”搜索寻找褐柳莺巢[27],随后使用GPS(集思宝R600,中国)定位。利用微型摄像机(HiLEME,中国)和定期人工检查的方法对巢监测,直至巢失败或者雏鸟离巢结束。褐柳莺巢为球形且巢口开在侧面,将摄像头固定在距离巢口10~20 cm的树枝上,摄像机设置为全天录制模式。整个繁殖季共找到30 个褐柳莺巢,对13 个视频监测,其他巢隔天检查。记录每个巢的命运(至少有1 只雏鸟离巢视作繁殖成功,否则,视为繁殖失败)以及繁殖失败的原因[28]。
2.2 巢址参数
测量巢树1 m 半径范围的巢址和植被参数,以及巢树5 m 半径范围微生境的植被参数[29]。(1)巢隐蔽度:测量参考Hoover 等[30]的方法,在1 张10 cm×10 cm 的纸上画10 个相同的栅格,用该纸挡住巢的各个方向,然后在距巢1 m 处观察并估测纸被植被遮挡的比例。(2)植被参数:即5 m 微生境范围的植被参数,以巢为中心,东南西北4 个方向距巢2.5 m处各取1 个1 m×1 m 的样方进行测量,再计算平均值。(3)植被盖度:采用目测法估算,巢距地面高和低于2 m 植被高采用卷尺测量,高于2 m 的植被用7 m长的伸缩杆测量,杆上每隔1 m 用鲜艳的色带标记[31]。(4)距道路和距林缘距离:通过GPS Toolbox 中的谷歌卫星地图确定。为避免人为误差,所有的参数测量均由同一人完成(表1)。
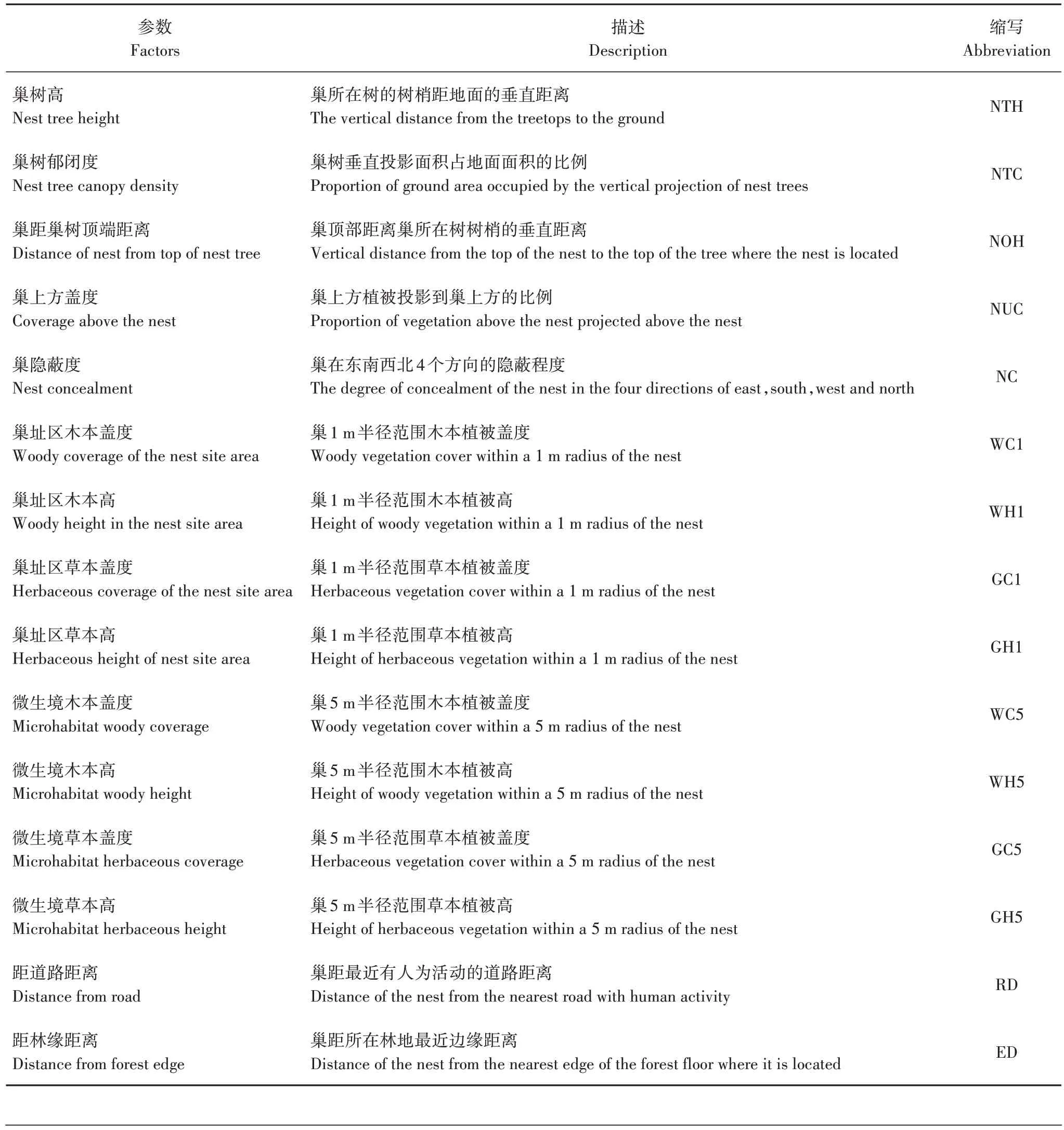
表1 褐柳莺巢址参数Tab.1 Parameters of dusky warbler nest site
2.3 数据分析
使用SPSS 26.0对所有参数进行主成分分析,研究褐柳莺巢址选择的主导因素;对于巢址因素的连续变量,用Kolmogorov-Smirnov 检验数据的正态性,符合正态分布的数据采用独立样本t检验,非正态分布的数据采用非参数检验中2 个独立样本的Mann-WhitneyU检验,分析繁殖成功巢和被捕食巢的巢址因素的差异,所有数据采用mean±SE 表示。P<0.05差异显著,P<0.01差异极显著。
3 结果
共记录30 个褐柳莺巢,巢树主要为云杉、樟子松,云杉上有18巢,占总数的60.00%;樟子松11巢,占总数的36.67%;黄花蒿(Artemisia annua)1 巢,占总数的3.33%。对所有巢的15 个巢址参数的主成分分析表明:前5 个主成分的特征值大于1,累计贡献率达到了87.582%,能够解释15 个巢址参数的全部信息,提取前5个因子作为主成分(表2)。
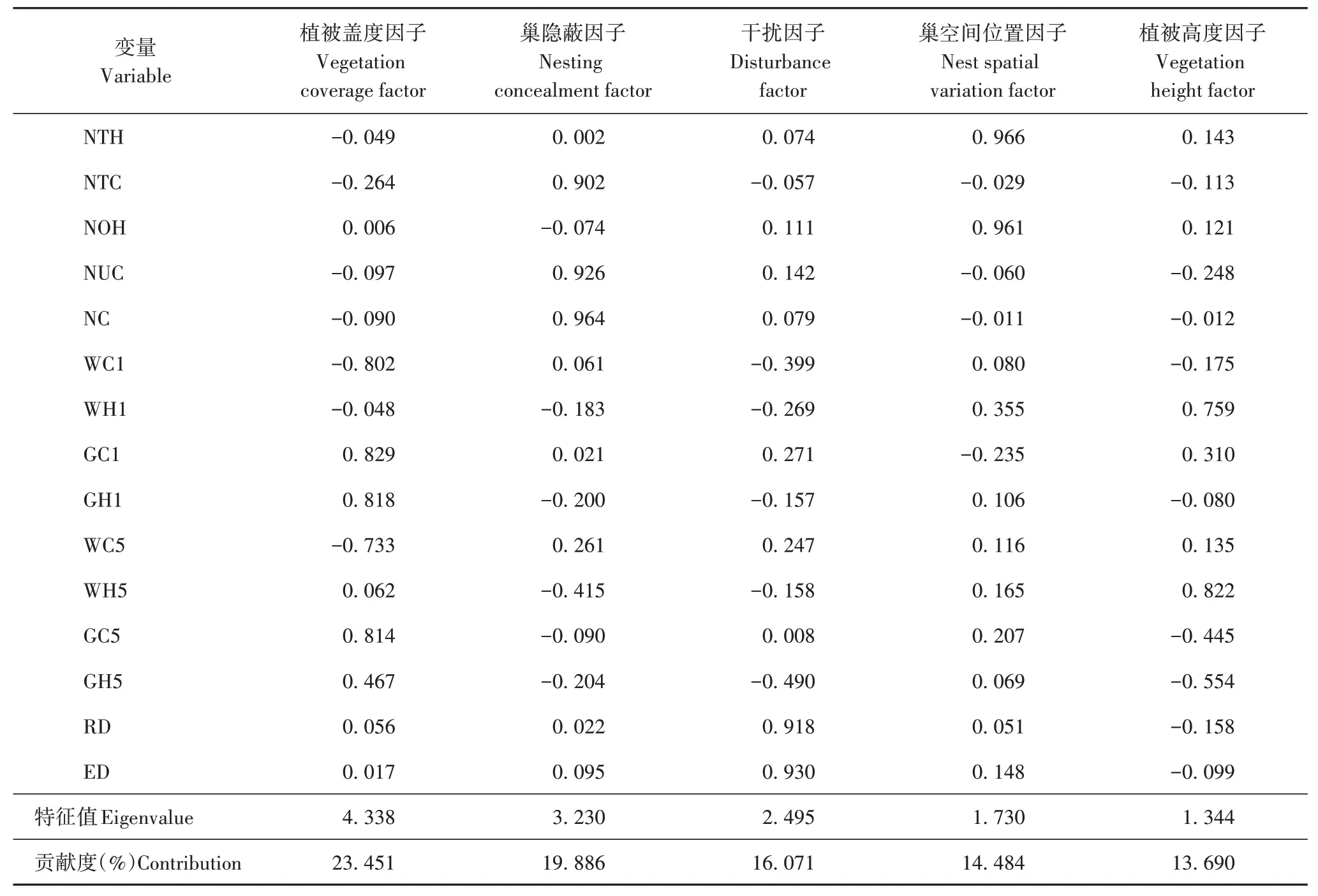
表2 褐柳莺巢址选择旋转后因子成分矩阵Tab.2 Rotated factor component matrix for dusky warbler nest site selection
主成分1 的特征值为4.338,贡献率最高,为23.451%,其中木本植被盖度、草本植被盖度载荷绝对值明显偏高,体现了巢址区域和5 m 微生境区域的植被盖度,因此将其命名为植被盖度因子;主成分2 的特征值为3.230,贡献率为19.886%,累计贡献率为43.337%,其中载荷较高的因子是巢树郁闭度、巢上方盖度和巢隐蔽度,三者都反映了巢的隐蔽性,因此命名为巢隐蔽因子;主成分3 的特征值为2.495,贡献率为16.071%,累计贡献率为59.408%,其中距道路距离、距林缘距离都反映了巢受人为干扰的程度,将其命名为干扰因子;主成分4 的特征值为1.730,贡献率为14.484%,累计贡献率为73.892%,其中巢所在植被高度、巢距巢树顶端距离载荷明显高于其他因子,都能反映巢的空间位置,因此命名为巢空间位置因子;主成分5 的特征值为1.344,贡献率为13.690%,累计贡献率为87.582%,其中巢址区木本植被高度、微生境木本植被高度载荷明显高于其他因子,二者都体现了木本植物的高度,因此命名为植被高度因子。
在已知命运的18个巢中,产第1枚卵的时间为5月28 日,最后1 只雏鸟离巢的时间为8 月12 日。66.67%(12/18)的巢至少有1 只雏鸟孵出,孵化率为70.77%(46/65)。46只雏鸟的离巢率为36.96%(17/46),繁殖成功率为27.78%(5/18)(至少有1 只雏鸟离巢视作繁殖成功)。在13个失败巢中,2巢为筑巢期和产卵期不明原因弃巢,1 巢孵化18 d 后失败,1 巢雏鸟饥饿而死,9 巢被捕食。在被捕食的9 巢中,根据视频监控及现场痕迹调查,有6 巢捕食者为赤狐(Vulpes vulpes),1 巢捕食者为牛头伯劳(Lanius bucephalus),其他巢捕食对象未被记录。
繁殖成功巢和被捕食巢在1 m 范围的巢址区植被盖度有显著差异,其中繁殖成功巢1 m 半径的巢址区木本植被盖度显著大于被捕食巢(Z=-2.525,P=0.012),而草本植被盖度显著小于被捕食失败巢(t=-2.847,P=0.014)(表3)。
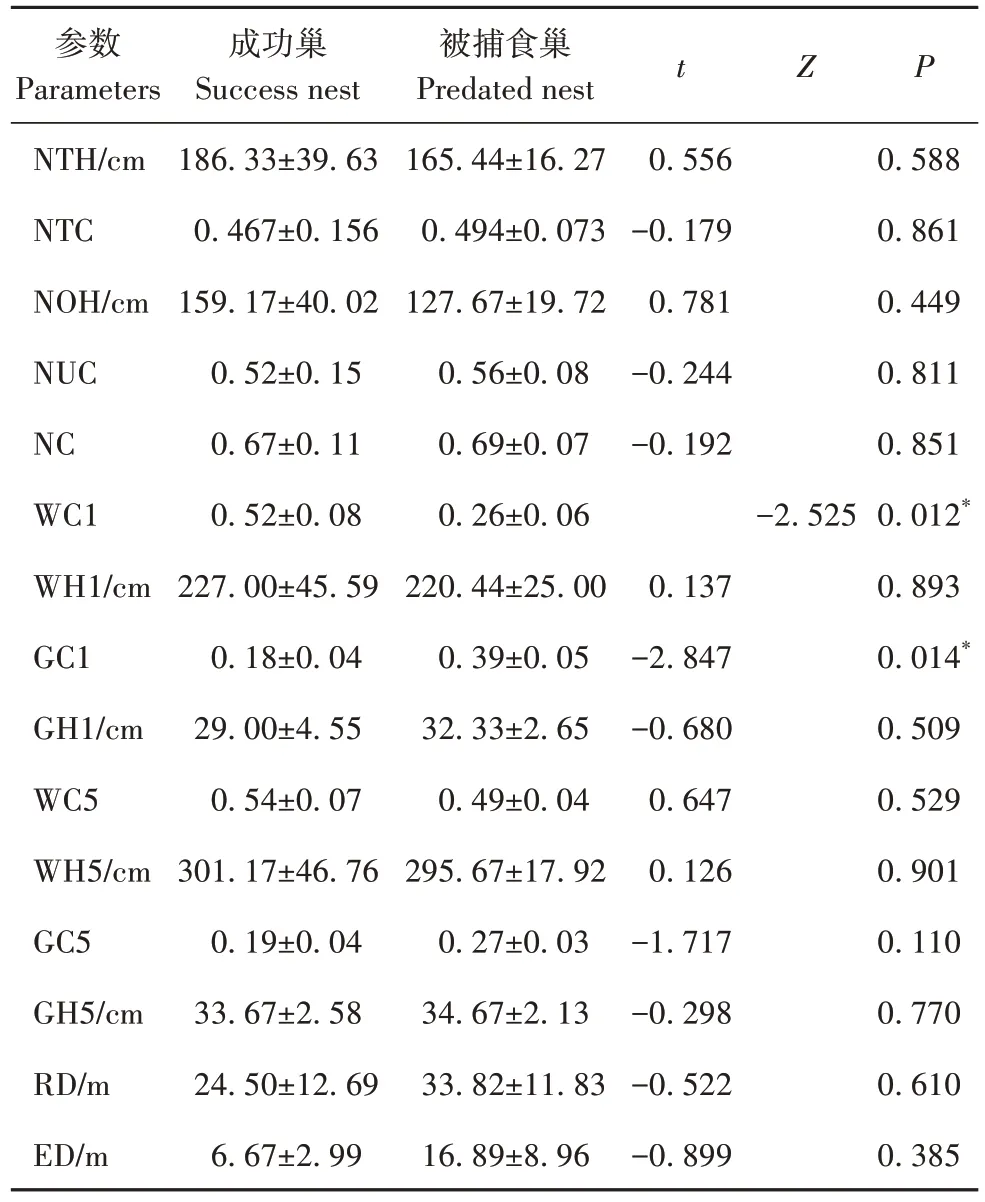
表3 褐柳莺繁殖成功巢与被捕食巢巢址参数的差异Tab.3 Differences in nest site parameters between breeding success nests and predated nests of the dusky warbler
4 讨论与结论
本研究所有褐柳莺的巢均位于人工苗圃地中的针叶林缘生境中,与赤狐的典型觅食地重合,赤狐引起的巢捕食是影响褐柳莺繁殖成效的主要因素,并且巢址附近的植被盖度是影响鸟类被捕食的主要因素。
褐柳莺主要营巢于低矮灌丛生境中,也有少量直接营巢于灌丛下的地面[25],本研究中褐柳莺均营巢于人工苗圃的幼苗或地面。事实上,本研究地区也分布许多北沙柳组成的天然灌木丛,繁殖期很多雄性褐柳莺频繁在其中鸣唱,但系统搜寻后,并未找到褐柳莺巢或找巢力度不够。这可能与本地的生境和植被结构有关,北沙柳构成的天然灌木林缺少主干,无法提供巢的放置点,5月底才开始萌芽,下端枝叶稀疏,隐蔽性差;而苗圃地的针叶林幼苗分叉较多,矮小茂密,与灌丛生境相似,可为褐柳莺筑巢提供更合适的营巢地点。据对巢址参数的分析,褐柳莺的巢址选择主要受植被盖度、巢隐蔽度、人为干扰、巢空间位置和植被高度5 个因子的影响,其中植被盖度和巢隐蔽因子显著影响巢周围的隐蔽度,而对人工苗圃中的针叶林来说,繁殖巢所在的位置与木本的高度决定着繁殖巢的隐蔽度,或许证明了人工苗圃生境独特的隐蔽性和安全性是影响本地褐柳莺巢址选择的重要因素。
对俄罗斯远东地区褐柳莺种群的研究发现,花鼠是影响其繁殖成效的重要因素,褐柳莺能够评估繁殖区花鼠的捕食风险,并根据风险高度调整巢址选择的策略[25,32]。依本研究,花鼠主要活动于成年针叶林或灌丛生境中[33],在苗圃地中未发现其活动痕迹,这可能是幼苗地中缺乏松塔等花鼠的食物;而褐柳莺选择在此种生境中筑巢,可能是对外界环境捕食风险评估后的自我调整策略。
本地区褐柳莺繁殖依然遭受50% 的捕食(9/18),捕食者主要为赤狐和牛头伯劳,与俄罗斯远东种群不同,其中赤狐捕食的巢占所有被捕食巢的66.67%。徐兴军等[34]对齐齐哈尔牧林交错区的草甸、灌丛、人工疏林和人工密林等不同生境类型的啮齿类(Rodentia)物种多样性调查发现,在人工疏林,啮齿动物的捕获率最高,灌丛和草甸较低,人工密林最低。本研究区植被类型和结构与其相似,因此在研究区人工种植的苗圃疏林中可能也拥有较多的啮齿类动物。而食物的丰富度可能是影响赤狐生境选择的主要因素[35-36],赤狐的主要食物为啮齿动物、野兔、腐肉、鸟类和水果,小型啮齿动物为其喜好食物[37],因此,人工种植的苗圃地可能是赤狐觅食的主要场所,拥有较高的活动强度。对繁殖成功巢和被捕食巢的巢址参数比较结果表明,1 m半径范围巢址区域的植被盖度是影响褐柳莺巢是否被捕食的关键因素,被捕食的巢在1 m 半径范围内的地面草本覆盖度较高,拥有较高的隐蔽度,但赤狐的听觉和嗅觉非常灵敏,在搜索小型啮齿动物时,同时搜索到筑于幼苗下半部的褐柳莺繁殖巢。此外,赤狐主要在夜间探测和捕食鸟类巢穴[38-40],因此,苗圃地中草本形成的地上隐蔽度不能有效降低赤狐的捕食风险;而苗圃中云杉和樟子松幼苗枝叶的硬度较大,较高的木本盖度可以有效阻挡捕食者的接近,从而提高繁殖成功的概率。这也可能是褐柳莺选择在人工苗圃生境中营巢的重要因素,但仍需要通过不同生境的捕食实验进一步验证。
综上,本研究提供了中国塞罕坝地区褐柳莺的巢址选择数据,扩充了褐柳莺在人工林中繁殖的相关资料。本地区褐柳莺偏爱筑巢于植被盖度高,隐蔽性好,远离人为干扰的苗圃地植被中下部;而巢址较高的木本植被盖度增强了捕食者接近的难度,降低了被捕食的风险。
