唐家河国家级自然保护区川金丝猴的种间关联和保护
2023-02-22王晓娟谌利民李兆元刘卓涛
王晓娟 ,谌利民 ,钟 义 ,李兆元,刘卓涛,李 谦
(1.西南林业大学地理与生态旅游学院,昆明,650224;2.唐家河国家级自然保护区管理处,青川,628109;3.中国野生动物保护协会,北京,100714)
川金丝猴(Rhinopithecus roxellana)属于喜马拉雅—横断山区物种[1],曾因栖息地丧失、破碎化及偷猎等导致其分布范围缩小、种群数量减少,现分布于陕西秦岭局部、湖北神农架、甘肃南部以及四川岷山、邛崃山和凉山山系[2]。在群落中,川金丝猴通过体内和体外2 种方式传播植物种子,有效促进植物的更新以及森林生态系统的稳定[3-4],发挥着重要生态学作用。
群落是物种通过种间生态学关系(如捕食、竞争和共生等)构成的网络[5],物种间的互动是群落生态学研究的核心,是物种演化和物种多样性保护研究的关键。川金丝猴的群落生态学研究已有初步探讨,除上述川金丝猴与植物的生态学关系研究[3-4]外,还有川金丝猴与其他野生动物间的生态学关系研究,如李爽等[6]认为川金丝猴与群落中的大熊猫(Ailuropoda melanoleuca)、扭角羚(Budorcas taxicolor)对环境因子的不同需求导致三者在生境利用上既有相似性又有差异性。在四川唐家河,藏酋猴(Macaca thibetana)和川金丝猴在很大程度上通过生境分离以及不同的生境利用和空间避让而共存[7]。类似研究也出现在灵长类其他物种中,如与川金丝猴类似的食果灵长类也是植物种子的重要传播者,对森林植被的更新以及群落的稳定具有积极作用[8-9]。生活在同一群落中的普通狨(Callithrix jacchus)使用声音信号作为线索,避免与金发悬猴(Sapajus flavius)相遇[10]。在婆罗洲丹南河谷,习性相似导致群落内5 种灵长类之间相互回避,从而失去种间空间关联[11]。灵长类与群落中其他非灵长类动物间的关系也有报道,如亚马逊森林的黑帽悬猴(Cebus apella)与白南美鵟(Leucopternis albicollis)[12]、巴西伊比亚帕巴高原(Ibiapaba Plateau,Brazil)的髯悬猴(Sapajus libidinosus)与岩豚鼠(Kerodon rupestris)[13]。目前,灵长类群落生态学的研究大多关注灵长类在种子传播对植被更新中的作用及其与其他类群间的互动,其研究思路植根于早期对昆虫与植物的协同演化研究[14]。
川金丝猴生态学研究还涉及家域[15-16]、活动节律和活动时间分配[17-18]、食性[19-20]以及生境[7,16],这些报道中均强调对川金丝猴保护的必要性。单种保护与生物多样性整体保护之间的矛盾已经被长期关注。当大熊猫从人类的保护中获益时,豺(Cuon alpinus)、豹(Panthera pardus)、雪豹(P.uncia)、狼(Canis lupus)[21]、林麝(Moschus berezovskii)和亚洲黑熊(Ursus thibetanus)[22]等物种却受损。这种矛盾的根源在于单种保护忽略了种间互动及其影响的传递作用,为此,物种互动网络的保护成为解决这种矛盾的钥匙[23]。一个特定物种与其他物种互动的网络构成该物种的群落环境,该物种在这个环境中生存和演化[24],然而,通过行为学观察结果建立物种互动网络长期以来存在资金、人力和现实约束等障碍[25]。物种互动以共同出现于相同地理空间为前提,Forbes[26]最早开始通过物种的空间关联来研究种间互动,在过去100 年中,这种尝试一直局限于种对的关联,从未提出关联网络,同时,在方法上也存在一系列问题[27-28]。2018 年以来,本课题组探索使用空间分析方法构建物种关联网络来探讨种间互动。杨虎等[29]认为,虎(Panthera tigris)的缺失导致卧龙扭角羚在生物群落中失去了种群控制因子;周厚熊等[30]提出,雪豹和赤狐(Vulpes vulpes)在群落中协调控制卧龙食草动物的种群数量;李谦等[31]认为,水鹿(Rusa unicolor)活动能够为卧龙的野猪(Sus scrofa)、中华鬣羚(Capricornis milneedwardsii)和毛冠鹿(Elaphodus cephalophus)创造适宜生境,因此有利于这些物种的生存。
据刘卓涛等[32]研究,卧龙川金丝猴是群落中的核心物种之一,其种群的发展可能会促进大熊猫、黄喉貂(Martes flavigula)种群的发展,也可能导致水鹿、毛冠鹿和马来豪猪甚至更多其他物种的衰落。同一物种进入不同区域的生物群落时,由于区系不同,群落环境也会发生变化。本研究基于唐家河国家级自然保护区川金丝猴的物种空间关联网络,探讨川金丝猴的群落环境,分析川金丝猴与其他物种的生态学关系及对其保护的传递影响,进而从群落生态学和保护生物地理学的新视角探讨川金丝猴的保护问题。
1 研究区概况
唐家河国家级自然保护区位于四川省广元市青川县境内(32°30'—32°41' N,104°36'—104°52' E),面积约400 km2,海拔1 100~3 864 m,地势自西北向东南倾斜。年平均气温12 ℃,年平均降水量1 022 mm,降雪最早在11月,终止于次年4月。区内水系发达,北路沟与唐家河构成主要支流。保护区内2 764 m的海拔差孕育出复杂的植被类型:从低到高分别为常绿阔叶林(海拔<1 500 m)、常绿与落叶阔叶混交林(1 500~2 000 m)、针阔叶混交林(2 000~2 500 m)、针叶林(2 500~3 400 m)、高寒草甸(3 400~3 600 m)以及高山灌丛(>3 600 m)。丰富的植被类型促成生境的多样性,孕育出丰富的动物种类。区内兽类共计103 种,包括国家一级重点保护野生动物大熊猫、川金丝猴、豹、云豹(Neofelis nebulosa)、林麝和马麝(Moschus chrysogaster)等,二级重点保护野生动物猕猴(Macaca mulatta)、藏酋猴、亚洲黑熊、喜马拉雅小熊猫(Ailurus fulgens)、黄喉貂、中华鬣羚和中华斑羚(Naemorhedus griseus)[33]。
2 研究方法
2.1 野外数据采集
2019 年10 月—2021 年12 月,在唐家河国家级自然保护区利用红外相机采集野外数据。利用Arc-GIS 软件将保护区划分为1 km×1 km 的网格,在可通达网格中的典型生境安装红外相机(猎科Ltl Acorn-6210),固定在距地面50~80 cm 的树干或其他固定物体上,保持镜头与地面平行,相机位点间距≥300 m。相机设置为连续拍摄3 张照片后录像10 s,24 h 制。记录相机位点的安装时间、经纬度、海拔以及内存卡编号等相关信息,同时巡视相机附近100 m 以内的区域,记录发现的动物实体及痕迹,以补充该相机位点的信息。每季度更换1 次电池和储存卡,重复上述操作。基于红外相机的工作原理和上述安装操作,所拍摄到的物种为地栖鸟类和兽类。
获得野外资料后,为每台相机建立数据集,逐一查看储存卡中的拍摄内容,参照《中国兽类野外手册》[34]和《中国鸟类野外手册》[35]对视频与图片中的物种进行鉴定。去除因故障导致无效数据的相机位点,按照每个公里网格选取1台且拍摄时间达到1年的标准,最终获得103 个相机位点数据(图1)。由于所获照片难以确保个体识别的准确性,仅提取每台相机中各物种出现与否的数据,即二分型数据。
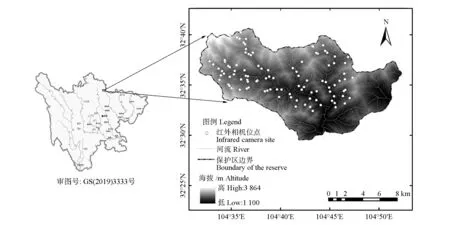
图1 唐家河国家级自然保护区红外相机位点分布Fig.1 The distribution of infrared cameras in the Tangjiahe National Nature Reserve
2.2 种间关联分析
2.2.1 种间关联度衡量及物种网络构建
采用佛爱系数衡量种间空间关联度[31,36],本研究中N=103。基于计算结果,舍弃不具备显著性(P>0.05)的佛爱系数及其物种种对,视为偶然因素导致的种间关联,保留具有显著性(P≤0.05)的种对,运用NetDraw 软件,以这些物种为节点,构建川金丝猴空间关联网络。在网络中,川金丝猴的关联物种团由与川金丝猴存在直接空间正关联的物种构成,它们是川金丝猴的群落环境。
2.2.2 空间关联属性分析
在地理要素的空间分布中,由于内在关系影响,一个要素的出现会诱发其他要素出现。在统计学上,当要素A出现时,判断要素B出现的出错率相应地下降[36],这时,要素的空间关联属性为不对称性,要素A对要素B存在预测性。要素间的预测性有时是单向的,即要素A预测要素B;有时是双向的,即A预测B的同时,B也预测A。当2 个要素不存在预测性时,则表明这2 个要素纯粹是因为其存在所需条件相同而出现在相同地理空间中,关联属性为对称性。本研究中,2 个物种被视为2 个地理要素,内在关系是它们之间的生态学关系。为了检验种间关联的属性,考虑到数据为二分型,采用兰布达(Lambda)系数检验,计算过程参阅《实用统计学方法》[36]和刘卓涛等[32]。
2.2.3 物种的空间关联多样性及其对群落维持的贡献率
采用香农-维纳指数(Shannon-Wiener index)来衡量空间关联多样性,计算公式如下:

式中:当计算某一特定物种的香农-维纳指数时,S为与该物种直接正关联的种对总数,Pi为种对i的关联系数占该物种关联系数总和的比例。H'越大,则该物种空间关联多样性越高。
计算出各物种的H'后,求取平均值。H'大于 平均值的物种被定义为网络核心物种,对网络维 持有重要作用;H'小于平均值的物种为外周物种,对网络维持的作用相对较小。物种对网络维持的贡 献率用于直观展示各物种对网络维持的重要性,计算公式如下:

式中:S为物种数;H'i为物种i的香农-维纳指数;Ci越大,物种i对网络维持的贡献率越高,对网络的重要性越大。
3 结果
3.1 唐家河川金丝猴地栖鸟兽区系环境
红外相机共获得20 093张清晰有效照片和4 062个视频,从中鉴定出6 目14 科29 种地栖鸟兽,包括1类未鉴定(影像模糊,无法准确鉴定)的鼠科(Muridae)动物(表1)。

表1 唐家河国家级自然保护区地栖鸟兽种类构成Tab.1 Species composition of terrestrial bird and mammal in the Tangjiahe National Nature Reserve
表1 显示,兽类共5 目13 科(包括1 类未鉴定的鼠科动物)21 属23 种,鸟类1 目1 科6 属6 种。兽类中食肉目(Carnivora)物种最多,共4科8属9种;其次为鲸偶蹄目(Cetartiodactyla),4 科7 属7 种;啮齿目(Rodentia)2 科3 属3 种,外加1 类未鉴定的鼠科动物;灵长目(Primates)1科2属3种,包括川金丝猴、猕猴和藏酋猴;兔形目(Lagomorpha)最少,仅1科1属1种,即间颅鼠兔(Ochotona cansus)。
3.2 川金丝猴及其空间关联物种
在103 个相机位点中,川金丝猴在57 个位点中出现,分布海拔为1 265~3 041 m。基于佛爱系数计算结果,唐家河国家级自然保护区地栖鸟兽存在2个网络:一个是由18 个物种及鼠科动物组成的低海拔网络(low altitude network,LAN);另一个是由11个物种组成的高海拔网络(high altitude network,HAN)。川金丝猴属于唐家河LAN物种(图2)。
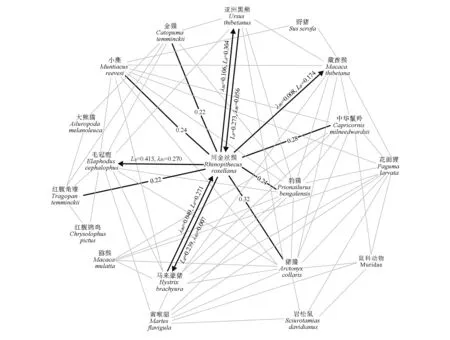
图2 唐家河LAN结构及川金丝猴关联物种团Fig.2 Structure of Tangjiahe LAN and the species guild of Sichuan snub-nosed monkey
由图2 见,川金丝猴与10 个物种存在直接空间正关联,包括毛冠鹿(rø=0.49,χ2=22.14,P<0.05;分布海拔为1 265~3 041 m)、亚洲黑熊(rø=0.37,χ2=12.57,P<0.05;分布海拔为1 300~2 759 m)、马来豪猪(rø=0.33,χ2=9.94,P<0.05;分布海拔为1 220~ 2 759 m)、猪獾(rø=0.32,χ2=9.59,P<0.05;分布海拔为1 264~2 759 m)、中华鬣羚(rø=0.28,χ2=6.62,P<0.05;分布海拔为1 264~2 759 m)、藏酋猴(rø=0.24,χ2=4.97,P<0.05;分布海拔为1 220~2 759 m)、豹猫(rø=0.24,χ2=4.97,P<0.05;分布海拔为1 220~ 3 041 m)、小麂(rø=0.24,χ2=4.95,P<0.05;分布海拔为1 220~2 759 m)、金猫(rø=0.22,χ2=4.31,P<0.05;分布海拔为1 265~3 373 m)和红腹角雉(rø=0.22,χ2=4.22,P<0.05;分布海拔为1 541~3 041 m)。它们与川金丝猴构成了关联物种团,物种团约占唐家河LAN物种总种数的58%。猕猴没有出现在该物种团中。
3.3 种对关联属性
对川金丝猴关联物种团成员进行的兰布达系数计算结果显示:
(1)双向非对称关联存在于川金丝猴和马来豪猪以及川金丝猴和亚洲黑熊之间(图2),其中川金丝猴对亚洲黑熊的预测率为5.6%~27.3%(LB=0.273,λB0=0.056,P≤0.05),亚洲黑熊对川金丝猴的预测率为10.6%~30.4%(LB=0.304,λB0=0.106,P≤0.05),表明亚洲黑熊对川金丝猴的空间分布吸引力更大。川金丝猴对马来豪猪的预测率为4.9%~27.1%(LB=0.271,λB0=0.049,P≤0.05),马来豪猪对川金丝猴的预测率为0.7%~23.9%(LB=0.239,λB0=0.007,P≤0.05),表明川金丝猴对马来豪猪的空间分布吸引力更大。
(2)单向非对称关联出现在毛冠鹿、藏酋猴与川金丝猴之间。毛冠鹿对川金丝猴的预测率为27.0%~41.3%(LB=0.413,λB0=0.270,P≤0.05),藏酋猴对川金丝猴的预测率为0.8%~17.4%(LB=0.174,λB0=0.008,P≤0.05)。川金丝猴在空间分布上以不同概率被这2 个物种吸引,其中毛冠鹿的吸引力远大于藏酋猴。
(3)川金丝猴与猪獾、中华鬣羚、豹猫、小麂、金猫和红腹角雉之间存在对称空间关联。
3.4 关联物种对网络维持的贡献
在唐家河LAN 中,物种香农-维纳指数的平均值为1.79,其中H'>1.79 的物种有10 个,分别为猪獾、豹猫、花面狸、马来豪猪、毛冠鹿、川金丝猴、中华鬣羚、藏酋猴、小麂和黄喉貂,属于网络核心物种;H'<1.79的物种有9个,分别为亚洲黑熊、红腹角雉、金猫、鼠科动物、猕猴、岩松鼠、野猪、大熊猫和红腹锦鸡,属于网络外周物种(表2)。川金丝猴为网络核心物种,香农-维纳指数及贡献率位居第6 位。川金丝猴物种团中,属于核心物种的还有猪獾、豹猫、马来豪猪、毛冠鹿、中华鬣羚、藏酋猴和小麂,占核心物种的80%,其余属于外周物种,包括亚洲黑熊、红腹角雉和金猫。
4 讨论
4.1 川金丝猴物种团的生态结构
本研究所获数据是种对在特定空间共同出现的频率,计算结果是有生态学意义的空间共现概率。依据空间关联原理,对称关联反映2 个物种在相同空间中对共同生态资源(如食物资源、活动场所)的需求,因此是竞争性关系。单向非对称关联反映一个物种倾向于分布在另一个物种出现的空间中,这种趋向性源于前一物种从后一物种的存在中获益,属于偏利性关系。双向非对称关联反映2 个物种倾向于分布在对方出现的空间中,相互从对方的存在中获益,属于互利性关系。依据这些规则,引用已发表的生态学文献,将空间关联进行解析。
4.1.1 互利生态学关系
川金丝猴—亚洲黑熊:本研究结果显示,川金丝猴与亚洲黑熊在空间分布上相互吸引。亚洲黑熊选择郁闭度适中(30%~50%)、阳坡、乔木和灌木发育较好、林下障碍物较少的山坡面和山脊的阔叶林和针阔混交林生境[37],这类生境也是川金丝猴的主要活动区域[16]。2 种动物的共同食物资源有猕猴桃属(Actinidia)、花楸属(Sorbus)、胡桃楸(Juglans mandshurica)、榛(Corylus heterophylla)、板栗(Castanea mollissima)、华中五味子(Schisandra sphenanthera)、木姜子(Litsea pungens)和灯台树(Cornus controversa)等[19,33,38]。相同的活动区域和觅食导致川金丝猴和亚洲黑熊产生空间关联。川金丝猴能传播蔷薇科(Rosaceae)、伞形科(Apiaceae)、猕猴桃属和紫菀属(Aster)等植物种子[3-4],促进植物幼苗生长;亚洲黑熊则以这些植物的嫩茎、叶和果为食[38]。冬春季节,受食物资源和气候条件影响,川金丝猴活动于落叶阔叶林和针阔混交林的阳坡生境,取食嫩枝、叶和树皮,常导致树冠畸形、小枝突发或全株死亡[39],改变林冠郁闭度,增加光照,促进下层灌木、草本生长[40],为亚洲黑熊创造适宜生境[39]。亚洲黑熊还可能掠食川金丝猴尸体[41]。可见,亚洲黑熊受益于川金丝猴,从而在空间分布上倾向于(5.6%~27.3%的可能性)趋近川金丝猴活动区。每年4—7月,亚洲黑熊大量觅食各种草本植物[42],导致草窗(herb gap)生境出现,促进乔木幼苗生长[43],并为川金丝猴所利用,导致川金丝猴的空间分布倾向于(10.6%~30.4%的可能性)趋近亚洲黑熊活动区。在这对互利关系中,亚洲黑熊为川金丝猴提供的利益大于川金丝猴对亚洲黑熊提供的利益。
川金丝猴—马来豪猪:马来豪猪选择山地草坡、灌丛或密林生境,洞穴多隐蔽于草丛中[44];而川金丝猴季节性迁移在阔叶林、针阔混交林以及针叶林间,活动于乔木层中[39]。这些乔木下主要是灌木层(竹林灌丛)和草本层,盖度适中,隐蔽性较好[33],是马来豪猪偏好的生境。冬春季节,2种动物均取食楤木属(Aralia)、杜鹃花属(Rhododendron)和菝葜属(Smilax)植物,进入相同地理空间[33,39,44]。川金丝猴属于群居物种,夜宿于高大乔木的树叉和树枝上[39],树下常出现大量猴粪[2],促进林下草本植物和土壤无脊椎动物生长,为马来豪猪提供丰富食物资源[44]。这种利益可能导致马来豪猪的空间分布倾向于(4.9%~27.1%的可能性)趋近川金丝猴活动区。同时,马来豪猪以竹丛中的树洞作为栖息地,春季取食竹笋[44],咬断竹子以便于通行[45]。竹林灌丛的存在阻碍乔木幼苗生长[46],将竹林疏化有利于乔木生长,促使川金丝猴空间分布倾向于(0.7%~23.9%的可能性)趋近马来豪猪活动区。这对互利关系中,川金丝猴为马来豪猪提供的利益大于马来豪猪为川金丝猴提供的利益。与川金丝猴—亚洲黑熊种对相比,该种对的种间互利性较小。
4.1.2 偏利生态学关系
川金丝猴—毛冠鹿:春季,毛冠鹿偏好距离食物和水源地近、隐蔽条件好、远离干扰的缓坡、阳坡阔叶林生境[47];夏季,活动在海拔较高、远离人为干扰的针阔混交林[48]。这些生境也是川金丝猴在春夏季节所选择的活动生境[7]。冬季,毛冠鹿常出现在灌木密度和草本覆盖大、竹子密度低和距离道路较近的空旷生境[48-49],与川金丝猴产生一定的空间分离[16]。2个物种均以松属(Pinus)、栎属(Quercus)、杜鹃花属、菝葜属和中华猕猴桃(Actinidia chinensis)等植物的叶、果及种子为食[20,33,50]。本研究结果显示,毛冠鹿在空间分布上吸引川金丝猴。冬季和初春,毛冠鹿活动于草本盖度大的生境,取食草本植物[47,49],改变草本层结构;春季偏好在隐蔽条件较好的竹林灌丛中取食竹叶和竹笋[47,51]而疏化竹林,为川金丝猴赖以生存的乔木幼苗生长创造条件[43,46]。这可能导致川金丝猴倾向于(27.0%~41.3%的可能性)分布在毛冠鹿的活动区。
川金丝猴—藏酋猴:在唐家河国家级自然保护区,藏酋猴全年活动于1 500~2 000 m 的常绿落叶阔叶混交林和落叶阔叶林中[52],而川金丝猴具有明显的季节性垂直迁移习性,仅在秋冬季节活动于落叶阔叶林以及针阔混交林中[7]。冬季,川金丝猴与藏酋猴均偏好落叶阔叶林中坡度较小的阳坡、靠近水源的生境[7]。2个物种有广泛的共同食源种,如中华青荚叶(Helwingia chinensis)、青荚叶(H.japonica)、灯台树、猫儿屎(Decaisnea insignis)、三叶木通(Akebia trifoliata)、山胡椒(Linderaglauca)、木姜子、桦叶荚蒾(Viburnum betulifolium)、中华猕猴桃、珙桐(Davidia involucrate)等,因而在相同地理空间中觅食[19-20,33,53-54]。在食物匮乏的冬季,2 个物种存在食物竞争现象[7]。藏酋猴的食源种大多属于草本层和灌木层成分,秋季偏好竹笋[53,55],摄食活动降低林下竹灌丛密度,促进川金丝猴偏好的灌木密度适中的乔木生境的形成[16]。在取食过程中,藏酋猴为山胡椒属(Lindera)、四照花属(Dendrobenthamia)、猫儿屎以及珙桐等植物传播种子[53],这些植物的叶和果也是川金丝猴的食物[20]。此外,藏酋猴有相对固定的家域[56],熟悉家域内食物资源分布,跟随藏酋猴有利于川金丝猴发现食物资源。这些利益可能导致川金丝猴倾向于(0.8%~17.4%的可能性)在藏酋猴活动区中出现。
4.1.3 竞争生态学关系
受食物资源和气候条件的影响,川金丝猴在唐家河国家级自然保护区内的落叶阔叶林、针阔混交林以及针叶林生境之间进行季节性垂直迁移。猪獾倾向于巢居在灌木盖度高、隐蔽性好、距水源较近和阳坡或半阴半阳坡的生境中[57]。在唐家河国家级自然保护区,猪獾见于海拔1 264~2 759 m 的阔叶林和针阔混交林中,这里属于川金丝猴春、秋和冬季的活动区域。此外,猪獾在秋季取食栎属、松属植物的种子[58],与川金丝猴存在部分食性重叠[20]。
中华鬣羚因对食物和隐蔽条件的需求也在上述植被间作季节性垂直迁移,但在春季偏好选择陡坡生境,冬季远离乔木、灌木生境而与川金丝猴存在的生境分离[16,59]。2种动物共同取食木姜子、三桠乌药(Linderaobtusiloba)、毛樱桃(Prunustomentosa)、峨眉蔷薇(Rosaomeiensis)、淡红忍冬(Lonicera acuminata)、荚蒾(Viburnum dilatatum)和接骨木(Sambucuswilliamsii)等[19-20,33,60],并因此进入相同地理空间竞争食物资源。
豹猫在川金丝猴活动的林型中以白腹鼠属(Niviventer)、姬鼠属(Apodemus)和鼠兔属(Ochotona)动物为主要捕食对象。豹猫和川金丝猴共同的食物物种包括蔷薇科、栗属(Castanea)和栎属植物[20,61-62]。
小麂活动于川金丝猴冬季常选择的低海拔平地或缓坡、隐蔽条件好的多灌丛生境[63-64],与川金丝猴共同取食蔷薇科、忍冬科、菝葜属、栎属和杜鹃花属等植物的叶、芽或果[19,65]。
金猫选择以冷杉(Abies fabri)为主的针叶林、针阔混交林、灌丛以及草甸等生境[66],与川金丝猴存在部分生境重叠。本研究结果显示,金猫与川金丝猴不存在非对称关系,表明这2个物种间不存在捕食关系,但胡锦矗等[39]的研究发现,金猫会在夜间爬树捕食川金丝猴,二者是否存在捕食关系有待进一步研究。
红腹角雉在冬季常活动于植被盖度较好、靠近水源的阳坡竹灌丛生境中[67],夏秋季主要取食灌木和乔木的熟果[68]。红腹角雉和川金丝猴均偏好人为干扰少、隐蔽条件较好的生境[16,69]。
以上讨论表明,川金丝猴与猪獾、中华鬣羚、豹猫、小麂、金猫和红腹角雉因共同食物物种或共同生境需求而产生部分空间关联。在这种生态学关系中,它们存在一定程度的资源竞争。川金丝猴在卧龙与毛冠鹿、马来豪猪和水鹿有类似的生态学关系[31-32]。
4.2 川金丝猴群落环境的比较
川金丝猴的群落环境在卧龙[32]和唐家河国家级自然保护区之间的共同点是均无捕食威胁,均有互利、偏利和竞争的生态学关系,但物种组成不同。卧龙互利物种是大熊猫和黄喉貂,唐家河国家级自然保护区是马来豪猪和亚洲黑熊;卧龙偏利物种是林麝和喜马拉雅小熊猫,唐家河国家级自然保护区是藏酋猴和毛冠鹿;卧龙竞争物种是水鹿、毛冠鹿和马来豪猪,唐家河国家级自然保护区是中华鬣羚、小麂、猪獾、豹猫、金猫和红腹角雉。
在卧龙,川金丝猴物种团有8 种物种(占卧龙LAN 物种的40%),且全部属于网络核心物种,但仅占核心物种的67%[32];而在唐家河国家级自然保护区有11 个(占唐家河LAN 物种的58%),有8 个属于网络核心物种,占核心物种的80%,物种团有27%的物种位列网络外周。川金丝猴在卧龙的香农-维纳指数(H')为1.920(唐家河国家级自然保护区H'为2.269),贡献率位居核心物种第7 位(唐家河国家级自然保护区位居第6位)[32]。
种间关联多样性及对网络影响力的差异可能与川金丝猴在两地适宜生境面积的不同有关。在卧龙,川金丝猴的适宜生境面积为72.45 km2[70];在唐家河国家级自然保护区,各季节的适宜生境面积均大于140 km2[7]。较大的适宜生境允许川金丝猴发展种间生态学关系并因此充分发挥其在群落中的生态学功能,这表明川金丝猴在唐家河LAN 中拥有更复杂的群落环境,对群落的影响更大。
4.3 网络动力学分析
在互利生态学关系中,物种倾向于相互跟随对方;在偏利生态学关系中,受益物种倾向于跟随施益物种。因此,偏利关系和互利关系为网络提供内聚力,对群落中物种的集结起促进作用。唐家河国家级自然保护区川金丝猴与亚洲黑熊和马来豪猪之间相互吸引,表明川金丝猴分别为亚洲黑熊和马来豪猪提供利益,并从这2 个物种中获益。生存利益促使3 个物种在空间上联结并维持生态学关系。毛冠鹿、藏酋猴吸引川金丝猴,促进川金丝猴的生存和发展,使川金丝猴在空间分布上趋近毛冠鹿和藏酋猴,进一步将毛冠鹿和藏酋猴联结到生态关系网络中。
在竞争关系中,种间存在相互排斥(高斯原理),为网络提供离心力,对群落中物种的集结起阻碍作用。猪獾、中华鬣羚、豹猫、小麂、金猫和红腹角雉与川金丝猴对称关联的生态学基础是对生境或食物资源利用的部分相似性,这种相似性导致一定程度的种间竞争。种间竞争形成种间排斥力,是群落的离心力。然而,内聚力能够通过种间关系延伸到网络中的其他物种,并且抵消这种离心力,如小麂与川金丝猴存在离心力,但小麂在空间分布上被藏酋猴吸引[27],并通过藏酋猴对川金丝猴的吸引关系抵消离心力,从而得以保留在网络中。2种力量的平衡取决于2 个物种种群的互动。另外,内聚力在种间的渐次延伸还将无空间关联的物种维系在一起,如野猪属于川金丝猴物种团外的物种,与川金丝猴没有空间关联(图2),但它通过吸引藏酋猴以及小麂实现与川金丝猴物种团间的内聚力[27],完成了12 个物种的内聚。内聚力还通过物种团其他成员实现延伸。可见,渐次联结不仅抵消群落离心力,还联结更多看似无关联的物种。群落的维持最终取决于离心力和内聚力的平衡。
4.4 关于川金丝猴保护的启示
网络动力学分析显示,物种间通过具体的生态学关系结成网络,并实现互相制衡的种间关系。离心力和内聚力的平衡是长期群落演化过程中不同种群间互动的结果,物种得以在这种环境下生存和演化。然而,生态学对种群间互动的了解非常有限[24]。灵长类及其他类群的种间互动研究大多基于种对分析,忽略了网络动力学分析。本研究仅包括1 个简化的地栖鸟兽群落结构(图2)。也仅能对网络中与川金丝猴关联的部分物种进行定性的动力学分析,但全面而细致的分析应是涉及全网络物种的复杂定量计算。定性分析已经表明,出于物种保护目的,人为促进一种动物的种群增长或栖息地面积增加可能会通过目前未知的途径危害其他物种,为生物多样性保护带来风险,正如Li 等[21]和Wang 等[22]所揭示的结果。为此,保护机构在针对单一物种保护制订行动计划时需要特别慎重。
面对庞大的生物多样性,保护资源永远是有限的,保护生物地理学的重要责任之一是为保护区域提出优先性排序[71]。以上对唐家河LAN 和卧龙LAN 的比较显示,川金丝猴在唐家河LAN 中拥有更多关联物种,群落环境更好,并且在群落维持中发挥更大作用。在保护资源短缺的情况下,对唐家河国家级自然保护区川金丝猴的投入能确保该物种生存的同时,惠及更多物种。因此,在对川金丝猴的保护中,应该优先投资唐家河国家级自然保护区。
致谢:感谢四川省林业和草原局古晓东以及唐家河国家级自然保护区肖梅、何芳、陈万里对野外工作的大力支持,感谢西南林业大学杨晓军老师、北京林业大学苏腾伟先生、昆明逸境生态工程咨询有限公司周厚熊先生对文稿提出的宝贵意见,感谢西南林业大学刘萍老师在该项目实施过程中提供的后勤保障。
