红壤有机碳组分中微生物群落构成及均匀度决定其矿化特征
2023-02-22单会茹段英华徐明岗
单会茹,张 璐,高 强,段英华*,徐明岗*
(1 吉林农业大学资源与环境学院,吉林 长春 130118;2 中国农业科学院农业资源与农业区划研究所 /农业农村部耕地质量监测与评价重点实验室,北京 100081)
土壤有机碳库是陆地生态系统中最大、最活跃的碳库[1],在全球碳循环体系中起到举足轻重的作用。土壤有机碳矿化是土壤有机碳向大气周转的一个重要的生物化学过程[2],该过程主要受土壤有机碳稳定性和土壤微生物群落等多种因素的影响[3]。土壤有机碳稳定性是指其抵抗外界干扰维持自身原有水平的能力[4]。为了更好的研究不同稳定机制碳组分的周转过程,研究者提出了多种有机碳稳定机制模型和分组方法[5–6],其中Six等[7]根据有机碳的物理保护机制提出的物理分组方法被越来越多的研究者采用。另一方面,有机碳的矿化过程主要是由土壤微生物参与和驱动的,有机碳组分的含量和性质不同,会影响微生物的生物量和群落特征,进而造成有机碳矿化的差异[8]。因此,明确不同组分有机碳的矿化特征及其微生物驱动过程,对于深入理解不同施肥管理措施下土壤的固碳机制和二氧化碳减排具有重要意义。
有研究表明,长期施肥显著影响土壤总有机碳及有机碳组分的含量。安永齐等[9]在山西矿区复垦土壤上的研究指出,单施有机肥或化肥配施有机肥能够快速提高土壤有机碳含量,且主要提升了大团聚体中游离活性态和物理保护态有机碳比例。稳定性不同的碳组分矿化过程必然有明显的差异。对贵州黄壤长期定位施肥后不同粒径土壤有机碳矿化过程的研究发现,粉粒有机碳和微团聚体有机碳在培养前期矿化速率较高,粗颗粒有机碳矿化速率最低,微团聚体和粉粒的有机碳累积矿化量也显著高于粗颗粒和黏粒[10]。微生物作为有机碳矿化过程的主要参与者受长期施肥和有机碳稳定性的显著影响。有研究表明,施用有机肥不仅可提高土壤肥力,还能增加土壤微生物群落多样性[11],进而促进其功能多样性和群落稳定性[12]。目前关于全土有机碳矿化的研究已多见报道,但对于长期施肥条件下不同有机碳组分的矿化过程却知之甚少,限制了对长期不同施肥下土壤碳的稳定性差异及其机制的理解和认识;对有机碳组分矿化过程中微生物群落的响应和驱动机制更是鲜见报道。
本研究依托典型红壤长期定位施肥试验,采用室内培养方法研究土壤有机碳组分矿化过程的动态特征,并利用PLFA技术深入分析有机碳矿化过程中的微生物群落多样性及其结构差异,明确微生物群落结构特征对红壤不同组分有机碳矿化的贡献,以期深入揭示不同施肥下有机碳在红壤中的固存过程和驱动机制,为肥料合理施用、减少农业生产的环境负效应提供理论依据。
1 材料与方法
1.1 试验区概况
供试土壤采自中国农业科学院祁阳国家红壤肥力长期试验监测站 (111°52′E,26°45′N)的长期定位施肥试验地。该试验站地处中亚热带,年平均气温18℃,年降水量1225 mm,无霜期约300天,年日照时数1610 h。试验地土壤为旱地红壤,成土母质为第四纪红土,是我国亚热带地区具有富铝化特征的地带性土壤。该长期试验起始于1990年,试验初始时耕层土壤(0—20 cm)主要化学性质为:有机质 11.5 g/kg,全氮 1.07 g/kg,全磷 1.03 g/kg,碱解氮 79 mg/kg,有效磷 10.8 mg/kg,速效钾 122 mg/kg,pH 5.7。试验设置及详细田间管理详见文献[13]。本研究选取其中4个处理:撂荒(CK0)、不施肥(CK)、常量氮磷钾化肥(NPK)、常量氮磷钾+常量有机肥(NPKM)。其中撂荒是指自1990年试验初始时,该处理休闲,使其恢复自然演替过程,不收割清理凋落物。目前该处理样方已演替至有大量灌木和乔木生长阶段。
1.2 土壤样品采集与有机碳分组
土壤样品采集于2019年的9月玉米收获后。田间用“S”型布点法采集耕层(0—20 cm)土壤,剔除可见根系与石头,实验室风干,过5 mm筛备用。
土样有机碳组分的分离采用改进的Six等[7]、佟小刚等[14]的方法。取50 g风干土样放入用水湿润的套筛(上层孔径250 μm,下层孔径53 μm,直径18 cm)。用土壤团聚体分析仪(XY-100)按照2 s/次的震速湿筛30 min。震荡结束后,收集粗颗粒 (粒径>250 μm)有机碳组分(cPOC)、微团聚体(粒径53~250 μm)和黏粉粒(粒径<53 μm)有机碳组分。将微团聚体内的组分进一步用密度浮选法分离:取烘干的微团聚体,按照土液比1∶10加入密度1.8 g/cm3的碘化钠 (NaI)溶液,2500 r/min 离心 60 min,收集上清液中的细颗粒有机碳组分(fPOC)。重组部分加入5 g/L的六偏磷酸钠溶液,分散过53 μm筛,粒径>53 μm为团聚体内颗粒有机碳(iPOC),粒径<53 μm的黏粉粒与前述该部分混合为矿物结合态有机碳(MOC)。
将未分组的全土和烘干后的各组分研磨过0.15 mm筛,采用EA3000型元素分析仪测定土壤有机碳和全氮含量。
1.3 室内培养试验
将cPOC、fPOC、iPOC、MOC这4个有机碳组分,按照Zhang等[15]的方法进行室内培养,分析其有机碳矿化特征。将50 g鲜土加入到500 mL去离子水中,手摇1 min后,过45 μm滤膜,获得接种液(内含微生物)。称取4 g分组后有机碳样品与16 g石英砂混合(质量比1∶4),加入1.71 g新制备的鲜土样接种液,使其质量含水量为30%。混合均匀后放入气密性良好的培养瓶中,置于25℃培养箱中预培养7天。
预培养完成后,向培养瓶中放入盛有10 mL 0.5 mol/L氢氧化钠溶液的小烧杯,用于吸收CO2,快速加盖密封培养瓶。25℃恒温培养24 h后取出碱液立即用去离子水稀释5倍,封口,用TOC自动分析仪(Phoenix 8000)测定所吸收CO2的量。根据24 h内的CO2吸收量分别计算第1、2、3、4、6、8、10、12、15、18、25、32、46、75、95、125 天的平均有机碳矿化速率。
土壤有机碳矿化速率和累积矿化量的计算如下:

式(1)中,F为土壤有机碳矿化速率[C mg/(kg·d)],ΔC为待测液上机实测C含量值(mg/L),V为待测液体积(L),i为碱液稀释倍数,m为样品干重(kg),t为培养时间(h)。

式(2)中,Ctotal为土壤有机碳累积矿化量(C mg/kg),F 为土壤有机碳矿化速率 [C mg/(kg·d)],i为采样次数,t为取样的时间。
土壤有机碳矿化过程用一级动力学方程拟合:

式(3)中,Ct是经过t时间后矿化CO2-C的累积释放量;C0代表有机碳的潜在可矿化碳库容量(C mg/kg);e为自然常数,取值2.718;k为周转速率常数(周转数/d);t为培养时间。
1.4 土壤有机碳组分磷脂脂肪酸(PLFA)分析方法
在本研究中,有机碳矿化速率在培养5~8天达到峰值,培养15 天时基本达到稳定。因此,在正式培养到第15天时取不同施肥处理的有机碳组分立即放入圆形铝盒中,置于冷冻干燥机进行真空冷冻干燥24 h,取出后密封置于−80℃冰箱保存,采用Zhang等[16]的方法并进行改进测定磷脂脂肪酸。主要操作为:取3 g冷冻干燥后的样品,依次进行磷脂的提取、分离和甲酯化,获得磷脂脂肪酸甲酯后以十九烷酸甲酯(C19:00)为内标,在安捷伦Agilent 6890N气相色谱仪上,采用美国MIDI Sherlock微生物鉴定系统平台的PLFA模块进行磷脂脂肪酸鉴定。根据PLFA数据分析其微生物生物量,并计算其香农指数(Shannon index)、辛普森指数 (Simpson index) 和均匀度指数 (Pielou index) 用以表征微生物群落结构多样性和物种分布均匀度,计算方法见文献[17]。
1.5 数据分析方法
试验结果统计与分析采用SPSS Statistics 25.0软件进行,数据测定结果均以平均值±标准误(mean±SE)表示。数据正态性和方差齐性检验后采用单因素方差分析(One-way ANOVA)结合邓肯(Duncan)法进行差异显著性检验(P<0.05为差异显著)。冗余分析(RDA)采用canoco5完成。动力学方程拟合和数据绘图采用 SigmaPlot 14.0 和 Origin 2021 完成。
2 结果与分析
2.1 长期不同施肥下红壤有机碳及其组分变化
从表1可见,各个施肥处理下土壤各有机碳组分均以矿物结合态有机碳(MOC)含量最高,为8.77~9.69 g/kg,占土壤有机碳(SOC)的68.8%~94.5%,其次为粗颗粒有机碳(cPOC)和团聚体内颗粒有机碳(iPOC),细颗粒有机碳(fPOC)含量最低。相较于CK,各处理cPOC、fPOC和iPOC (除NPK外)组分占比均有显著提升,分别提高了4.02~6.56、2.01~2.29和2.07~2.26个百分点;而矿物结合态有机碳(MOC)组分占比有明显的下降趋势,在CK0、NPK和NPKM施肥下分别下降了24.8、11.8和25.7个百分点。这表明长期撂荒和施肥均能够增加土壤中活性有机碳的含量。
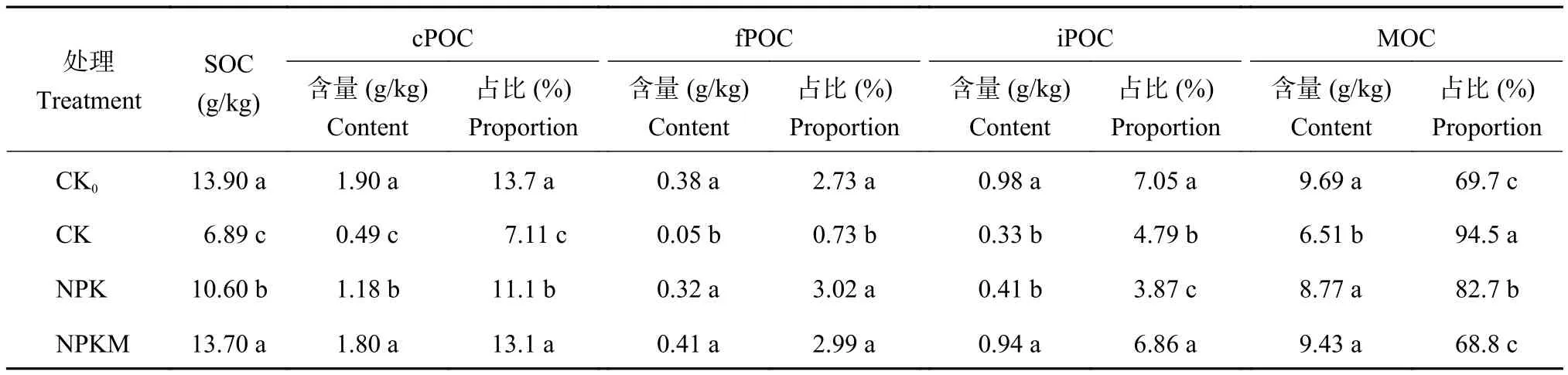
表1 长期不同施肥下红壤有机碳组分含量及占比Table 1 Contents and proportions of organic carbon fractions in red soil under different long-term fertilization
与不施肥相比,撂荒与长期施肥均显著提升了红壤SOC及各组分碳(NPK处理iPOC除外)的含量。CK0处理下,土壤SOC含量较CK处理提高了102%,其中,cPOC、fPOC、iPOC和MOC分别显著增加了2.9、6.6、2.0和0.5倍。NPK和NPKM处理土壤SOC含量较CK处理下分别提高了54%和99%,其中除NPK的iPOC组分与CK无显著差异之外,其他组分均显著高于CK处理,说明长期施肥显著增加了各组分有机碳的含量。与NPK相比,NPKM显著提高了cPOC和iPOC的含量,而对fPOC和MOC含量没有显著影响。
2.2 长期施肥条件下各有机碳组分的矿化特征
2.2.1 不同有机碳组分的矿化速率 长期不同施肥下,各组分有机碳矿化速率均表现出前期(矿化1~35天)快速矿化,后期逐渐放缓直至趋于平稳的趋势(图1)。各处理的4个组分中,平均最大矿化速率表现为 fPOC [25.9 mg/(kg·d)]>cPOC[16.0 mg/(kg·d)]>iPOC [14.7 mg/(kg·d)]>MOC [8.7 mg/(kg·d)]。

图1 长期施肥下红壤各组分有机碳的矿化速率Fig. 1 Mineralization rate of organic carbon in organic carbon fractions of red soil after long-term fertilization
与CK相比,CK0处理cPOC、fPOC、iPOC和MOC组分的最大矿化速率分别提高了158.0%、36.4%、67.3%和146.0%,NPKM处理分别提高了246.0%、62.9%、21.4%和183.0%。可见,撂荒和有机无机肥配施处理显著提升了各有机碳组分的最大矿化速率,尤其是cPOC和MOC组分。NPK处理下各组分最大有机碳矿化速率除MOC外,均与CK处理下无显著差异。与NPK相比,NPKM处理cPOC、fPOC、iPOC和MOC组分的最大有机碳矿化速率分别提高了240%、76.7%、48.2%和35%。
2.2.2 不同有机碳组分的矿化动力学参数 长期不同施肥下,红壤有机碳组分的累积矿化量随时间的动态变化可以用一级动力学方程 Ct=C0(1−e−kt) 拟合,其R2均在0.97以上,达到极显著水平(P<0.01),说明该方程可以很好地描述红壤各有机碳组分矿化的动力学过程(表2)。相较于CK处理,CK0、NPK和NPKM处理的潜在可矿化碳库(C0)容量都有所提升,平均分别提高了351.7、117.2和559.7 mg/kg,尤其是NPKM处理,其cPOC、fPOC、iPOC和MOC组分的潜在可矿化有机碳含量(C0)分别比对照处理提高了59.7%、46.8%、137%和432%。各组分中fPOC的潜在可矿化碳含量最高,平均为1093 C mg/kg,cPOC、iPOC和MOC含量相近,为608~669 mg/kg。

表2 土壤各有机碳组分矿化的动力学参数Table 2 The kinetic parameters for soil organic carbon mineralization of each fractions
不同有机碳组分的矿化率表现为iPOC (5.3%)>MOC (4.1%)>cPOC (3.6%)>fPOC (0.56%)。不同施肥处理下各有机碳组分库的周转速率常数(k)变化范围为0.007~0.022,各组分平均周转速率表现为fPOCC>cPO>iPOC>MOC,这与观察到的最大矿化速率趋势一致。
2.3 长期施肥条件下有机碳组分矿化过程中的微生物群落结构特征
2.3.1 不同组分有机碳矿化过程中的磷脂脂肪酸含量
长期施肥条件下,各有机碳组分快速矿化阶段(矿化15天)的总PLFA含量如图2所示。各组分中,fPOC组分矿化过程中的总PLFA含量最高(平均94.5 nmol/g),约为其他组分 (平均26.1~35.0 nmol/g)的3倍。与CK处理相比,NPK和NPKM处理的PLFA含量在cPOC中分别显著提高了29.7%和21.7%;在MOC组分中分别显著提高了38.3%和17.2%;而CK0处理只在MOC组分中显著提高了41.9%,其他组分无显著差异。NPK和NPKM处理下各组分的PLFA含量均无显著差异。

图2 土壤有机碳组分矿化15天时各施肥处理的总PLFA含量Fig. 2 Total PLFA contents of each soil organic C fraction after mineralized for 15 days
2.3.2 不同组分有机碳矿化过程中的微生物多样性指数 长期不同施肥条件下,有机碳组分矿化过程中微生物群落多样性指数见表3。从各处理的平均值来看,香农指数在fPOC组分中最高,其次为iPOC组分中,在cPOC和MOC组分中最低。辛普森指数在fPOC和iPOC中最高,在cPOC中其次,在MOC中最低。均匀度指数在iPOC中最高,其次为fPOC和MOC,在cPOC中最低。iPOC组分中CK0、NPK、NPKM处理的香农指数分别为1.30、1.27和1.31,与CK相比,显著增高了23.3%~27.2%,均匀度指数分别增加了6.0%、10.5%和28.4%,其余各处理之间差异不显著。
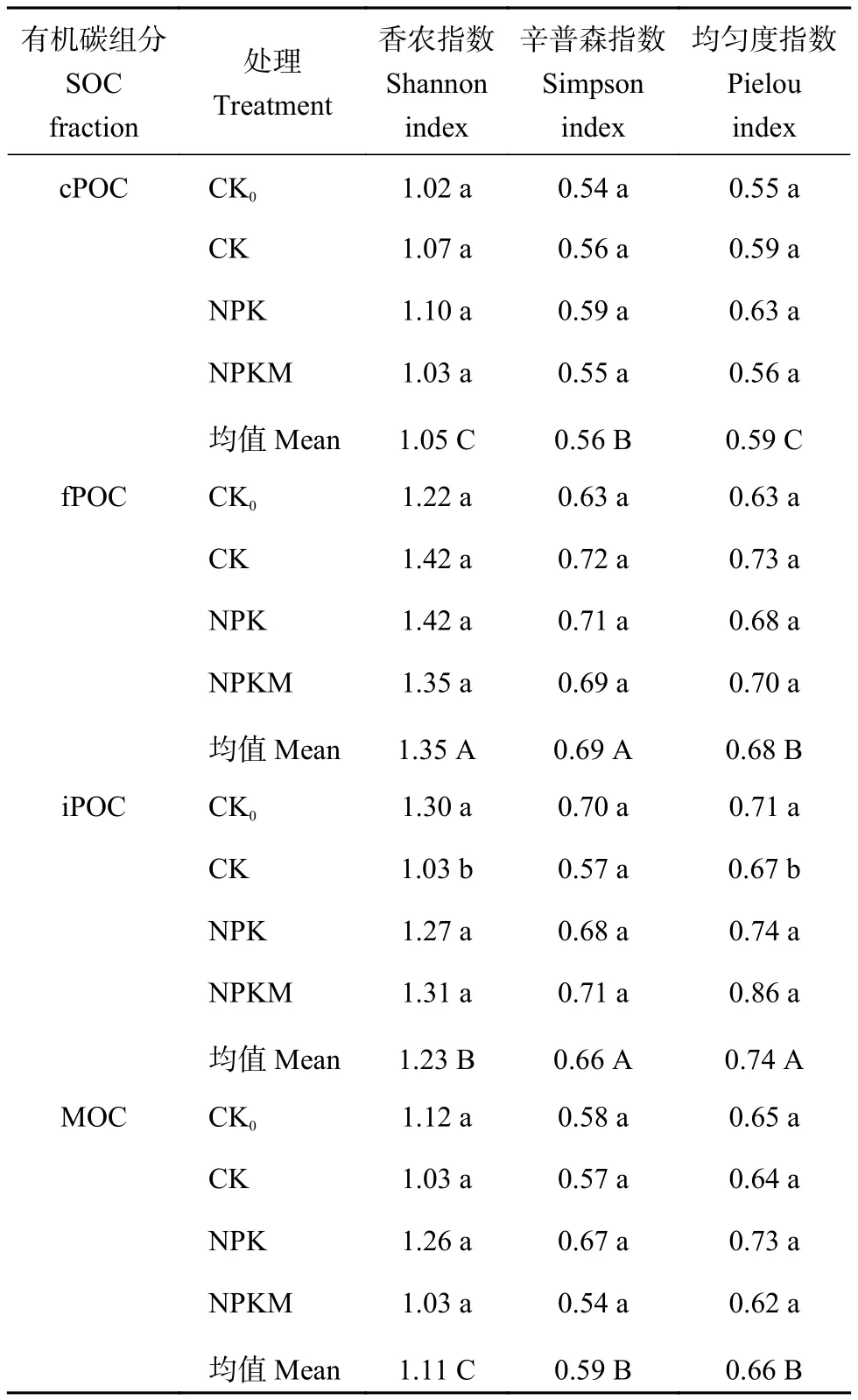
表3 土壤各有机碳组分矿化过程中的微生物群落多样性指数Table 3 Diversity index of soil microbial community in each soil organic carbon fraction during mineralization
2.3.3 不同组分有机碳矿化过程中的微生物群落结构组成 图3为不同组分有机碳快速矿化15天时革兰氏阳性菌与阴性菌比例(G+/G−)和真菌与细菌比(F/B)。总体来说,G+/G−在 MOC (6.06)与 cPOC(5.18)中较高,fPOC (1.26)和 iPOC (1.03)中较低。与CK相比,CK0、NPK和NPKM处理均能显著提高cPOC组分的G+/G−,分别提高了 23.0%、23.4%和32.0%,而MOC组分中的G+/G−显著降低了44.9%、78.3%和16.7%。fPOC中G+/G−在CK0和NPK处理下显著高于CK和NPKM处理下。iPOC中的G+/G−在各处理间无显著差异。
与 G+/G−相反,F/B 在 iPOC (0.11)和 fPOC (0.10)中较高,在 cPOC (0.03)和 MOC (0.06)中较低。iPOC中的F/B在NPKM处理下较其他处理显著高71.9%~142.9%。fPOC中的F/B在NPK处理下较其他处理显著低23.7%~46.5%。cPOC和MOC组分的F/B在各施肥处理之间无显著差异。
2.4 不同组分有机碳矿化的影响因素分析
将土壤矿化特征与快速矿化阶段的PLFA相关指标进行冗余分析,可明晰微生物群落结构与有机碳组分矿化之间的关系。图4a中,RDA1和RDA2轴对有机碳矿化的解释率达到了96.4%,其中RDA1轴的解释度为84.2%,RDA2轴解释度为12.2%。由矿化特征与微生物群落指标之间的夹角可知,组分矿化率与G+/G−、F/B、均匀度指数呈正相关,而与其他微生物相关因子呈负相关;潜在可矿化碳、累积矿化量和最大矿化速率与真菌、细菌、革兰氏阳性菌、革兰氏阴性菌及香农指数等呈正相关,与F/B、均匀度指数正相关性较弱,而与G+/G−呈负相关。由冗余分析结果 (图4b) 可知,总PLFA含量对有机碳矿化的解释度为54.2%,是对长期施肥条件下有机碳组分矿化影响最显著的因子;其次为真菌生物量(6.7%)、革兰氏阴性菌生物量(4.1%)和均匀度指数 (Pielou index,3%)。
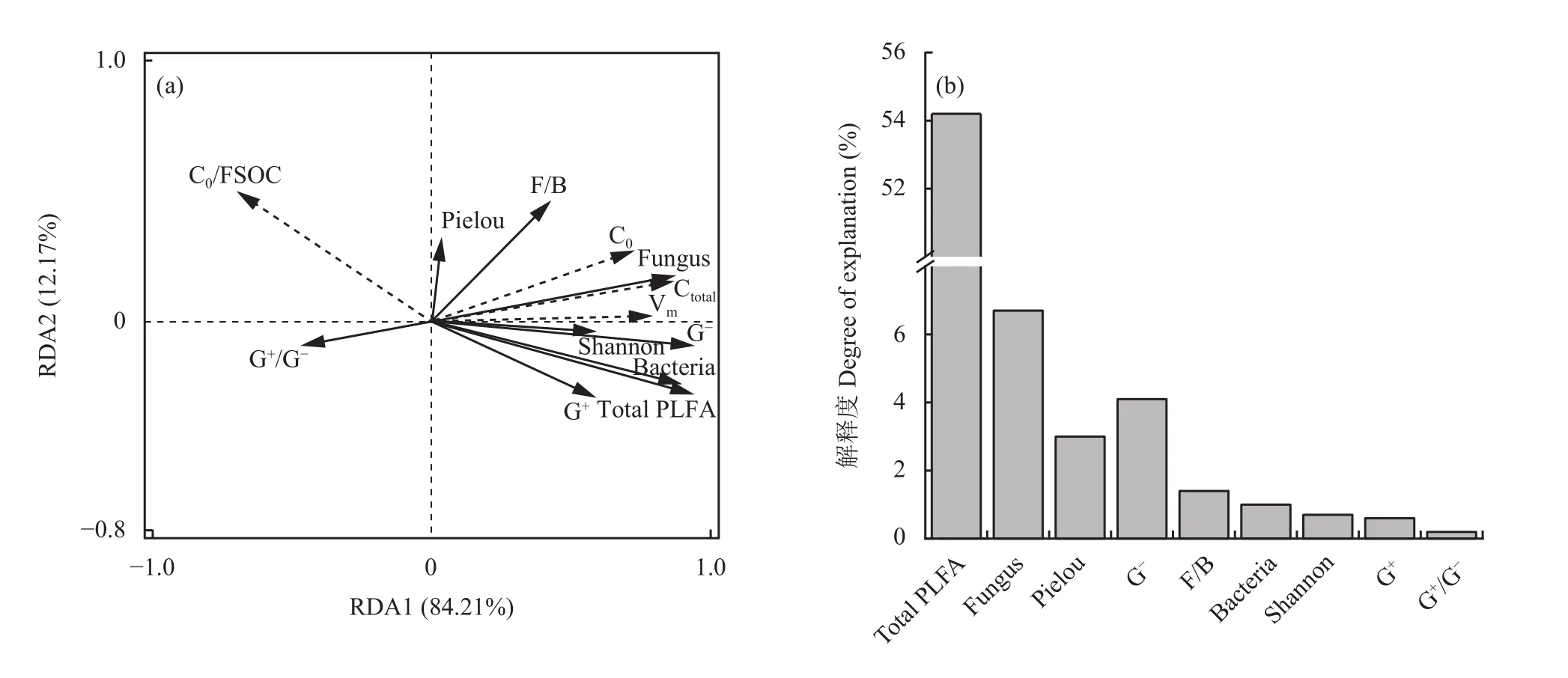
图4 土壤微生物群落结构对不同组分有机碳矿化特征的影响Fig. 4 Effects of soil microbial community structure on mineralization characteristics of different organic carbon fractions
3 讨论
3.1 长期不同施肥下红壤有机碳组分的矿化特征
土壤有机碳含量的变化主要取决于外源有机碳输入与输出之间的平衡。前人的研究表明,合理的施肥措施可以有效的增加红壤有机碳含量及其活性[18–19]。本研究结果也表明,长期施肥较不施肥不仅提高了土壤有机碳含量,还改变了土壤有机碳在组分中的分配比例。尤其是NPKM处理,相较于CK,其cPOC、fPOC、iPOC等活性有机碳组分的占比都有一定的提升,但惰性有机碳组分MOC的占比有所下降。其原因一方面是施肥增加了碳投入改善了土壤质量,使更多的有机碳截留在土壤中,提高了土壤有机碳水平[20];另一方面是活性有机碳组分主要来源于作物残茬、有机残留和有机质的初级分解产物等,是土壤中相对“较新”的有机碳[14],经过29年的不同施肥,土壤有机碳优先富集在这些活性组分中,从而提升了活性有机碳组分的占比。
有机碳矿化速率和累积矿化量的大小反映了土壤有机碳矿化的快慢程度和强度(图1、表2)。本研究中各有机碳组分的矿化速率表现为前期快、后期慢的特点。这主要是由于有机碳组分矿化前期含有较多的糖类、蛋白质等易被微生物分解利用的碳源,矿化速率较高[21]。随着矿化过程的进行,易矿化的碳源消耗殆尽,逐渐变成以纤维素、木质素等较难分解的碳为主,矿化速率逐渐下降至一个相对平稳的状态[22]。矿化率指矿化作用所消耗的土壤有机碳的比例,能够在一定程度上反映有机碳组分的可矿化潜力。本研究结果表明, iPOC组分的矿化率最高,这可能是因为该组分具有较低的C/N值[14],是易于被微生物利用的优质碳源[23],长期来看具有一定的可矿化潜力。
撂荒和有机肥处理下各有机碳组分的最大矿化速率和潜在可矿化碳库容量明显高于CK,而单施化肥时cPOC、fPOC、iPOC有机碳组分的变化不大。这可能是因为,长期不施肥处理条件下,土壤有机碳尤其是易矿化有机碳组分的含量在不断降低,而撂荒处理下自然演替过程的发展,使植物多样性和凋落物数量不断增加,进而增加了有机物投入,使土壤有机碳的含量增加[24]。而有机无机肥配施处理也增加了土壤碳含量,改善了土壤养分状况,更有利于微生物的生长,进而促进了有机碳的矿化[25–26]。而单施化肥处理,一定程度上加剧了土壤酸化,破坏了土壤良好的结构,从而对微生物群落产生了不利影响[27]。表3中iPOC的Shannon指数在撂荒和有机无机肥配施处理下均显著高于不施肥处理,也说明了这两个施肥处理能够显著提高土壤微生物多样性。此外,真菌被认为是主要分解难分解、具有高碳氮比的有机物[28],而有机无机肥配施处理显著提高了土壤中的真菌细菌比(图3),说明施用有机肥显著提高了土壤中微生物矿化分解复杂有机物的能力,这可能是有机无机肥处理矿化速率较高的原因之一。本研究中各组分的最大矿化速率和潜在可矿化量均表现为fPOC>cPOC>iPOC>MOC,进一步验证了fPOC、cPOC、iPOC组分中含有更多的活性碳源,是土壤有机碳中较活跃的碳库。
3.2 不同组分有机碳矿化过程中的微生物群落结构
微生物是有机碳矿化的主要参与者和驱动力,不同有机碳组分矿化过程中的微生物群落特征有所差异。本研究中不同有机碳组分矿化过程中总PLFA生物量表现为fPOC>cPOC>MOC>iPOC,尤其是fPOC组分明显高于其他组分。其原因可能是fPOC组分主要由高活性碳源组成[29],微生物能优先利用这些简单的碳大量增殖,这也进一步揭示了fPOC组分最大矿化速率和累积矿化量较高的原因。多样性指数、F/B和G+/G−常作为研究微生物群落结构和多样性的指标,其中多样性指数是群落结构和功能多样性的表征[17];F/B能够反映微生物群落的稳定性,且比例越高越稳定[30];G+/G−指示土壤养分状况,比值越高,表示营养胁迫越强[31]。本研究中iPOC和fPOC组分F/B的值更高,其香农指数、辛普森指数和均匀度指数也都高于其他组分,而G+/G−值明显低于cPOC、MOC组分。说明矿化过程中其群落结构更稳定,受到的养分胁迫较小,微生物群落多样性也更高。其原因可能是fPOC、iPOC组分是土壤有机碳转化的中间碳库,有机碳活性高且具有更适合微生物生长的化学计量比,从而形成了更稳定多样的微生物群落结构。
土壤有机碳矿化受多种因素影响,冗余分析结果表明,总PLFA含量、真菌PLFA和革兰氏阴性菌量是对组分碳矿化影响最大的3个因素。其中总PLFA含量决定了参与矿化的微生物总量,冗余分析也显示两者有显著的正相关关系。本研究中施肥处理较不施肥处理显著提高了cPOC和MOC中总PLFA含量(图2),因此促进了这两个组分中的有机碳矿化。真菌与G−菌是土壤中主要的“挖掘”微生物,对有机碳的矿化起了关键作用。潜在矿化率与G+/G−、F/B及均匀度指数呈正相关,表明了较高的群落物种均匀度、革兰氏阳性菌和阴性菌比例以及真菌细菌比,可提高土壤有机碳的可矿化程度。可见,对于复杂的微生物种群结构来说,不同种群之间的配合和结构合理性是调控有机碳固存和分解之间平衡的重要因素,在土壤和肥料管理中应给予高度重视。另外,目前对土壤细菌的研究较多,但真菌的数量和群落结构组成均对有机碳矿化起到重要调控作用,未来研究工作中充分挖掘有机碳调控真菌种群和作用机制,将进一步加深对碳循环及其环境行为的认知。
4 结论
1)长期施肥不仅提高了红壤总有机碳含量,还提升了有机碳中活性碳组分(cPOC、fPOC、iPOC)的占比,活化了土壤有机碳。
2)长期施肥条件下,轻组有机碳(fPOC)组分中潜在可矿化碳含量最高,是土壤中有机碳活性较强的组分。有机肥配施化肥和撂荒均能提高各有机碳组分的潜在可矿化碳库容量,长期施用化肥仅提高了矿物结合有机碳的潜在可矿化量,对其他组分没有影响。
3)fPOC组分的微生物量也明显高于其他组分,这可能是其矿化能力更强的原因之一。土壤有机碳的可矿化程度与微生物群落结构及群落的物种均匀度密切相关。长期施用化肥和有机肥均显著提高了iPOC组分中的微生物多样性和均匀度,使其群落结构更稳定,这可能是其矿化率较高的原因之一。
