毛竹入侵对杉木人工林表层土壤真菌多样性及群落结构的影响1)
2023-02-22江思源程雪飞张金池唐瀛洲聂晖王宇浩刘京
江思源 程雪飞 张金池 唐瀛洲 聂晖 王宇浩 刘京
(南方现代林业协同创新中心(南京林业大学),南京,210037)
毛竹(Phyllostachysedulis)禾本科竹亚科刚竹属,广泛分布于亚热带地区[1]。因毛竹具有独特的地下茎(竹鞭),可以横向快速蔓延实现种群克隆扩张[2],是典型的本土入侵物种,对临近林分的植物物种多样性构成严重威胁[3]。在东亚日本和南美洲的亚热带地区,已经广泛观察到毛竹对邻近阔叶林或针叶林的入侵[4]。目前,我国毛竹正在向邻近森林扩张,据估计,近几十年来毛竹入侵地区面积每年以3%的速度扩大[5]。毛竹的入侵导致其他植物物种被取代,在过渡区形成混交林[6]。随着入侵的进行,甚至替代原有的森林形成毛竹纯林[7]。这导致森林物种生物多样性和生态系统稳定性恶化的危险性增加[8]。杉木(Cunninghamialanceolate)为中国亚热带地区主要用材树种,其面积与蓄积量分别约占我国主要优势人工林树种的四分之一及三分之一[9],其人工林在长江流域和秦岭以南地区广泛栽培,但集约化经营导致杉木人工林群落结构简单,极易受到毛竹的入侵[10],造成杉木人工林生产力下降、地力衰退,并且严重制约林业经济发展[11]。
土壤微生物是土壤生态系统的重要组成部分,也是生态系统中最活跃的生态因素之一[12]。植被-土壤-微生物是一个相互影响的有机体。事实上,入侵物种往往具有更强的土壤养分利用能力和更高的生物量产量[13]。因此,当发生外界植物入侵时,入侵种会改变侵域土壤中的理化性质和养分结构等[14],土壤微生物群落结构和多样性也会受到影响[15]。以往的研究发现毛竹入侵会显著影响天然阔叶林土壤的微生物群落,且真菌群落对植被转换的响应比细菌更敏感[16]。这些影响可能是由于入侵树种在根系分泌物、凋落物质量和养分利用等方面差异造成的[17]。土壤真菌作为土壤微生物的重要类型之一,能够分解凋落物、植物残株及土壤中的有机质[18],在土壤生态系统的养分循环和能量流动过程中扮演重要角色[19]。因此,通过研究毛竹入侵对表层土壤真菌多样性和群落结构的影响,可以更好的认识植物入侵对土壤地下生态系统的影响。
目前,关于毛竹入侵对森林土壤微生物群落结构和多样性的研究多使用传统微生物研究方法,如微平板分析(Biolog)和磷脂脂肪酸(PLFA)分析法等[20],尚未有研究运用高通量测序技术对毛竹入侵杉木人工林后土壤真菌群落结构进行分析。鉴于此,本研究以杉木人工林、杉竹混交林和毛竹纯林3种入侵程度林分表层土壤为研究对象,基于ITS rDNA的Illumina Miseq高通量测序技术分析土壤真菌群落变化,探讨毛竹入侵杉木人工林对表层土壤真菌结构和多样性的影响,揭示毛竹入侵人工林对表层土壤真菌群落生态稳定性的影响,为合理评价和治理该入侵系统提供数据参考,也为深入了解毛竹的入侵机制提供理论依据。
1 研究区概况
试验样地位于南京林业大学下蜀林场,林场属江苏省句容市下蜀镇内,东经119°14′,北纬31°59′,总面积3 891 hm2。属北亚热带季风气候区、干湿寒暑分明、光照充足、水热资源丰富,具有发展林业的良好条件。据句容市气象站和林场多年观测资料,年平均气温15.5 ℃,年平均降水量1 099.1 mm,但实际变化较大,降水以夏季最多,其次为春季和秋季,年平均空气相对湿度为79%,平均无霜期233 d。林场内土壤以黄棕壤和山地黄棕壤为主,土层厚度一般在50 cm以上。林场内人工林主要有:杉木林、马尾松林、火炬松林和毛竹林等[21]。在研究区域,毛竹最初是由当地人种植的,人为干扰(如收获竹笋)促进了竹子的快速生长及其对邻近森林的入侵。杉木人工林经历了毛竹入侵,入侵后形成的毛竹林面积约占研究区域总面积的五分之一。三种不同的森林类型代表毛竹入侵导致的演替阶段,包括杉木林(针叶林,未入侵原生林,林龄30 a),杉竹混交林(混交林,称为毛竹中度入侵的过渡林,毛竹入侵15 a)和毛竹林(竹林,在毛竹入侵后形成;毛竹严重入侵,毛竹入侵25 a)。
2 研究方法
2.1 土壤样品采集与处理
按照毛竹扩张水平方向依次选择杉木林、杉竹混交林和毛竹林样地,每种林分设置3个10 m×10 m的样地,每个样地之间相距20 m,共计9个样地。于2021年4月在每块样地中选取5个代表性样点,去除地表凋落物后,采集0~10 cm土壤样品,过2 mm筛后均匀混合组成一个混合样,3种林分共得到9个混合土样。带回实验室将新鲜土壤分为两份,一份立即冷冻干燥,用于提取土壤微生物总DNA,供真菌群落分析;另一份室内自然风干,研磨过筛后用于土壤基本理化性质分析。
2.2 土壤理化性质测定

2.3 土壤微生物测定
采用广州美基生物科技有限公司的D3142试剂盒土壤微生物DNA进行提取,利用NanoDrop微量分光光度计和琼脂糖凝胶电泳仪对提取的DNA产物进行检测,检测合格的DNA用带有barcode的特异引物扩增ITS的ITS2区。引物序列为:ITS3_KYO2:GATGAAGAACGYAGYRAA;ITS4:TCCTCCGCTTATTGATATGC。PCR反应经过两轮扩增,使用AMPure XP Beads对第二轮扩增产物进行纯化,将纯化后的扩增产物(即扩增子)连接测序接头,构建测序文库,Illumina上机测序。用ABI Step One Plus Real Time PCR System(Life Technologies,产地美国)进行定量,根据Novaseq6000的PE250模式pooling上机测序。
2.4 数据处理
利用Usearch软件按照97%相似度对土壤中OTU进行聚类分析,对其进行丰度与多样性指数分析,采取Chaol指数和Ace指数表征土壤中真菌群落的丰富度,Shannon指数表征真菌群落的多样性;分别运用R语言进行土壤真菌Anosim检验和PCoA聚类分析;运用Lefse1.0软件分析组间差异;利用Excel 2016等软件进行数据处理,运用SPSS 23.0统计软件进行方差分析与相关性分析。
3 结果与分析
3.1 毛竹入侵对表层土壤理化性质的影响
如表1所示,毛竹入侵对杉木人工林表层土壤理化性质产生了不同的影响。毛竹入侵杉木人工林后形成毛竹纯林后,土壤含水量显著减少,毛管孔隙度先增加后减少,非毛管孔隙度先减少后增加。土壤硝态氮质量分数和有机碳质量分数均随毛竹入侵程度的加深逐渐减少,三者间硝态氮质量分数差异显著,但三者间有机碳质量分数未达到显著差异。铵态氮质量分数随入侵的加深逐渐增多,毛竹纯林的铵态氮质量分数显著高于杉木人工林和杉竹混交林。入侵前杉木人工林的有效磷质量分数显著大于杉竹混交林和毛竹纯林。此外,毛竹入侵过程中形成的杉竹混交林pH显著高于杉木人工林和毛竹林。

表1 不同林分类型土壤理化性质
3.2 毛竹入侵对表层土壤真菌群落组成结构的影响
在本研究的所有土壤样品中,经高通量测序得到有效序列698 500条,可分类OTU共2 325个(见图1),所有土壤样品Tags中识别到可注释真菌类群达到5界14门49纲137目246科375属347种。不同林分类型的OTU种类数目各不相同:杉木人工林830,杉竹混交林为937,毛竹纯林为1012。三种林分共同包含的OTU有321个,占总OTU的13.8%。杉木人工林、杉竹混交林和毛竹纯林土壤中特有的OTU分别为315个、347个和460个,分别占总OTU数量的13.55%、14.92%和19.78%。

图1 不同林分类型表层土壤真菌OTU韦恩图
3.2.1 门水平的真菌群落组成结构
表2为门水平下真菌群落物种分布堆叠图,检测出主要类群为子囊菌门(Ascomycota)、担子菌门(Basidiomycota)和被孢霉门(Mortierellomycota)等,相对丰度大于1%。杉木人工林在门水平上的优势类群分别为子囊菌门(38.89±17.13)%、担子菌门(30.24±18.03)%、被孢霉门(5.08±4.19)%和壶菌门(Chytridiomycota,1.54±1.44)%。杉竹混交林的优势类群分别为子囊菌门(66.18±3.15)%、担子菌门(13.68±3.96)%、被孢霉门(6.50±1.64)%和毛霉门(Mucoromycota,1.46±0.42)%。毛竹纯林在门水平上的优势类群分别为子囊菌门(60.46±2.80)%、担子菌门(26.26±5.42)%和被孢霉门(5.63±2.33)%。子囊菌门的相对丰度由大到小依次为杉竹混交林(66.18%)、毛竹纯林(60.46%)、杉木人工林(38.89%)。担子菌的相对丰度由大到小依次为杉木人工林(30.24%)、毛竹纯林(26.26%)、杉竹混交林(13.68%),被孢霉门的相对丰度由大到小依次为杉竹混交林(6.50%)、毛竹纯林(5.63%)、杉木人工林(5.08%),杉竹混交林土壤中未分类的真菌群落相对丰度最高,其次是毛竹纯林和杉木人工林。在门水平上,子囊菌门和担子菌门的相对丰度在毛竹入侵后变化较大。
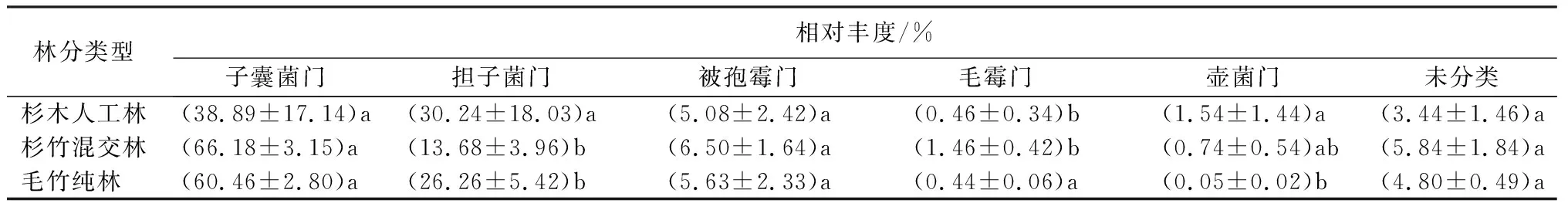
表2 毛竹入侵前后土壤真菌门水平群落组成
3.2.2 属水平的真菌群落组成结构
表3为属水平下真菌群落物种分布堆叠图,检测出主要真菌种群为圆孢霉属(Staphylotrichum)、被孢霉属(Mortierella)、沙蜥属(Saitozyma)、木霉属(Trichoderma)、粗糙孔菌属(Trechispora)、湿伞属(Hygrocybe)和Myrmecridium等。毛竹入侵改变了一些真菌属的相对丰度,其中圆孢霉属相对丰度升高,为杉木人工林(1.64%)、杉竹混交林(7.41%)、毛竹纯林(8.47%)。木霉属(Trichoderma)和小菇属(Mycena)相对丰度均随着毛竹入侵显著增加。杉竹混交林中被孢霉属、湿伞属、青霉属(Penicillium)和未分类在三种林分中相对丰度最高。杉木人工林中粗糙孔菌属和Myrmecridium的相对丰度显著高于有毛竹入侵的林分,可能是杉木林的优势属。毛竹纯林中沙蜥属(Saitozyma)相对丰度(7.80%)显著高于杉木人工林(2.07%)和杉竹混交林(1.83%)。
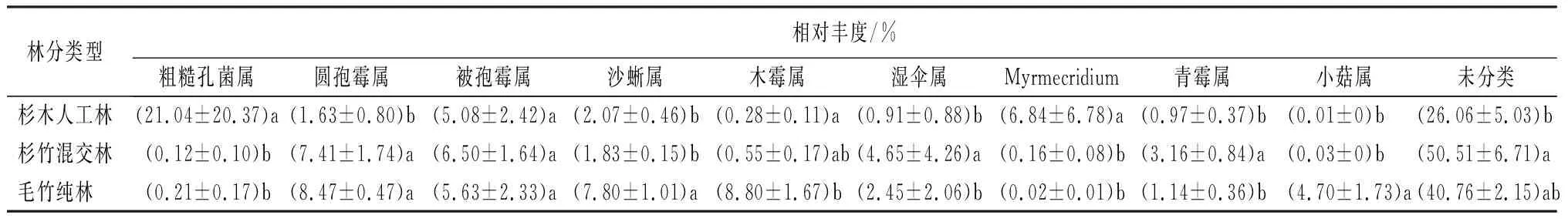
表3 毛竹入侵前后土壤真菌属水平群落组成
3.3 毛竹入侵对表层土壤真菌多样性的影响
本试验通过OTU数、Chao1和ACE指数反映物种的丰富度信息;Shannon和Simpson指数体现物种的丰富度和均匀度,PD指数结合物种进化距离反映物种多样性。总的来说,通过这些指数进行样本的α物种多样性分析。从表4可知,杉木人工林、杉竹混交林和毛竹纯林的OTU数分别为667、774和835个。说明杉木人工林样地真菌群落在总体数量上处于弱势。Shannon指数数值越大,表明群落的多样性越高。真菌Shannon指数、Simpson指数和PD指数均表现为杉竹混交林>毛竹纯林>杉木人工林。Chao1指数和ACE指数可以评估样品中真菌群落的丰富度,Chao1指数和ACE指数均表现为毛竹纯林>杉竹混交林>杉木人工林,说明毛竹入侵后真菌丰富度有所升高。

表4 毛竹入侵前后土壤真菌的多样性指数
3.4 毛竹入侵条件下表层土壤真菌群落组成的主坐标分析
PCoA是基于样本间距离进行降维分析,平面上点距离越近,表明样本的菌群结构越相似。对三种林分的表层土壤真菌群落在OTU水平上进行基于Bray距离算法的PCoA分析(见图2),主成分1与主成分2分别解释真菌群落变异的23.82%和21.42%,两者累计贡献率为45.24%。杉木人工林和杉竹混交林分布距离较近,相似度高,表现为聚集效应;毛竹纯林在图中较其他林分分布较远,表现出分离效应。同时结合基于bray距离算法的Anosim检验(表5)结果表明,三个林分表层土壤真菌群落结构总体上差异显著,且组间与组内差异极显著,与图3表现一致。

表5 组间真菌群落结构差异Anosim显著性检验
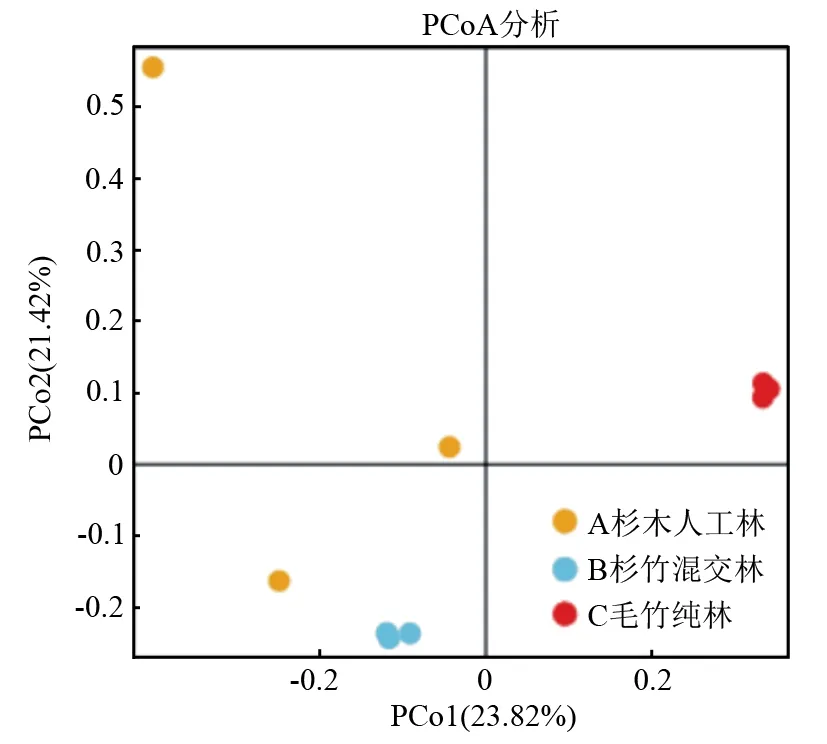
图2 毛竹入侵条件下表层土壤真菌群落结构PCoA分析
3.5 土壤真菌群落结构与环境因子相关性分析
图3为属水平下真菌群落结构与土壤理化性质的相关性热图,沙蜥属(Saitozyma)与土壤中的铵态氮(AN)质量分数和非毛管孔隙(NCP)质量分数呈显著正相关,与土壤中的自然含水率(WC)和毛管孔隙度(CP)呈极显著负相关;木霉属(Trichoderma)与土壤中的铵态氮(AN)质量分数呈极显著正相关,与非毛管孔隙(NCP)质量分数呈显著正相关,与自然含水率(WC)和毛管孔隙度(CP)呈极显著负相关;Myrmecridium与土壤中的速效钾(AK)质量分数、有效磷(AP)和总孔隙度(POR)呈显著正相关,与容重(BD)呈显著负相关;青霉属(Penicillium)与土壤中的pH呈极显著正相关;小菇属(Mycena)与土壤中的铵态氮(AN)质量分数呈极显著正相关,与自然含水率(WC)和毛管孔隙度(CP)呈显著负相关。结合表3可以得到,毛竹入侵后沙蜥属(Saitozyma)、木霉属(Trichoderma)、Myrmecridium、青霉属(Penicillium)和小菇属(Mycena)等菌属相对丰度明显改变,说明毛竹入侵后土壤理化性质的改变是土壤真菌群落结构发生改变的主要原因。

横向为环境因子信息;纵向为物种信息;*表示显著相关(p<0.05);** 表示极显著相关(p<0.01)。
4 讨论
植物入侵通过地上植被的演替来改变土壤微生物的群落结构及多样性[22]。土壤真菌易受到土壤物理性质、土壤化学因子和植被类型的影响[23]。本研究表明,毛竹入侵使土壤真菌多样性升高,其中所形成的杉竹混交林表层土壤真菌多样性最高。多数研究认为土壤微生物多样性与植物群落多样性呈正相关[24],毛竹入侵过程中形成的杉竹混交林相较杉木人工林和毛竹纯林具有更丰富的植物多样性,是该林分表层土壤真菌多样性最高的原因。同时有研究表明,pH是土壤微生物组成和分布的关键因素之一[25],也是土壤真菌群落变化的主要驱动因子[26],杉竹混交林土壤酸性减弱,为入侵提供适宜的环境。毛竹入侵后表层土壤真菌丰富度也显著提高,其中毛竹纯林的表层土壤真菌丰富度最高。
毛竹入侵改变了原有的林分类型,从而改变了土壤真菌的优势菌群[27]。本研究表明,在门水平上,子囊菌门(Ascomycota)和担子菌门(Basidiomycota)的相对丰度变化较大,被孢霉门(Mortierellomycota)相对丰度变化较小。表明子囊菌门与担子菌门对毛竹入侵较为敏感,被孢霉门则相对稳定。子囊菌门真菌多为寄生菌或腐生菌,可以降解土壤中的有机质,且能增强土壤抗侵蚀能力[28]。担子菌门真菌也是土壤中重要的分解者,能产生胞外水解酶和氧化酶参与木质素和纤维素的分解,在碳循环过程发挥重要作用[29]。担子菌门一般存在于通气性良好的土壤中[30]。毛竹林因根系发达,土壤中非毛管孔隙度显著大于未入侵林分,使土壤环境更适合担子菌门的生长。杉木人工林的优势属为粗糙孔菌属(Trechispora)和Myrmecridium,其均为土壤腐生真菌[31],目前仍缺乏与粗糙孔菌属共生植物的研究[32]。本研究结果表明该属真菌可能与杉木林下堆积大量枯落物形成腐生环境有关。杉竹混交林的优势属中假裸囊菌属(Pseudogymnoascus)和青霉属(Penicillium)具有腐生菌的功能[33],对纤维素具有明显降解作用[34]。且青霉属是溶磷微生物[35],因而杉竹混交林的有效磷质量分数最低。毛竹纯林拥有较高丰度的木霉属(Trichoderma)、小菇属(Mycena)和沙蜥属(Saitozyma)与土壤含水率、毛管孔隙度呈显著负相关。木霉属(Trichoderma)真菌有利于提高植物抗病性与抑菌性,通过分泌几丁质酶和纤维素酶水解病原体细胞壁,控制大范围的植物病原体[36]。小菇属(Mycena)对凋落物中的纤维素有较强的分解能力,沙蜥属(Saitozyma)可修复土壤污染,促进土壤中氮、磷、钾元素的矿化。
相关研究表明,土壤真菌群落结构受到土壤理化性质和植被类型的影响,其中土壤水分、有效养分、pH和有机质等土壤理化性质是影响土壤真菌群落结构的重要因素[37]。本试验结果显示土壤真菌类群(属水平)与土壤氮、磷、钾元素的质量分数及容重、含水率等理化性质存在相关关系,说明土壤理化性质显著影响真菌群落的相对丰度。部分真菌菌属耐受性较高,受环境因子影响较小,如大型真菌湿伞属(Hygrocybe)和被孢霉属(Mortierella)等。有效磷与速效钾作为土壤有效养分可反应土壤肥力状况,Myrmecridium与土壤中有效磷质量分数和速效钾质量分数呈显著正相关,杉木人工林在毛竹入侵后土壤肥力下降与Myrmecridium丰度降低有关。同时,有研究显示土壤真菌群落结构的变化与毛竹入侵过程中无机氮浓度的变化有关[38]。本文中随着毛竹入侵杉木林,表层土壤中铵态氮质量分数显著上升,硝态氮质量分数显著下降,表明毛竹的氨化作用增强,硝化作用减弱[39]。毛竹纯林优势真菌群落与土壤铵态氮质量分数呈显著正相关,沙蜥属(Saitozyma)、木霉属(Trichoderma)和小菇属(Mycena)的相对丰度也与铵态氮质量分数呈相同的变化趋势,表明此类菌属参与了无机氮循环过程。土壤真菌多属好氧型菌,土壤容重、含水率及孔隙度会影响土壤真菌的呼吸作用[40],适宜的土壤含水率将更利于土壤真菌的生存。毛竹纯林土壤含水率显著下降,且非毛管孔隙度高于其他林分类型。沙蜥属(Saitozyma)、木霉属(Trichoderma)与土壤含水率、毛管孔隙度呈显著负相关,与非毛管孔隙度呈显著正相关,其相对丰度在毛竹纯林中最高。由于真菌对不同环境的适应性不同,毛竹入侵后表层土壤中的优势菌群发生了改变。综上,可以认为土壤真菌多样性和群落结构与多种环境因子相关。因此,可以通过不同环境因子对真菌群落的影响来评价毛竹入侵带来的一系列影响。同时,土壤真菌如何与局部环境互相影响有待进一步研究。
5 结论
子囊菌门(Ascomycota)和担子菌门(Basidiomycota)的相对丰度在毛竹入侵后变化较大,被孢霉门(Mortierellomycota)相对丰度变化较小。圆孢霉属(Staphylotrichum)、木霉属(Trichoderma)和小菇属(Mycena)的相对丰度均随着毛竹入侵显著增加。杉木人工林中粗糙孔菌属(Trechispora)和Myrmecridium的相对丰度显著高于有毛竹入侵的林分,毛竹纯林中沙蜥属(Saitozyma)相对丰度显著高于杉木人工林和杉竹混交林。
毛竹入侵后形成的杉竹混交林土壤真菌Shannon指数、Simpson指数和PD指数最高;随着入侵程度加深,Chao1指数和ACE指数均有所升高。
环境因子能够影响土壤中真菌的相对丰度,青霉属(Penicillium)与土壤pH呈极显著正相关(p<0.01),木霉属(Trichoderma)和小菇属(Mycena)与土壤中铵态氮质量分数呈极显著正相关(p<0.01),沙蜥属(Saitozyma)、木霉属(Trichoderma)与土壤自然含水率和毛管孔隙度呈极显著负相关(p<0.01)。可见,毛竹入侵杉木人工林从一定程度上改变了表层土壤真菌多样性和群落结构。
