武夷山中亚热带常绿阔叶林群丛分类及群落构建1)
2023-02-22陈浩赵荣解雷方炎明丁晖陈水飞郑笑
陈浩 赵荣 解雷 方炎明 丁晖 陈水飞 郑笑
(南方现代林业协同创新中心(南京林业大学),南京,210037)(生态环境部南京环境科学研究所)
常绿阔叶林是常见的植被类型之一[1]。中国具有的常绿阔叶林是世界上分布最广、最典型的森林类型[2]。选择有代表性的地段进行群落生物多样性研究,对了解常绿阔叶林的生物多样性形成和维持机制有着重要意义,也可为该地区生物多样性保护提供科学依据[3-5]。近年来,大型森林动态样地已经从建立之初以群落生态学研究为主,发展为多学科交叉的生物多样性科学综合研究平台[6-8]。
植被分类是植物生态学研究中复杂的问题之一[9],由于植被分布的复杂性和主要植物学学派研究对象的地域局限性,至今尚未形成一个能被全球植被生态学领域普遍接受的植被分类系统[10-11]。以往研究植物群落数量分类的主要方法是双向指示种分析法(TWINSPAN)[12],其在生态学领域得到了广泛应用,如Bolliger[13]在威斯康星州研究景观生态学。近期研究表明,多元回归树更具优势[14],尤其是在划分森林动态监测样地连续样方或具有过渡性质样方时,也成为研究植被群落物种分布与环境因子之间关系的数量分类方法。该方法是结合环境因子作为分类节点,利用递归划分法,将样方尽可能分为同质的类群[15-16],避免了人为因素的干扰,分类结果更为客观,能够揭示植被空间分布格局[17]。
群落构建研究对于分析物种共存和物种多样性的维持是至关重要的,因此一直是生态学研究的中心论题,但同时也是发展中的论题[18]。在生态学领域提出了各种各样的假说阐述物种多样性维持机制,其中目前最具代表性的是生态位理论和Hubbell提出的群落中性理论[19-20]。生态位理论主要强调物种的相互作用,即选择过程;中性理论则不考虑选择过程,只考虑漂变、成种、扩散过程,虽然对许多事实无法解释,但是其合理的部分可以很好地弥补生态位理论的不足[21]。已有研究对浙江古田山亚热带常绿阔叶林的研究表明,群落物种的丰富度和组成主要是由扩散限制决定的,而生境特化是局域尺度木本植物多样性维持主要的机制[22-23]。越来越多的研究者认为,群落构建的机制为两者的整合[24-25],在对浙江鼎湖山的研究表明,二者联合作用对其样地物种多样性的贡献率高于二者的单独贡献率[26];但对这些理论的实践研究仍然较少。为此,本研究在武夷山市星村镇四新村附近区域,以武夷山常绿阔叶林9.6 hm2森林动态监测样地为试验地,以2018年对森林动态监测样地复查的物种分布及地形因子数据为基础数据;将物种分布与地形因子结合,使用多元回归树(MRT)将样地内群落划分,应用非度量多维标度(NMDS)、相似分析(Anosim)确认多元回归树群丛划分准确性,应用相似性百分比分析群丛之间差异性和群落构建机制;综合研究结果对武夷山中亚热带常绿阔叶林群丛进行划分。旨在为中亚热带森林群落群丛的科学划分、群落构建机制的研究提供参考。
1 研究区概况
研究区位于武夷山市星村镇四新村附近区域(地理中心坐标27°35′24″N、117°45′55″E)。该区属于典型的亚热带季风气候区,年均温为19.2 ℃,相对湿度为78%~84%,年降水量为1 486~2 100 mm,年无霜期达253~272 d。土壤类型呈明显的垂直分布特征,其土壤类型主要分为山地草甸土、黄红壤、黄壤、红壤4种类型;海拔800 m以下地区为红壤,pH为4.5~5.0,呈强酸性,为常绿阔叶林带发育的地带性土壤;海拔800~1 000 m为黄红壤,自然植被从常绿阔叶林过渡到常绿针阔混交林;黄壤带分布于海拔1 000~1 200 m,除少量灌丛外,均为常绿针阔混交林植被带;海拔高度1 200 m以上区域分布的是山地草甸土,其pH在5.0以上,坡度平缓,为5°~8°,以草甸类植被占优势[27-28]。武夷山地区最为典型的森林生态系统是亚热带常绿阔叶林,该生态系统的群落结构比较复杂,林冠层相对整齐;遍布壳斗科(Fagaceae)、樟科(Lauraceae)、木兰科(Magnoliaceae)、山茶科(Theaceae)等常绿树种,这些树种也是我国中亚热带季风气候区其他森林群落的重要组成成分。
2 研究方法
2.1 样地设置
2013年10—12月份,原环境保护部南京环境科学研究所、南京林业大学、福建省武夷山生物研究所3家单位合作,参照美国史密森热带研究所热带森林研究中心(CTFS)的调查技术规范在研究区内设置了1处9.6 hm2的动态监测样地。本研究将9.6 hm2的动态监测样地设为研究样地,应用2018年复查的物种分布及地形因子数据进行分析。样地呈长方形,长边为东北—西南方向(长400 m),西北—东南方向为短边(宽240 m);所在主山脊呈东北—西南走向,而西北—东南方向则主要由主山脊向两侧山脚延伸。样地被分成240个20 m×20 m大样方,每个大样方4个角的坐标位置均使用全站仪精确测定,并用水泥桩对这4个角进行永久标记。植物调查正式开始前,将每个20 m×20 m大样方分成16个5 m×5 m的小样方,对每个小样方内所有胸径在1 cm以上的木本植物种名、胸径、坐标、树高等信息详细记录,并挂牌标记,以便进行长期监测。首次样地调查表明,样地内共有乔木植物171种,64 973株独立个体(包括分枝和萌枝),隶属于44科86属。近年来,样地未受到太多人为干扰,常绿树种占据绝对优势,样地垂直结构复杂,分层现象也较为明显,种群天然更新状况良好。
2.2 群丛划分及命名方法
为了减小胸径较小的个体对群丛划分的影响,本研究使用样地内170种木本植物在240个样方重要值作为评价指标,重要值(VI)的计算方法:VI=(相对多度+相对优势度)/2。
以20 m×20 m大样方的海拔、凹凸度、坡度、坡向作为影响因素,构建多元回归树。地形因子参考美国史密森热带研究所热带森林研究中心的调查技术规范常用算法[29-30]。
本研究参考宋永昌[4]关于群丛的定义和中国常绿阔叶林群丛的命名方法,尽可能地展现该群丛的优势种和指示种,使得群落的层片结构得到充分反映。优势种,即群丛中重要值最高的2个个体,2者用“+”相连;指示种,群丛中具有标志性的物种,通常采取指示值较高的2物种,2者用“+”相连。确认优势种与指示种后,指示种在前、优势种在后,2者用“-”相连。
2.3 群丛相似性的判别方法
本研究使用非度量多维标度(NMDS)、相似性分析,比较与评估划分后群丛的组成特征和差异;根据Bray-Curtis距离的物种矩阵,表征不同群丛物种在空间上的分布情况;依据999次置换后的相似性分析,判断群丛间与群丛内差异性。
2.4 群落构建方法
置换相似性百分比分析是一种依据物种的有无,推断群落构建过程的方法。通过构建3种零模型(生态位-扩散限制模型、生态位限制模型、扩散限制模型);然后,对于每个零模型进行相似性百分比分析,计算零相似性百分比与经验相似性百分比之间的平方偏差之和的对数(E值),依据E值判断群落构建受到哪个过程的影响。
2.5 数据处理
本研究所有的数据处理都是在R4.1.1中进行的。应用plantlist程序包,并且结合《中国植物志》电子版查询和处理植物学名;应用R包vegan进行非度量多维标度、相似性分析;应用R包labdsv进行指示值计算;应用Gibert et al.[31]描述的方法,评估确定性与随机性过程在武夷山常绿阔叶林群落结构构建中的重要性。
3 结果与分析
应用多元回归树分析表明,依据海拔因素将样地划分为4个群丛(见图1),分别是群丛A(编号为0101~0103等3个样方,其海拔高于567.7 m)、群丛B(编号为0104~1005等110个样方,位于海拔515.0~567.7 m)、群丛C(编号为1006~1809等100个样方,位于海拔479.7~515.0 m)、群丛D(编号为1810~2012等27个样方,位于海拔低于479.7 m),4个群丛分布(见图2),其具有很明显的海拔分布差异。
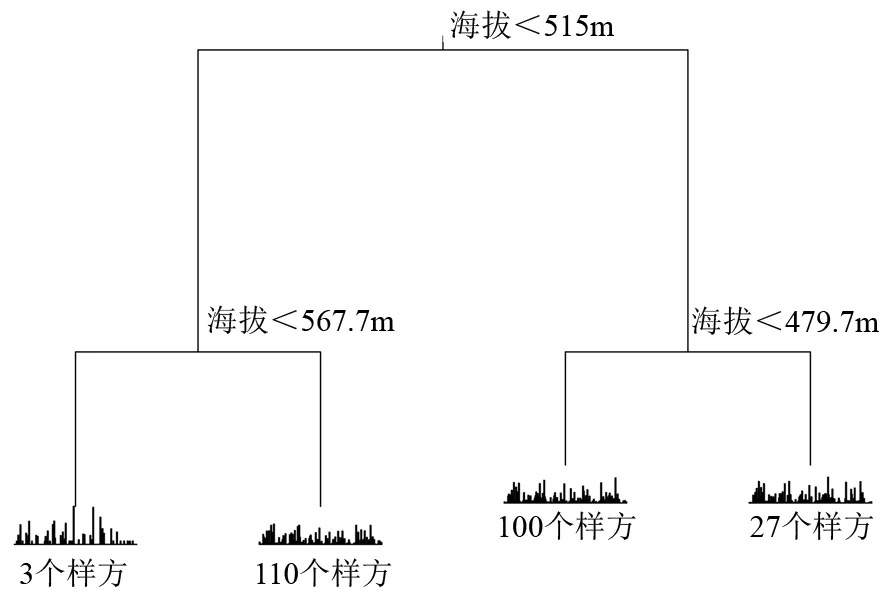
误差为0.915,交叉验证误差为0.964,标准误差为0.019 8。
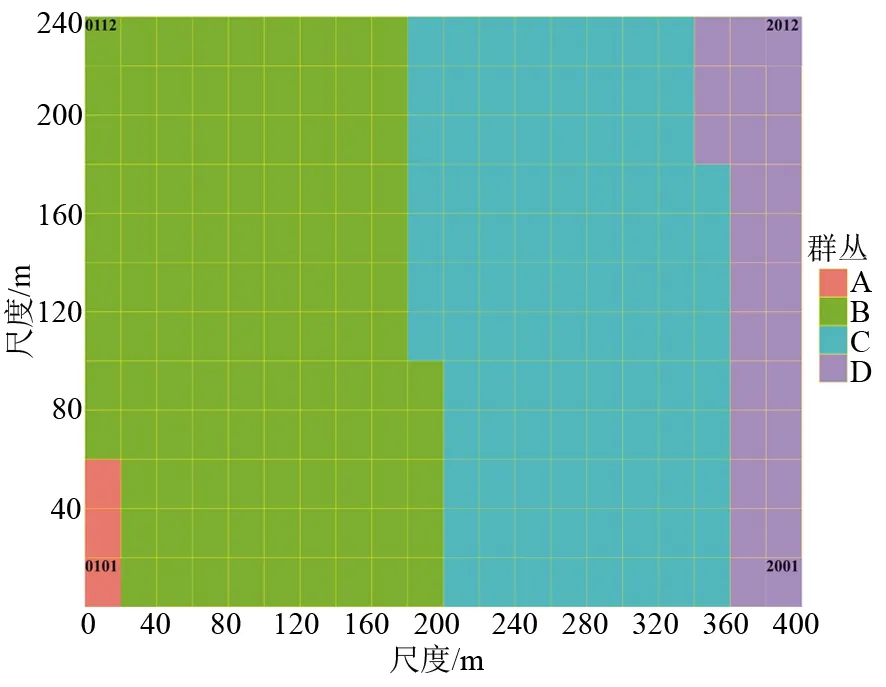
图2 武夷山9.6 hm2常绿阔叶林森林动态监测样地20m×20m的4类群丛划分
4个群丛的指示值各不相同(见表1),群丛A中红毒茴(Illiciumlanceolatum)、山桐子(Idesiapolycarpa)、椤木石楠(Photiniadavidsoniae)的指示值相对较高,分别为0.970 0、0.883 7、0.775 2;群丛B和群丛C各仅有1种指示物种,分别为米槠(Castanopsiscarlesii)、赤楠(Syzygiumbuxifolium),指示值为0.376 5、0.426 5;群丛D中樟叶泡花树(Meliosmasquamulata)、凸脉冬青(Ilexkobuskiana)、窄基红褐柃(Euryarubiginosavar.attenuata)的指示值相对较高,分别为0.606 1、0.570 6、0.492 0。4个群丛的重要值见表2。
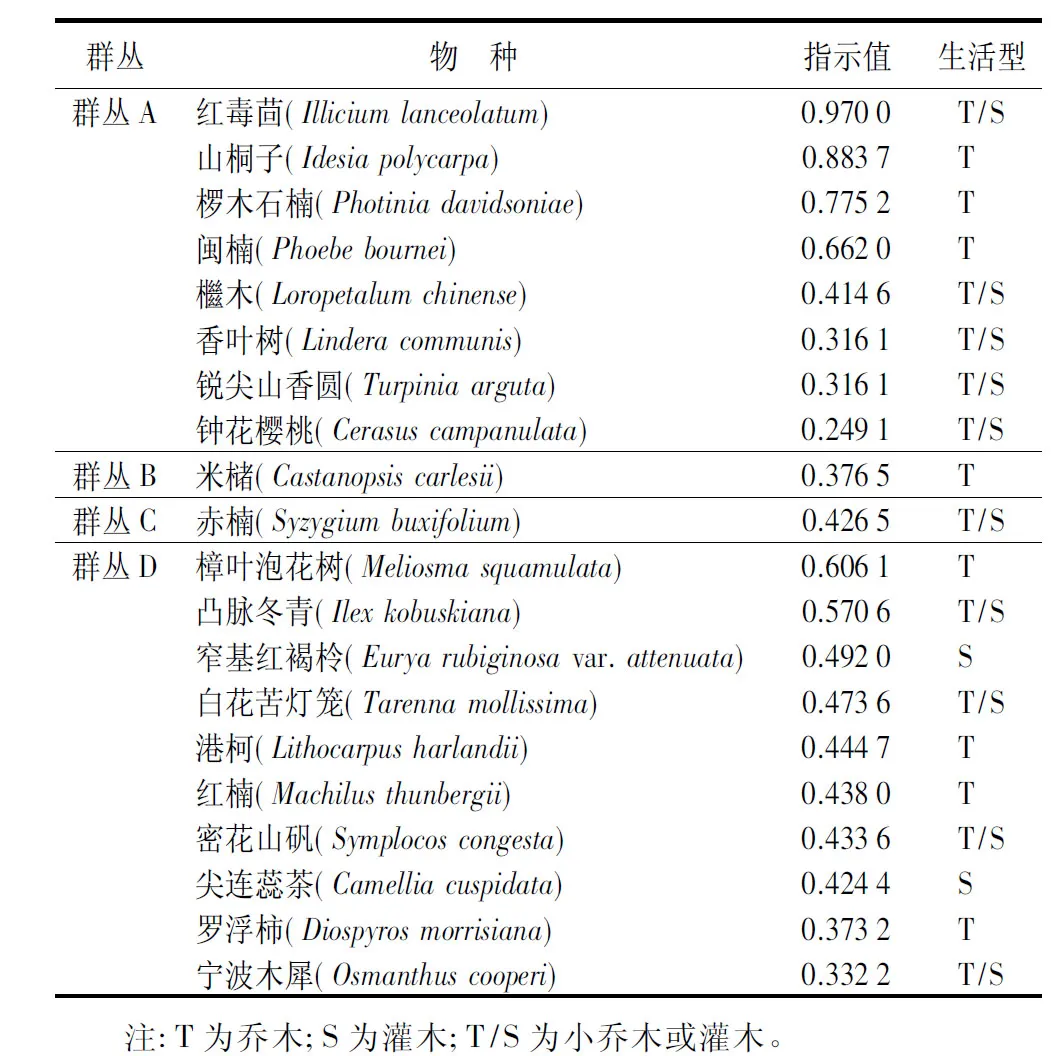
表1 武夷山9.6 hm2森林动态监测样地4类群丛内物种指示值排序

表2 武夷山9.6 hm2森林动态监测样地4类群丛内物种重要值排序
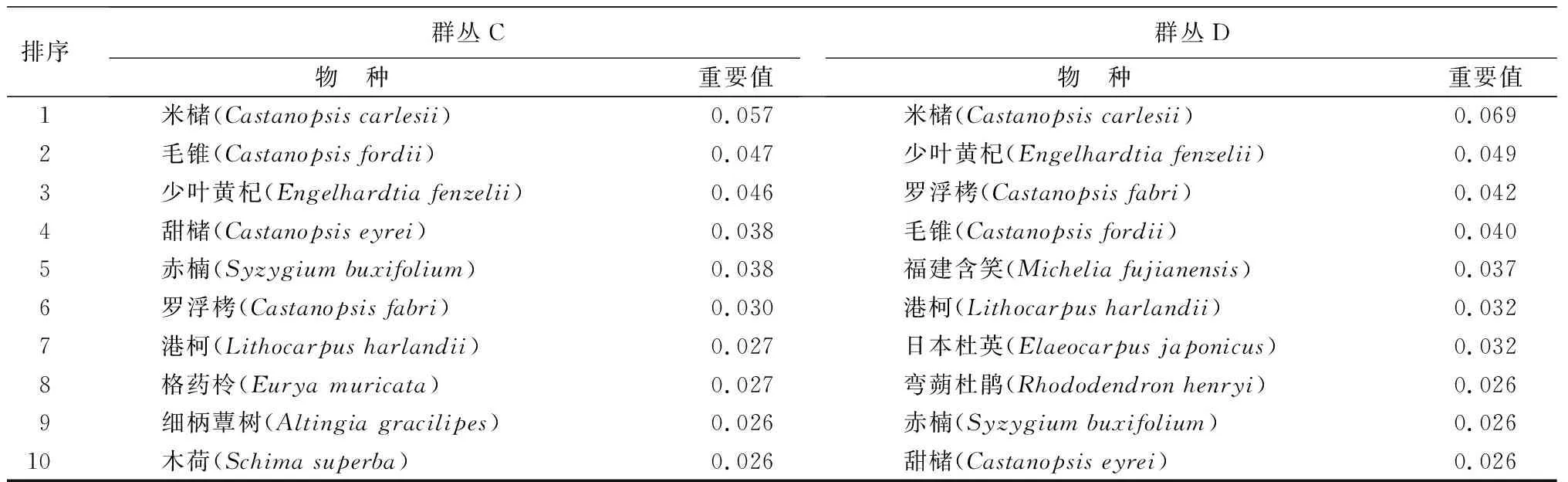
排序群丛C物 种 重要值群丛D物 种 重要值1米槠(Castanopsis carlesii)0.057米槠(Castanopsis carlesii)0.0692毛锥(Castanopsis fordii)0.047少叶黄杞(Engelhardtia fenzelii)0.0493少叶黄杞(Engelhardtia fenzelii)0.046罗浮栲(Castanopsis fabri)0.0424甜槠(Castanopsis eyrei)0.038毛锥(Castanopsis fordii)0.0405赤楠(Syzygium buxifolium)0.038福建含笑(Michelia fujianensis)0.0376罗浮栲(Castanopsis fabri)0.030港柯(Lithocarpus harlandii)0.0327港柯(Lithocarpus harlandii)0.027日本杜英(Elaeocarpus japonicus)0.0328格药柃(Eurya muricata)0.027弯蒴杜鹃(Rhododendron henryi)0.0269细柄蕈树(Altingia gracilipes)0.026赤楠(Syzygium buxifolium)0.02610木荷(Schima superba)0.026甜槠(Castanopsis eyrei)0.026
结合重要值与指示值,群丛命名如下:
群丛A——“红毒茴+檵木-闽楠+福建含笑”群丛(Illiciumlanceolatum+Loropetalumchinense-Phoebebournei+Micheliafujianensisassociations)。
群丛B——“米槠-毛锥+甜槠”群丛(Castanopsiscarlesii-Castanopsisfordii+Castanopsiseyreiassociations)。
群丛C——“赤楠-米槠+毛锥”群丛(Syzygiumbuxifolium-Castanopsiscarlesii+Castanopsisfordiiassociations)。
群丛D——“樟叶泡花树+凸脉冬青-米槠+少叶黄杞”群丛(Meliosmasquamulata+Ilexkobuskiana-Castanopsiscarlesii+Engelhardtiafenzeliiassociations)。
非度量多维标度分析进一步表明,群丛A与群丛B、群丛C、群丛D所在样方的物种丰度差距较大(见图3)。群丛B中所在样地的数量较多,群丛D的分布与群丛B、群丛C有较大的相似性。结合指示值和重要值,群丛A的样方数量虽然较少,但是其中的物种部分较为特殊,例如香叶树(Linderacommunis)、锐尖山香圆(Turpiniaarguta)、钟花樱桃(Cerasuscampanulata)等,这些物种在整个大样地中的数量较少,大部分都集中在群丛A中,即样方0101~0103中;一方面是由于群丛A的海拔是样地中最高的,另一方面是由于这些样方位于样地边缘,存在较强的异质性。决定系数(R2)为0.04,相似性分析表明,群丛内差异小于群丛间差异(P<0.05)。

图3 依据4种群丛物种丰度的非度量多维标度排序图
由图4可见:群丛B的特有种最多,有8个特有种,分别为大萼杨桐(Adinandraglischrolomavar.macrosepala)、乌药(Linderaaggregata)、青灰叶下珠(Phyllanthusglaucus)、海金子(Pittosporumillicioides)、微毛山矾(Symplocoswikstroemiifolia)、木蜡树(Toxicodendronsylvestre)、棕榈(Trachycarpusfortunei)、山黄麻(Trematomentosa)。群丛C的特有种有6个,分别为油茶(Camelliaoleifera)、鹿角锥(Castanopsislamontii)、大花枇杷(Eriobotryacavaleriei)、异叶榕(Ficusheteromorpha)、毛竹(Phyllostachysedulis)、满江红(Azollapinnatasubsp.asiatica)。群丛B和群丛C的物种相似性较高,处于中海拔地区的样方。群丛A和群丛D没有特有种,由于海拔差距的原因,两者的共有种也很少。
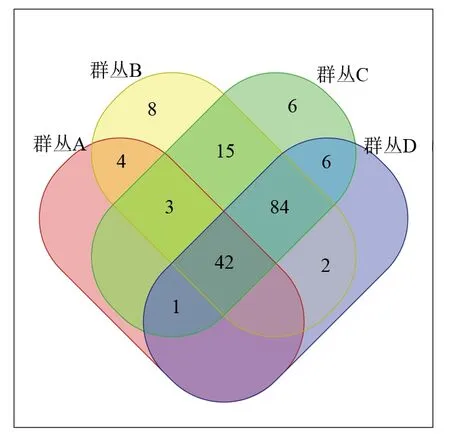
4种颜色图形交叉中的数据为交叉群丛中共有物种数,图形非交叉中的数据为该群丛的特有物种数。
4 结论与讨论
大量的研究表明,地形与环境异质性具有关联性[32],说明地形在生态学研究中是1个重要的指标。近年来,在对常绿阔叶林群丛分类的研究中,结合地形因子已经成为热门的方法。不同的地形存在着不同的生境,不同的植物对其有各自的适应性。本研究认为,影响武夷山9.6 hm2动态监测样地的主要地形因子为海拔,多元回归树的每个分支节点均有其影响,虽然该样地上坡位和下坡位的绝对高度不是很大,但海拔反映出的是水分和温度的影响,同时也是影响植物空间分布格局的一个重要因素。在该样地内划分的4个群丛中,群丛A位于海拔较高的地区,且旁边溪流,此独特的生境生长着与其他3个群丛截然不同的物种。群丛B和群丛C的地形主要以山坡为主,生长的物种多为样地的优势种。群丛D位于海拔较低的位置,且因为有数条较深的山沟,其物种与群丛B、群丛C相似,但有一定的差异。群丛的指示种代表着1个物种在该群丛内的密度比其他群丛高,且在该群丛内分布较为均匀。群丛A和群丛D的指示种数量较多,且指示值较大,说明在这2个群丛内的一些物种,如群丛A中的红毒茴、山桐子、椤木石楠等对该群丛具有显著的指示作用,在群丛内的分布也较为均匀;但是这些物种对其他群丛没有显著的指示意义,分布也较少。通常指示种数量较多的群丛,其指示值也较大[2],上述2个群丛符合这一特性;原因是由于这2个群丛具有较为极端的环境,而具有较窄生态幅的物种在这些特异的生境占据着优势。相似性分析表明,划分的4个群丛具有差异,说明多元回归树适用于植物群丛划分。
已有研究中,群落构建的机制一直存在着很多不同的观点和理论[33];越来越多的研究者认为,其最终将是生态位理论和中性理论的整合[34]。本研究结果表明,依据生态位理论的环境过滤与竞争排斥、依据中性理论的扩散限制,均对武夷山群落构建起到促进作用。在局域尺度上,无论是依据气候还是依据地理的物种库,都不能完全解释当地群落的分类结构;所以环境过滤和扩散限制,通过物种分类共同调节当地群落的结构。本研究对武夷山9.6 hm2森林动态监测样地的群落构建机制进行初步探究,但考虑的因素不够全面,多元回归树使用的地形因子仅有4个;在今后的研究中,如能加入更多因子,如土壤含水量、土壤pH等,研究结果会更加深入。依据植物功能性状研究群落的构建机制也很重要,它能通过性状分布模式的方式确定环境过滤和生态位分化两者的重要性[34],本研究结果可为今后大样地更深入研究提供参考。
