2株猪繁殖与呼吸综合征病毒海南株的ORF5基因分析
2023-02-21马佳镁钟植文范悦轩黄春媛郑佳馨曹芳芳王金泉刘光亮曹宗喜
马佳镁,钟植文,范悦轩,黄春媛,郑佳馨,曹芳芳,王金泉,刘光亮,4,曹宗喜
(1.新疆农业大学动物医学学院,乌鲁木齐 830052;2.海南省农业科学院畜牧兽医研究所/海南省热带动物繁育与疫病研究重点实验室/国家动物疫病数据中心海口观测实验点,海口 571100; 3.广东永顺生物制药股份有限公司,广州 511536;4.海南省农业科学院畜牧兽医研究所海南省院士团队创新中心,海口 571100)
0 引 言
【研究意义】 猪繁殖与呼吸综合征(Porcine reproductive and respiratory syndrome,PRRS)是引起各年龄段猪的不同程度的呼吸道病症及妊娠母猪的繁殖障碍等临床特征的高死亡率疾病。而随着重组毒株的流行,使得PRRS更难以防控,市场上的疫苗仍不能完全控制PRRSV 的入侵。采集海南省PRRSV临床样本选取典型的毒株通过分析ORF5判断市场流行毒株,分析海南流行毒株的特性,对PRRSV的防控和疫苗的选择有重要意义。【前人研究进展】PRRS自20世纪80年代末在欧洲和美国出现[1],1995年中国报道出现了猪繁殖与呼吸综合征[2],由于PRRSV在宿主体内作用时不断的进化,因此PRRSV变异株与以前的分离株相比有着更强的毒力,2006年高致病性毒株的暴发以及2012年NADC30样毒株流行[3]。尽管PRRSV基因型不同,但临床表现一致[4]。PRRSV属套式病毒目,动脉炎病毒科,与马动脉炎病毒(Equine arteritis virus, EVA)、鼠乳酸脱氢酶升高症病毒(Lactate dehydrogenase elevating virus,LDV)和猴出血热病毒(Simian hemmorrhagic fever vieus,SHFV)同属动脉炎病毒属[5]。 PRRSV是一种有囊膜的单股正链RNA病毒[6]。基因组大小约为15.4 kb,5’和3’端分别有非编码区,包含至少10个开放阅读框(ORF)[7]。ORF5编码的GP5作为PRRSV的主要结构蛋白,不仅参与细胞免疫和体液免疫,也是病毒复制必不可少的因素之一[8],是分析PRRSV遗传变异和分子流行病学的重要指标。SHI MANG等[9]对8 624个PRRSVORF5的样品进行序列对比,利用MrBayes v3.2中的BMCMC的方法将北美型PRRSV划分9个谱系,构建了PRRSV较为完整的分类系统。根据分类系统, 将PRRSV-Ⅰ分为4个亚型(1-4亚型),将PRRSV-Ⅱ分为9个亚型。由于遗传多样性的不同,毒力和抗原性也出现较大的差异。目前我国PRRSV流行毒株仍以北美型为主,我国流行的PRRSV主要可分为4个谱系:1、3、5(5.1 亚系)和 8(8.7 亚系)。Liu等[10]将分离的病毒命名为FJLIUY-2017,与 GenBank中24株PRRSV的序列比对,使用SimPlot v3.5.1和RDP 4.80软件进行重组分析,发现毒株为谱系1、3、5、8重组毒。我国PRRSV的流行情况较为复杂,出现不同谱系病毒重组现象。【本研究切入点】目前,猪繁殖与呼吸综合征仍被认为是猪最重要的病毒性疾病。我国海南在2007年有PRRSV相关报道,而近年尚未有数据。需研究海南新分离2株PRRSV毒株流行情况。【拟解决的关键问题】采用PCR方法对ORF5基因进行扩增和序列测定,利用MegAlign软件进行多重序列比对,利用MEGA X软件邻接法构建进化树,通过PRRS分子流行病学分析,为我国海南PRSSV的有效防控打下基础。
1 材料与方法
1.1 材 料
PRRSV HNHK1-2021 株和PRRSV HNLG1-2021株从我国海南不同猪场分离而来,由海南省热带动物繁育与疫病研究重点实验室保存;RNAiso Plus(9 108Q)、dNTP Mixture(4 030Q)、Recombinant RNase Inhibitor(2 313Q)、Reverse Transcriptase M-MLV(2 641Q)、DL 2 000 DNA Marker(3 427A)、E.coliDH5α Competent Cells(9 057)、pMD18-T Vector(6 011)均购自宝生物工程(大连)有限公司;DNA纯化试剂盒(DC301-01)、质粒快速提取试剂盒(DC201-01)均购自南京诺唯赞公司。
1.2 方 法
1.2.1 病毒RNA提取及ORF5基因RT-PCR扩增
采用RNAiso法对PRRSV HNHK1-2021株和HNLG1-2021株提取RNA。反转录体系:5×M-MLV缓冲液4 μL,dNTP 4 μL,RRI 0.5 μL,反转录引物1 μL,RNA提取液10 μL。42°水浴,1 h。PCR反应体系:灭菌蒸馏水7 μL,2×ExTaq10 μL,上、下游引物各1 μL,cDNA 2 μL。PCR反应条件:94℃ 3 min;94℃ 30 s,50℃ 1 min,72℃ 50 s,共 35 个循环;72℃ 7 min。表1
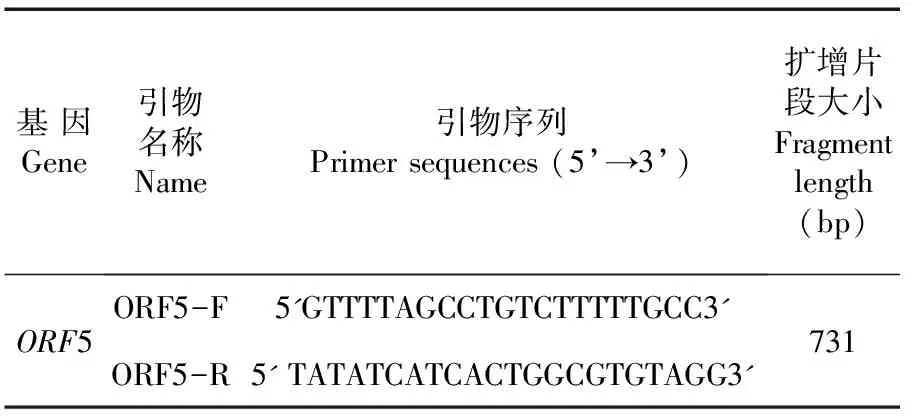
表1 引物序列信息
1.2.2 目的基因的克隆与序列测定
PCR扩增的目的片段进行琼脂糖凝胶电泳,用DNA凝胶回收试剂盒回收目的片段,胶回收产物连接pMD18-T Vector,16℃,30 min。连接产物转化至DH5α感受态细胞,用Amp抗性固体培养基温箱37℃,16 h培养。挑取单个菌落进行菌液PCR验证,阳性菌液送生工生物工程(上海)有限公司测序。
1.2.3ORF5基因的遗传多样性和系统发育
利用GenBank中代表性的毒株,主要为北美型PRRSV,针对我国流行毒株选择NADC30类PRRSV、QYYZ类 PRRSV、VR-2332 类 PRRSV、JXA1类PRRSV的ORF5片段,对 HNHK1-2021株和HNLG1-2021株的ORF5基因进行分析。采用MegAlign软件进行多重序列比对,利用 MEGA X软件1000 bootstrap值邻接法构建进化树。表2
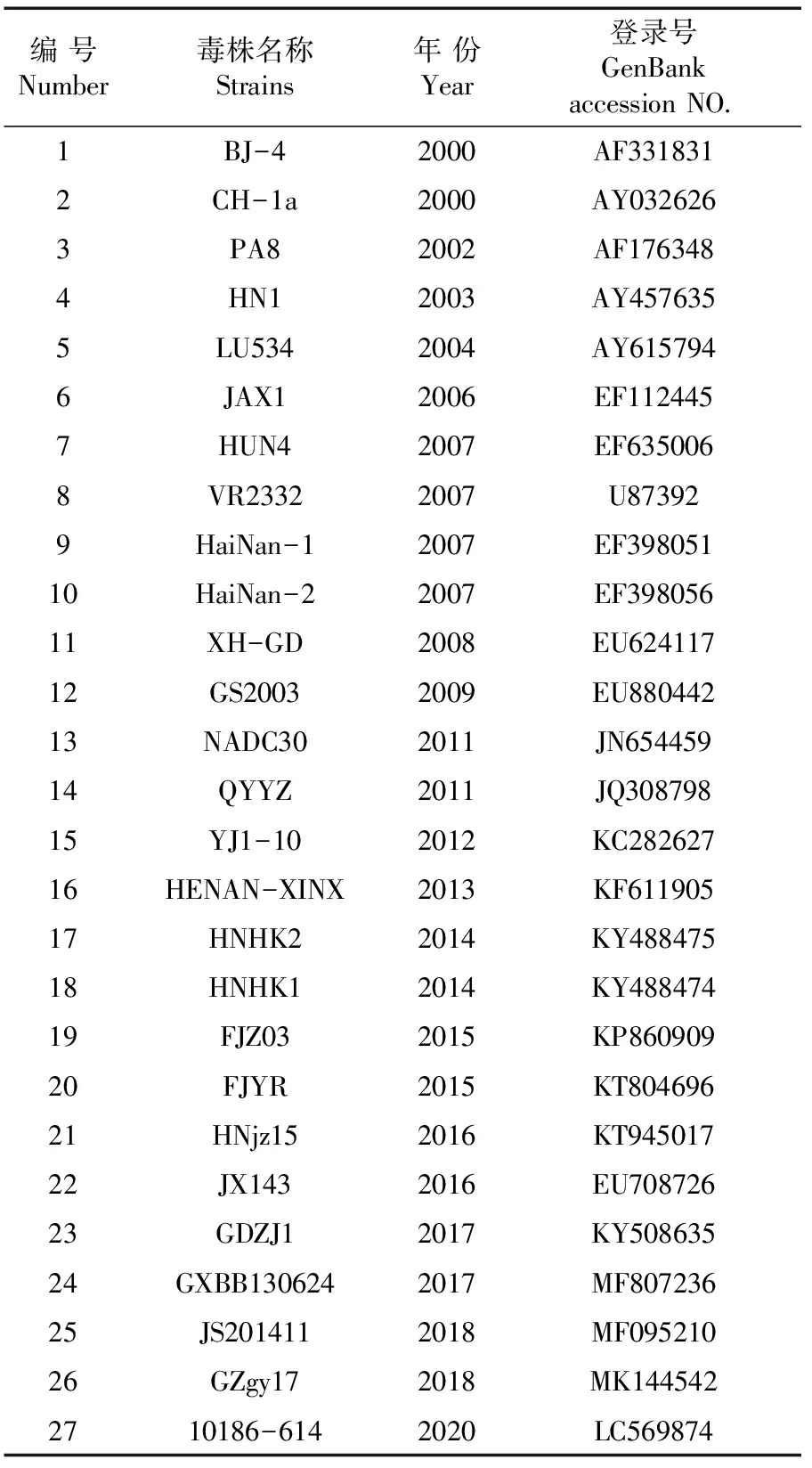
表2 PRRSV代表毒株信息
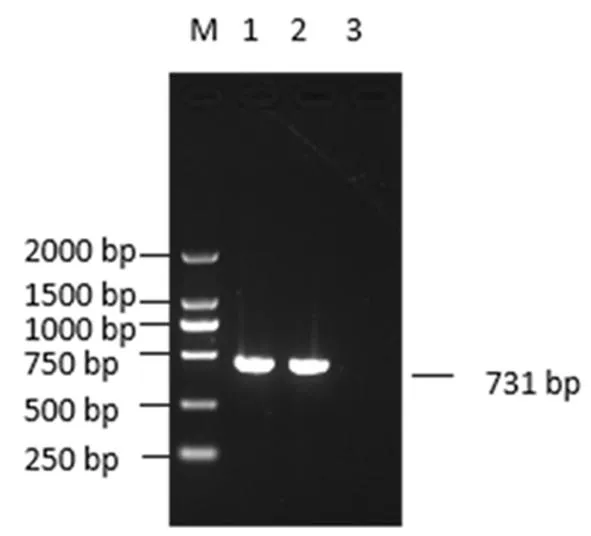
注:M:DL-2 000 DNA Marker;1:HNHK1-2021毒株ORF5;2.HNLG1-2021 毒株ORF5;3.阴性对照
2 结果与分析
2.1 目的片段的扩增
研究表明,利用ORF5-R和ORF5-F引物扩增目的片段,PCR产物用1%琼脂糖凝胶电泳鉴定得到两株PRRSVORF5条带大小约为603 bp,包含5’端ORF4基因和3’端ORF6基因的部分片段(128 bp),总扩增大小731 bp,与预期大小相符。图1
2.2ORF5基因的遗传多样性
研究表明,采用MegAlign软件对PRRSV HNHK1-2021和HNLG1-2021的ORF5序列比对,两株毒之间核苷酸相似性为82.9%,氨基酸相似性分别为82.6%。2毒株同源性差异较大,不属于同一谱系。PRRSV HNHK1-2021株与海南PRRSV株ORF5核苷酸相似性为81.5%~82.3%,氨基酸相似性为83.5%~84.1%。PRRSV HNLG1-2021株与海南PRRSV株ORF5核苷酸相似性为86.8%~86.9%,氨基酸相似性为88.0%~88.1%,新发现的2株毒和海南之前的毒株差异较大。表3
PRRSV HNHK1-2021和其他27株毒的ORF5序列比对,核苷酸相似性为80.8% ~ 92.0%,氨基酸相似性为80.1% ~ 94.5%。HNHK1-2021毒株和NADC30类PRRSV序列比对,核苷酸相似性分别为80.8% ~ 82.8%,氨基酸相似性分别为82.1% ~ 84.6%;和QYYZ类PRRSV序列比对,核苷酸相似性分别为90.2% ~ 92.0%,氨基酸相似性分别为91.5% ~ 94.5%;和VR-2332 类 PRRSV序列比对,核苷酸相似性分别为82.1% ~ 83.4%,氨基酸相似性分别为80.1% ~ 83.1%;和JXA1类 PRRSV序列比对,核苷酸相似性分别为81.9% ~ 84.4%,氨基酸相似性分别为82.6% ~ 84.6%。其中,与YJ1-10毒株核苷酸相似性最高92.0%,与谱系3代表毒株QYYZ的同源性为90.2%。和QYYZ毒株氨基酸序列比对发现有信号区Q13R突变,中和表位(PNE)与QYYZ株完全一致,跨膜区TM2和TM3中H102Y、F117L、I124V突变。
核苷酸相似性为81.8% ~ 99.5%,氨基酸相似性为81.1% ~ 98.5%。HNLG1-2021毒株和NADC30类PRRSV序列比对,核苷酸相似性分别为83.7% ~ 84.6%,氨基酸相似性分别为83.6% ~ 85.1%;和QYYZ类PRRSV序列比对,
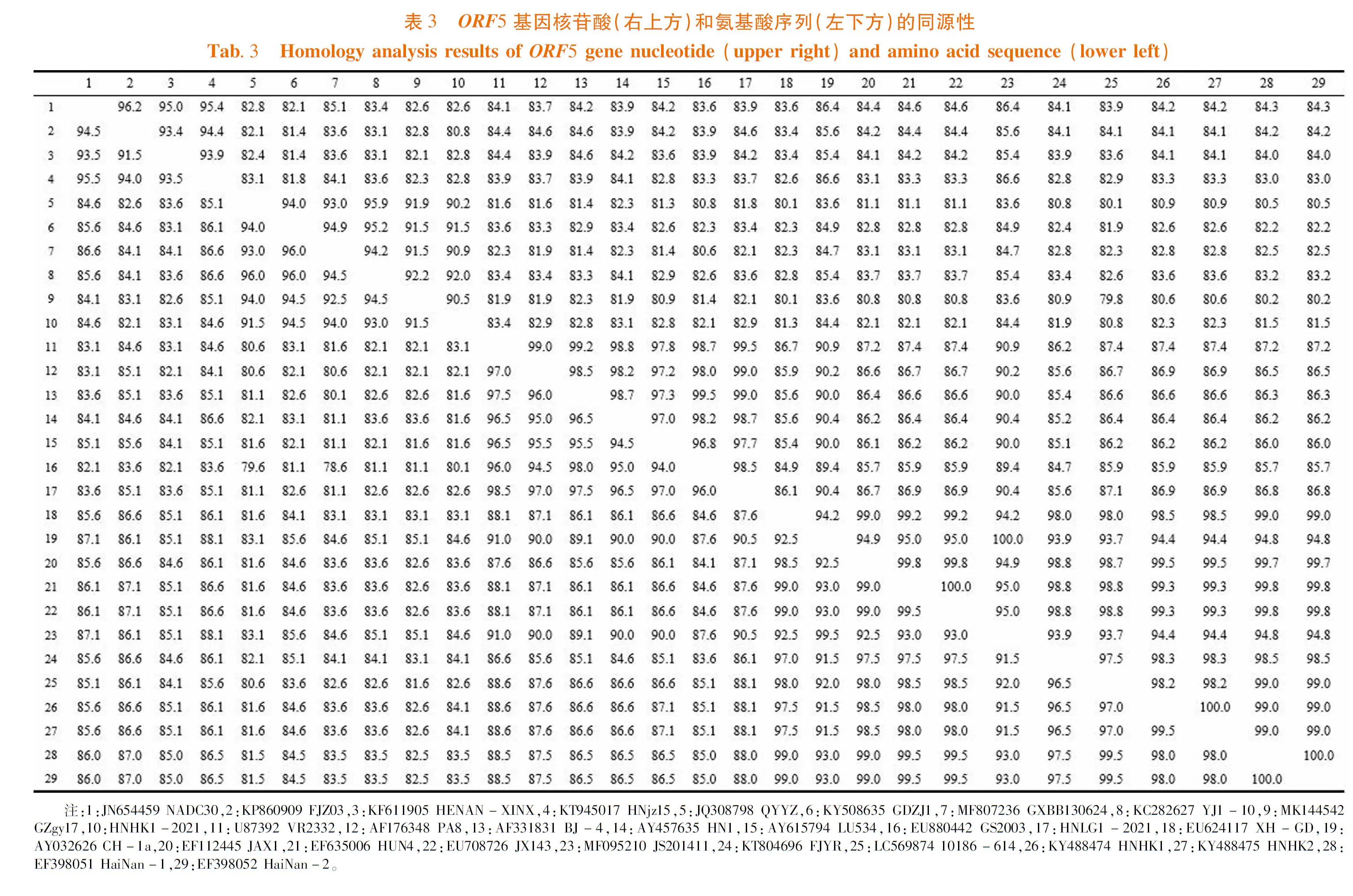
核苷酸相似性分别为81.8% ~ 83.6%,氨基酸相似性分别为81.1% ~ 82.6%;和VR-2332 类 PRRSV序列比对,核苷酸相似性分别为97.7% ~ 99.5%,氨基酸相似性分别为96.0% ~ 98.5%;和JXA1类 PRRSV序列比对,核苷酸相似性分别为85.6%~90.4%,氨基酸相似性分别为86.1% ~ 90.5%。其中,与谱系5代表毒株VR-2332的核苷酸同源性高达99.5%。和VR-2332毒株序列相比尽管N30、N101位点突变,但不引起氨基酸改变。图2


图2 PRRSV ORF5氨基酸序列比对Fig.2 PRRSV ORF5 amino acid sequence alignment
2.3 进化关系
研究表明,GenBank上传的中国海南PRRSV株均属于谱系8(JXA1类PRRSV)。PRRSV HNHK1-2021株和HNLG1-2021株均属于北美型。HNHK1-2021毒株和GZgy17毒株最为接近且同属一个分支,属于谱系3(QYYZ类 PRRSV); HNLG1-2021毒株和VR-2332毒株、HN1毒株最为接近,属于谱系5(VR-2332类PRRSV)。图3

注: 为本次分离得到的毒株, 为海南不同时期分离得到的毒株
3 讨 论
3.1GP5 蛋白作为PRRSV主要囊膜蛋白和主要的免疫原性蛋白,对PRRSV感染有着重要作用,可诱导机体产生特异性的中和抗体[11]。在体外可以诱导易感的Marc-145细胞和PAM细胞产生凋亡,在体内也可诱导肺和淋巴组织的细胞产生凋亡[12]。由于GP5高变异性,因此利用GP5的差异性来研究遗传变异性[13]。根据ORF5基因分型,PRRSV主要被分为欧洲型和北美型。北美型PRRSV分为9个不同的遗传谱系(谱系1-9)。谱系1进一步划分为9个亚系[14]。中国PRRSV最常见三大类病毒NADC30样毒株、高致病性毒株、经典株主要分布在谱系1、谱系3、谱系5、谱系8[15]。
PRRS病原体由分离得到,分离的菌株分别命名为CH-1a和BJ-4[16],但与经典株相比,HP-PRRSV样株和NADC30样株显示出较高的遗传变异和重组发生率。文献[17]发现抗HUN4-F112的高免疫血清不能中和谱系3的毒株,可能是由于GP5的高度变异引起的。
3.2基因组序列分析表明,PRRSV HNHK1-2021株与QYYZ、YJ1-10、GZgy17等谱系3的毒株的ORF5序列同源性均高于90.0%,与其他谱系毒株的ORF5序列同源性均在85%左右,与CH-1a经典株的ORF5同源性为86.1%高于NADC30样毒株和高致病性毒株。HNLG1-2021株与VR-2332株、PA8株、BJ-4株、HNⅠ株、LU534株、GS2003PA8株的ORF5序列同源性均高于97.5%,同谱系1、谱系3、谱系8的毒株ORF5相比较,HNLG1-2021株与谱系8毒株的ORF5核苷酸同源性更高,与CH-1a核苷酸同源性为91.5%。系统发育树分析表明,HNHK1-2021与QYYZ株、GDZJ1株、GXBB130624株、YJ1-株、GZgy17株属同一分支,位于谱系3。HNLG1-2021株与VR-2332株属同一分支,位于谱系5。GP5 的残基R13和R151与PRRSV毒力有关[18]。氨基酸序列比对发现PRRSV HNHK1-2021株R151K突变,PRRSV HNLG1-2021株R13Q突变,这与NADC30样毒株部分吻合。AA37-44 的初级中和表位 (PNE) 在诱导免疫反应中发挥了重要作用[19],2株毒的中和表位相对保守。GP5的四个潜在N-糖基化位点的N30、N34、N44和N51与病毒感染和抗原特征有关[20],PRRSV HNHK1-2021株N34S突变。N-糖基化位点的改变可能有利于突变病毒逃脱中和[21]。
4 结 论
从我国海南分离到的2株PRRSV毒株HNHK1-2021株和HNLG1-2021株,和GenBank上传的海南株差异较大,PRRSV HNHK1-2021株属于谱系3,和QYYZ重组毒株同源性相近;PRRSV HNLG1-2021株属于谱系5,和VR-2332毒株同源性相近,PRRSV HNHK1-2021株的ORF5序列中碱基突变导致氨基酸发生改变,而PRRSV HKLG1-2021株的ORF5序列中碱基突变不影响氨基酸改变。
