外来植物入侵对喀斯特乡土物种多样性影响研究进展
2023-02-18王世雄张广奇何跃军
王世雄,张广奇,何跃军
(贵州大学 林学院,贵州 贵阳 550025)
随着全球化进程的加快,生物入侵已成为普遍现象[1-2],外来物种入侵对生物多样性、生态系统服务以及社会经济造成了严重的影响和损失[3-4],已成为21世纪全球性的生物安全问题[5]。喀斯特环境是一种脆弱的生态环境,具有生境严酷、土层薄、肥力低、环境容纳量小、抗干扰能力弱,受干扰后自然恢复的速度慢、难度大等鲜明特点[6]。尽管喀斯特生境严酷,然而,外来植物却在该区成片分布,并持续扩散。据贵州省林业科学研究院调查统计,截至2018年12月,贵州共有外来入侵物种217种。据调查,外来植物侵害区域均为典型的少数民族和贫困地区,人民生活水平低,生态保护意识淡薄,森林管理不到位,只用不管,原生植被破坏,水土大量流失,造成外来植物无生态屏障阻隔,籽实四处飘散,疯狂蔓延。研究表明,在未来5~10年内,贵州省的入侵物种将达500种以上,世界100种恶性外来入侵生物都会入侵到贵州[7]。
外来植物入侵的危害主要表现为竞争排挤和取代当地植物而很快形成单种优势群落[8],造成生物多样性不可逆转性的降低,危及当地物种的生存,甚至导致当地物种特别是珍贵植物资源的濒危或灭绝,最终导致生态系统退化,改变或破坏当地的自然景观[9-10]。随着外来入侵物种种类增多,以及原有入侵物种在全省不断扩散,天然草场面积将日益减少、畜牧业不断萎缩,耕地撂荒面积持续扩大,水利水电等基础设施的受影响程度加大,喀斯特生态系统更加脆弱。外来植物的入侵导致贵州喀斯特地区原本并不发达的农业、林业、牧业雪上加霜,再加上该区特殊的喀斯特生境,外来植物的适生区面积持续增大,遏制外来植物在喀斯特生态脆弱区的入侵迫在眉睫。
1 喀斯特生境的特点
贵州是一个喀斯特广泛分布的地区,喀斯特环境是一种易受干扰而遭破坏的脆弱生态环境,主要表现为土层薄、肥力低、水土易流失、耕地匮乏、环境容量小等多个方面。其中,土壤水分亏缺是喀斯特人工造林、植被恢复的主要障碍因子[6]。其成因不是由于降水量不足,而是因为土层浅簿、土壤总量少、贮水能力低及岩石渗漏性强,是一种湿润气候背景上的临时性干旱。同时,干旱也是外来植物入侵后初始种群建立所面临的主要环境胁迫因子[10],土壤湿度大的生境更容易受到外来植物的入侵[11]。综上所述,水分资源是喀斯特生态脆弱区限制外来植物入侵的重要环境因子,应该在这个梯度上开展相关研究。
同时,喀斯特地貌上虽然地形起伏不大,但却有丰富多样的小生境,表现为高度的生境异质性[6]。既有岩石裸露、气温变化剧烈的石芽、石面、崩塌的大块岩石等干旱生境,也有土层相对深厚、土壤营养元素丰富、有机质含量很高、气温变幅较小的石沟、石缝等肥沃生境。植物生长的关键资源如光、土壤水分和养分的有效性在这种喀斯特小空间尺度上表现了较大的变异性,这对喀斯特生境植物多样性具有一定的促进作用,但取决于群落分层[12]。外来植物要想成功入侵新生境,就必须能够应对资源供应精细尺度上的环境异质性。研究表明,无论是土壤资源的有效性还是小尺度的土壤资源的异质性,都能提高入侵植物喜旱莲子草(Alternantheraphiloxeroides)与本地同源植物莲子草的相对竞争能力[13]。一个可能解释是当外来克隆植物进入一个高度空间异质性的小尺度区域时,外来入侵植物将比乡土植物从克隆整合中获益更多,这可以部分解释外来植物在高异质性的地区(如氮沉降、施肥和干扰)成功入侵的原因[14-15]。
2 外来植物入侵对乡土物种多样性的影响过程
植物入侵会显著降低入侵生态系统内乡土植物多样性,这种影响可以通过生态系统级联、营养和能量的储存和流动以及更高营养水平的多样性而产生[16]。然而,入侵植物改变生态系统功能和营养相互作用的方式是高度可变的,可以通过入侵物种多度产生直接影响,或者通过乡土群落多样性变化产生的间接影响来实现。
2.1 入侵引起物种多度变化
外来物种与乡土物种之间往往相互抑制[17-18],相互抑制并不一定表现为竞争排斥。因此,这种效应可能仅仅表现为个体多度的变化。外来物种的入侵可以降低土壤的营养水平,加剧物种间的抑制,从而引起物种个体数量的改变。化感作用是紫茎泽兰种群竞争扩张的一个重要武器,但是,其分泌的化感物质并不是对所有物种都表现为抑制。研究表明,紫茎泽兰对一年生黑麦草(Loliummultiflorum)、多年生黑麦草(L.perenne)、紫花苜蓿(Medicagosativa)和光叶紫花苕(Viciavillosa)4种牧草无化感抑制作用,甚至存在一定的促进作用[19]。对北美入侵性杂草铺散矢车菊(Centaureadiffusa)的研究发现,铺散矢车菊在原产地亚欧大陆不造成大的影响,进入北美后却泛滥成灾,排挤了许多乡土物种。栽培试验的结果表明,在原产地与铺散矢车菊共存的植物,其根系分泌物可以抑制铺散矢车菊根系对磷的吸收并抑制其生长。然而,新栖息地的类似植物,其生长反被铺散矢车菊根系的分泌物所抑制[20]。
2.2 入侵引起种—多度格局改变
种间竞争关系是塑造植物物种空间分布格局和影响物种共存的重要机制之一。外来物种的进入,常常伴随着资源的重新分配,可以改变入侵物种与乡土物种、甚至本土物种之间的固有关系,也将必然导致乡土物种分布格局的重大改变,主要表现为物种多度格局的改变[3,4]、物种分布的范围改变[4]等。同时,物种多度格局是群落结构的重要反映,它将反映群落内不同物种个体数量的分布情况,能更好的体现出外来物种的入侵影响和种间关系变化。对巴西塞拉多生物多样性热点地区植物入侵的研究表明,入侵改变了本地物种的分布格局,促进了大多数物种的广泛传播,导致乡土物种丰富度和群落均匀度的显著增加[4]。
根据物种的多度格局追溯物种的作用机制过程是揭示群落本质的重要方式。由于物种多度的区域分布规律可能与环境资源的数量和质量有关[21],相比较传统的生物多样性指数,物种多度格局能更好的体现出物种关系和作用机制。例如,生态位优先占领模型(Niche-preemption model,NPM)认为种的个体数量与它们所占有的资源数量成比例,即优势度越高的物种应该占有的资源越多,而且NPM的群落最优势种不占绝对优势,其生态位小于总生态位的一半。随机分配模型(Random assortment model,RAM)假定一个群落中各个种的多度之间没有联系,种间竞争不存在,多数情况下物种不能全部占用其生态位。这样的群落体系的一个最大的特点是没有足够的时间提供给基于资源竞争的内部关系去发展进化,因此,在整个生态位空间中,各个物种的生态位不可能达到比较好的分配。生态学过程发生变化时,物种多度格局会从一个模型形式转变为另一个模型形式。例如,在中等迁移速率的条件下,物种多度分布会表现出对数正态分布,图形左偏说明新迁移进来的稀有种规模较小;当迁移增加时,稀有物种的迁移规模逐步提高,直至达到很高的迁移水平并主导多度分布,就会变成几何级数或者对数级数分布[22-23]。
2.3 入侵引起物种更替
入侵物种对乡土物种的竞争与排斥作用可能导致乡土物种的消失与灭绝。外来植物往往是先锋植物,有强大而快速的繁殖能力,有迅速扩大种群的能力和极强的竞争力。研究发现,成功的外来物种在新栖息地的环境条件下竞争能力往往强于处于相似生态位的乡土物种,在这种情况下外来物种可以通过排挤乡土物种而获得成功。外来物种被引入异地后,由于其新生环境缺乏能制约其繁殖的自然天敌及其他制约因素,其后果便是迅速蔓延,大量扩张,形成优势种群,并与当地物种竞争有限的食物资源和空间资源,直接导致当地物种的退化,甚至被灭绝,即“天敌释放假说(Enemy release hypothesis)”[24]。很多入侵物种通过竞争排挤和取代当地植物而很快形成单种优势群落,造成生物多样性不可逆转性的降低,危及当地物种的生存,甚至导致当地物种特别是珍贵植物资源的濒危或灭绝,最终导致生态系统单一和退化,改变或破坏当地的自然景观。
如前所述,化感作用是紫茎泽兰种群竞争扩张的一个重要策略。外来植物分泌的化感物质往往是土著植物无法短期适应的新毒素,新化感物质会成为迅速排挤土著植物的“利器”,从而有助于入侵植物种群的快速建成[25],即入侵植物的“新武器假说(Novel weapon hypothesis)”。另外,紫茎泽兰分泌的化感物质可以抑制土壤病原真菌对其的影响[26],同时,紫茎泽兰的入侵可以提高土壤的有效磷、速效钾、硝态氮、氨态氮含量[27],从而创造对自身生长有利的土壤环境,形成一个自我促进(self-reinforcing)式的入侵机制,加剧物种间的更替[10]。
3 喀斯特生境中外来植物入侵的可能机制
3.1 内禀优势假说
一般认为,成功入侵的外来物种必然具备某些独特的内禀优势(如形态、生理、生态、遗传和行为等),即“内禀优势假说(Inherent superiority hypothesis)”[5,28]。由于其独特的内禀优势,入侵物种常常能占据多样化的生境,具有较强的适应性、繁殖力和散布力,其在新栖息地的竞争能力也往往强于处于相似生态位的乡土物种,在新生境中很容易抑制或排挤本地物种[29],导致入侵地物种多样性及遗传多样性的丧失。研究发现,紫茎泽兰之所以拥有强大的适应能力和在逆境下的生存能力,这与其植物体内的热激蛋白是密不可分的,热激蛋白具有多样性,存在于多种细胞器中,可以减轻逆境胁迫对植物体引起的伤害[30]。紫茎泽兰已被证明是多克隆植物,基因型具有多样性[31],不同的基因型在不同水平上得到表达,对紫茎泽兰入侵而言,使其适应环境的范围和生态位更大,降低了个体被淘汰的机率,增加了它的可入侵性。
3.2 资源机遇假说
“内禀优势假说”虽然充分肯定了外来物种本身的内禀优势在入侵中的作用,并且在统计意义上来讲,并且较好地解释绝大多数外来物种的入侵性。但是,必须注意到,在入侵形成的过程中,除了外来物种本身需具备入侵的特性外,资源的可利用性亦应起到关键作用。如果不考虑资源问题,单依靠物种的内禀特征,还是无法解释外来物种为什么只在特定地区、特定生态系统中发生入侵的现象。“资源机遇假说(Resource opportunity hypothesis)”认为,可利用资源是决定生态系统可入侵性的关键因素,群落一旦具有入侵物种必需的生态资源,包括营养、食物、水分、光照、土壤等,而且这些生态资源大多没有被乡土物种有效利用,即存在空余生态位,一旦有外来物种的繁殖体进入,便会发生生物入侵[32]。一般认为,若外来植物在新栖息地光、热、水、营养等资源可获得性好,则易于入侵,而在高温、高盐等不利生境则可能限制其入侵[33]。同时,入侵物种对物种多样性的影响明显依赖于生境条件[34]。如在某些案例中,物种多样性丰富的地区反而还有较多的入侵者个体,这并非生物多样性屏障机制失效,而可能是由于生物多样性丰富的地区往往资源可获得性较好的原因[35]。
然而,喀斯特生境严酷,资源水平并不高,“资源机遇假说”似乎并不能有效解释外来植物的快速入侵以及入侵的严重性,对于贵州喀斯特的外来植物入侵,仅靠一种机制似乎都不能很好解释其入侵效应的严重性。那么可能的解释是,“内禀优势假说”和“资源机遇假说”交互作用的共同结果。一些乡土物种由于喀斯特资源限制不能在该区很好生长、繁殖,而外来植物由于其独特的内禀优势可以耐受该严酷生境,即喀斯特生境对入侵物种和乡土物种的相反作用极大减小了两类物种的共存机会,加之入侵物种强大的扩散能力、繁殖力,一旦入侵,将会迅速扩散,对喀斯特生态系统产生恶劣的影响。综上所述,关于外来植物入侵对喀斯特物种多样性的影响与机制,不仅要考虑外来植物本身的内禀优势,而且要注意喀斯特资源环境的影响,特别要关注入侵物种内禀优势和资源环境的交互作用,二者的交互可能是比各自单独作用更有效的机制来解释外来植物对喀斯特生态脆弱区乡土物种多样性的重要影响。
4 未来研究重点
将未入侵群落与入侵群落相结合,通过入侵种去除前后物种多样性和群落功能的比较,有助于推断生态入侵研究中的因果关系[36]。因此,要正确认识外来植物及其入侵后果,有必要将未入侵群落(即乡土群落)、入侵群落以及入侵种去除群落整合到一个框架当中,在入侵强度梯度上,明确不同群落间物种多样性的差异以及入侵种去除后的群落构建动态,这可能是全面了解外来植物对喀斯特乡土物种多样性影响与机制的重要途径(图1)。同时,还要关注乡土群落本身的可入侵性等。明确这些将为外来植物的入侵机制提供独到见解,也可为诸如喀斯特等生态脆弱区的外来植物防治提供理论依据。
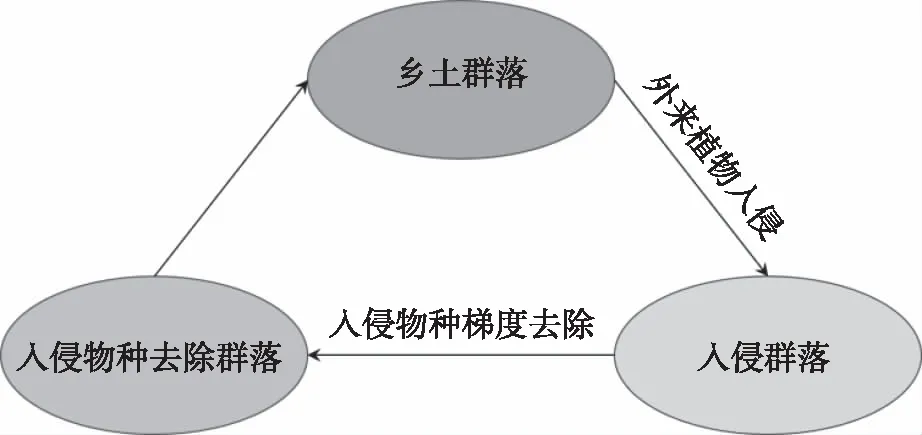
图1 外来植物入侵对喀斯特乡土群落影响的研究框架Fig.1 Research framework of the impact of alien plant invasion on karst native communities
4.1 入侵强度的影响
目前,关于入侵物种对乡土物种多样性影响的研究,主要是通过“空间代替时间的方法(Space for time substitution approach)”,即通过野外调查,比较入侵和未入侵群落间的差异来揭示[3]。事实上,入侵群落和未入侵群落间的差异具有很多不确定性,毕竟二者的初始条件并不完全相同。重要的是,采用这种方法的研究,常常选择了“最糟糕情况(Worst-case scenarios)”,即选择外来物种入侵强度大或入侵影响比较严重的群落进行比较研究[37],但这不能提供入侵强度不同(即入侵物种多度的梯度变化)而引起的入侵效应差异[3]。因此,“梯度分析法(Gradient approach)”便应运而生了,即在入侵物种的不同入侵强度上研究外来物种的入侵效应[34]。然而,自然界寻找不同入侵强度的群落比较困难,重要的是这些群落仍然具有很多“事前”的不一致性。因此,要深入了解外来物种对乡土物种多样性的影响,仅仅研究已发生的入侵是不够的,需要针对特定类群开展受控实验,通过跟踪不同入侵强度的外来物种对入侵地物种多样性的动态影响,才能有效探究入侵效应及其作用机制[38]。“梯度分析法”与受控实验结合,将会给研究者带来全新的研究视角。在研究入侵物种对所侵入的生态系统带来的影响时,人为引进入侵物种的方法是不被提倡的,去除试验为人们研究物种多样性与物种入侵间关系提供了一种行之有效的方法[39-40],通过入侵物种的梯度去除可以实现不同入侵强度的控制。从管理和恢复物种多样性的角度考虑,入侵物种的梯度去除效应将会提供更多的信息,毕竟完全去除入侵物种是不现实的。
4.2 外来植物去除后的群落构建动态
一个物种从群落中减少或消失通常会对群落中剩下的物种产生强烈的作用,尤其当该物种是群落的优势物种时[41-42]。研究表明,外来植物互花米草(Spartinaalterniflora)去除后,剩余乡土物种可以得到更多的空间和资源,植物群落分株密度升高,群落内剩余物种占比有所上升,为提高群落的稳定性提供了可能[42]。这与很多研究结果相似,去除一个或多个功能群至少在短期内能在一定程度上促进了保留种的生长[43-44]。整合分析表明,入侵群落仅仅包含了入侵物种去除后群落平均30%都不到的物种多样性[3]。相关的研究也表明,去除入侵物种对乡土群落有很强的正向作用,表现为去除后群落的物种多样性、群落的盖度、以及群落的更新均相应改善[45],入侵物种的去除使得其与未入侵群落间的相似性逐渐增加[36]。可以预见,外来植物去除后可能会促进乡土群落的正向演替,继而促进乡土物种多样性的恢复。
4.3 喀斯特乡土群落的可入侵性
考虑到生物入侵能够通过多种方式改变生态系统的功能而且改变的程度也存在差异(如增加生物量产生但是降低营养可利用性;同质化功能多样性但是增加单个的生态系统功能/服务)[16,46],在生物多样性—生态系统功能及多营养级互作的背景之下考虑喀斯特乡土群落的可入侵性也是十分重要的。尽管国内外学者围绕生物多样性与外来物种入侵开展了大量研究,但由于两者间的关系非常复杂,这也使得外来物种入侵和乡土群落物种多样性间仍存在着很大的不确定性。喀斯特群落的可入侵性不仅依赖于乡土物种多样性的高低,还依赖于群落中的物种组成,如不同的功能群组成、不同的稀有种和常见种比例等。
5 结语
针对贵州省喀斯特脆弱的生态环境,外来植物的入侵引发了一系列的生态学问题。遏制生态退化,根本解决外来植物的入侵是关键。在贵州典型的喀斯特区域,通过控制试验创设外来植物入侵限制因子的环境资源梯度(包括资源的可利用性以及资源的异质性),通过梯度去除的方式创设不同入侵强度,构建外来植物入侵群落、未入侵群落以及入侵物种去除群落间的物种多样性关系及其时间动态,分析外来植物入侵对乡土物种多样性影响及其环境依赖性,度量入侵种内禀优势和入侵地资源及其交互作用在外来植物入侵过程中的相对重要性,这可能是全面理解外来植物入侵过程和寻求减小入侵影响的有效途径。通过喀斯特生态系统的外来植物入侵研究,不仅可以丰富入侵生态学的内容,而且可为其他生态脆弱区的其他外来物种入侵的影响评价和控制策略提供理论依据和实践指导。
