土壤有机碳同位素组成在农田生态系统碳循环中的应用进展*
2023-02-17李发东栗照鑫乔云峰刘山宝HubertHirwaSimonMeasho
李发东, 栗照鑫, 乔云峰, 刘山宝, 田 超, 朱 农,Hubert Hirwa, Simon Measho
(1.中国科学院地理科学与资源研究所生态系统网络观测与模拟重点实验室 北京 100101; 2.山东禹城农田生态系统国家野外科学观测研究站/中国科学院禹城综合试验站 禹城 251200; 3.中国科学院大学资源与环境学院 北京 100190)
大气中的二氧化碳浓度(CO2)从1750年工业时代开始时的277.0 mg·kg-1增加到2019年的(409.8±0.1) mg·kg-1[1], 使得大气平均温度上升1.1 ℃。全球变暖会造成如极端气候频发、海平面上升、淡水资源短缺、危害人体健康等[2]诸多危害。因此, 治理全球变暖成为许多国家的目标。2020年第75届联合国大会上, 我国向世界郑重承诺力争在2030年前实现碳达峰, 努力争取在2060年前实现碳中和。为了达到这一目标, 我国一直在积极探索固碳减排的有效途径。植树造林被认为是固碳减排的有效方法,与森林面积的变化和已建森林的增长相对应, 中国森林生态系统的碳库在这几十年中显著增加[3]。草原是一个高度动态的土地利用系统, 提供重要的生态系统服务功能, 具有较强的碳储存能力[4]。此外,湿地生态系统是生物圈中土壤碳的重要储存库, 在全球碳循环以及应对气候变化过程中发挥着重要作用[5]。国际学术界在研究森林、草原、湿地等生态系统对增强土壤碳汇作用的过程中, 发现农田生态系统在固碳方面的作用越来越重要[6-7]。在农业领域,中国积极倡导秸秆还田, 使得全国范围内表层土壤(0~20 cm)的平均土壤有机碳(SOC)储量从1980年的28.6 Mg(C)·hm-2增加至2011年的32.9 Mg(C)·hm-2[8]。有机质是土壤的重要组成部分, 其数量和质量在一定程度上反映生态系统初级生产力和次级生产力的大小, 因此, 与土壤肥力、碳截获潜力密切相关。土壤有机碳库是陆地生态系统的最大碳库, 约占66.7%~75.0%, 其轻微的变化会对大气的CO2浓度产生很大影响, 因此SOC对研究全球气候变化和碳循环具有重要意义, 农田生态系统中SOC稳定性过程和机制也已成为陆地生态系统碳循环研究的热点问题[9]。对农田生态系统土壤碳循环的研究, 将有助于预测未来大气中CO2浓度的变化以及理解大气圈和生物圈相互作用机制。
稳定碳同位素技术是研究土壤碳循环最为科学有效的方法之一[10], 已在农田SOC周转[11-12]、土壤呼吸及其来源区分[12-13]、地上与地下碳分配[14]等方面得到广泛应用。然而, 运用稳定碳同位素技术针对农田管理方式影响下的土壤碳周转、分配及碳固持效应的机理性研究目前开展的较少。自然界中存在7种碳同位素(10C、11C、12C、13C、14C、15C、16C), 只有12C、13C两种为稳定性碳同位素, 其中12C、13C天然丰度分别为98.89%、1.11%,14C只有极微量且具放射性, 半衰期为5730年; 其他同位素由人工核反应获得, 均有放射性且含量极少[15]。不同同位素之间物理化学性质上的差异使反应前和反应后的物质在同位素组成上存在差异, 这种现象称为同位素分馏效应[16-17]。基于此, 利用稳定碳同位素技术研究农田生态系统碳循环是一种有效的手段, 同时也是研究农田生态系统有机碳稳定性的有效工具。稳定性同位素(12C 、13C)与放射性同位素(14C)相比, 具有安全、无污染、易控制等优点, 被广泛应用于陆地生态系统碳周转的生物地球化学过程研究[18]。
目前, 国内一些研究人员分别从农田耕作制度[19]、施肥方式[20]、秸秆还田[21]等方面研究了农田生产力、碳固持速率、温室气体排放效应等问题。但至今对如何提升农田生态系统有机碳稳定性的机制认识尚未完全明确, 导致农田生态系统碳储量和关键碳周转过程的估算还存在很大不确定性, 迫切需要深入开展农田碳固持潜力的机制研究。
1 不同耕作措施对土壤有机碳固持过程的影响
缓解气候变化和确保由于全球人口呈指数增长所带来的粮食安全问题是未来农业面临的最大挑战。改进土壤耕作措施对于通过提高农业生态系统生产力、土壤肥力和碳固持能力来解决这些问题至关重要[22]。应对气候变化需要降低大气中的CO2浓度,这可以通过减少CO2排放量和增加碳汇来实现[23]。土壤中碳的储量和存储时间在农业生态系统中分别是最高和最久的, 耕作措施会影响其储量和周转周期, 采用有效的耕作措施会使土壤成为碳汇, 增强土壤碳的稳定性[24]。SOC含量是一个重要的土壤质量指标, 对包括初级生产力、气候调节和生态系统服务在内的不同土壤功能具有重要影响, 其损失是加剧气候变化的主要原因之一[25]。SOC的动态平衡过程决定着土壤碳储存过程, 因此研究SOC的动态变化对揭示不同耕作措施影响下的土壤碳固持过程以及调控机制具有重要意义。
在进一步深入探讨之前, 需要先了解几个重要概念。首先是碳饱和度, 是指SOC存量的增加与碳输入不呈正比, 这意味着土壤并不是一个无限的碳汇[26]。产生这一现象的原因主要与土壤中矿物粒度百分比有关, 尤其是细粒矿物(<20 μm)[27]。它们通过有机-矿物相互作用(物理-化学相互作用)保护有机碳不被淋滤, 或限制有机化合物进入土壤微生物群落, 或阻碍分解者的氧气供应(物理保护)[27]。其次是碳固持, 是指将大气中的CO2通过植物、植物残留物和其他有机固体转移到土壤中的过程, 这些有机固体作为土壤有机质(腐殖质)的一部分储存或保留在土壤中[28]。某一耕作措施的碳固存潜力代表在特定气候、特定时间段和特定土壤深度(主要是0~20 cm)下SOC存量的最大增加或减少量。总的来说, 负碳固存潜力代表SOC的净损失, 而正碳固存对应SOC存量的增加[29]。最后是碳储存, 是指在特定土地单元和一定土壤深度以及时间段内SOC储量的增加[29]。总的来说, 碳储存被定义为特定时间段内特定土壤深度的碳质量变化, 而碳固持则被定义为特定时间段内特定土壤深度的碳固定速率。因此, 碳固持更能准确反映土壤SOC动态变化过程。
不同耕作措施(如免耕、翻耕、秸秆还田、施肥等)主要通过改变土壤结构(特别是团聚体)、土壤理化性质而影响土壤的碳固持效率[30], 从而影响SOC的稳定性, 最终影响土壤碳排放过程和气候变化(图1)。

图 1 耕作方式影响土壤CO2排放的过程和监测方法展望Fig.1 Prospect of processes and measuring methods of soil CO2 emission affected by tillage method
传统耕作(如犁耕)机械地破坏土壤表面的团聚体, 使以前受保护的土壤有机质(SOM)暴露在微生物的分解环境下[31], 促进土壤侵蚀, 减少有机碳储量[32]。经证明, 采用免耕或少耕方法的农田表层土壤(0~10 cm)的有机碳含量显著高于传统耕作(如犁板耕法)[33]。然而, 随着土壤深度的增加(>10 cm), 耕作措施对SOC储量没有显著性影响[33]。耕作引起的SOC损失与土壤侵蚀有关[34], 尽量减少机械干扰可通过增强土壤团聚体稳定性来减少侵蚀[35]。不同组分碳在土壤中的周转时间是影响SOC稳定性的重要因素。随着深度增加, 矿质态SOC的稳定性强于颗粒态SOC, 但翻耕农田的矿质态SOC稳定性弱于免耕[36], 表明免耕对上述两种形态碳的固持和稳定性影响明显。传统的翻耕措施能增加土壤颗粒暴露面积, 加快大团聚体的崩解破裂, 从而加速土壤碳矿化。相反, 免耕减少了对土壤的扰动, 增加了土壤大团聚体对SOC的包裹使其减少矿化, 进而增加SOC库[37]。长期定位试验研究还认为, 免耕能显著增加0~10 cm土层SOC含量, 但与传统耕作措施相比, 10~30 cm土层SOC含量差别不大[38]。大多取样深度超过30 cm的试验中, 免耕深层SOC含量相对翻耕降低[39]。对农田生态系统而言, 由于耕作措施的改变, 土壤剖面的水分、养分、热量、微生物及根系的生长都产生变化, 从而影响了土壤碳的平衡状况, 使得耕作措施对土壤碳的影响机制更为复杂。因此, 需要用新技术手段(如稳定碳同位素技术连续观测)识别不同耕作农田SOC库的特征组分, 并了解其形成与周转机制, 对探索SOC在不同时期(长期或短期)的动态变化至关重要。然而, 免耕增加的SOC储量主要集中在表土层(0~10 cm), 而且这部分SOC大多为易分解碳[40], 其活性和矿化能力都很高, 不利于形成稳定碳长期固持下来, 而且免耕深层土壤因其物理结构的差异(如土壤容重增大), 碳的垂直运移过程受阻, 其碳的蓄积能力相比传统耕作农田要弱[41]。如何利用稳定碳同位素对比研究免耕与传统耕作农田中垂直剖面土壤碳组分的固持效应和周转机制, 是提升深层SOC稳定性的关键问题。
秸秆还田不仅可以提高SOC储量, 还可以改善土壤结构、降低容重、减少蒸发、减少侵蚀并提高土壤中的营养物质入渗速率[42]。在农业管理系统中,秸秆被用于热能生产或动物饲养/铺垫, 秸秆被剪短并移除, 这减少了作物残留物的总量[42]。一般来说,玉米(Zea mays)或多年生作物产生大量残留物, 而马铃薯(Solanum tuberosum)等根茎作物产生的残留物较少。残留碳氮质量也会影响SOC, 碳氮比较低的作物残留[如大豆(Glycine max)]会促进微生物分解,这将促进土壤有机质的分解[43]。碳氮比较高的作物残留物(玉米)通常与有机碳累积有关。在类似的作物残留管理下, 有机碳储量随产量的增加而增加[44]。尽管如此, 由于生长调节剂的应用和短茎作物类型的选择, 作物残留量可能不会随作物产量呈比例增加。然而, 前人的研究也指出低碳氮比的作物残留物可能会增加有机碳储量[45]。在对长期野外原位试验的荟萃分析中, 残留物掺入使SOC存量增加了7%[46]。同时, 免耕+秸秆还田被认为可增加碳固持效率、减少CO2排放。此外, 免耕和秸秆还田相结合较少扰动土壤, 增加了土壤肥力与养分, 提高作物产量, 已被认为是理想的固碳、减排、增产的管理机制[47]。
施肥水平也是影响SOC固持的重要因素。尤其在华北平原, 大量的氮肥输入成为集约农业增加产量的主要渠道, 这也产生了很多不利的后果, 如增加温室气体排放、水体污染、加剧气候变化等等。合理的无机氮肥输入成为解决此问题的关键。总的来说, 施氮对SOC的影响有两种截然不同的趋势: 一方面, 氮肥为初级生产提供“燃料”, 从而提高地上和地下生物量, 进而丰富SOC储量[29]; 另一方面, 氮肥可以刺激凋落物和土壤有机质的生物降解[48], 减少SOC库存[49]。因此, 最佳氮肥供应可能对土壤碳固持至关重要[50]。在一项全球荟萃分析中, 氮肥使农业土壤的有机碳减少了10%[50]; 而在另一个全球荟萃分析中, 氮肥使有机碳储量增加了3.5%[51]。土壤有机质可通过宏观和微观聚集或与淤泥和黏土颗粒的相互作用进行物理稳定[52], 或通过形成难降解的SOM化合物进行生化稳定[53]。在氮有限的环境中, 土壤微生物群落利用来自稳定SOM的氮来满足其生长和繁殖需求[54]。氮肥可通过促进凋落物分解, 同时减少微生物养分利用, 改善稳定有机碳库(细粒径<0.4 mm)[55]。然而, 施氮肥对SOC固持过程的影响还与土壤质地有关, 矿物氮肥添加倾向于增加土壤中的颗粒有机碳, 而淤泥和黏土粒级组分中的有机碳则减少, 产生不太稳定的SOC[56]。与单独施用矿质氮肥类似, 化学氮肥、磷肥和钾肥(NPK)增加了作物产量[13,53]和植物生物量、土壤微生物量以及SOC存量[57]。与不施肥条件相比, 一项长期野外试验的荟萃分析表明, NPK、N、P和K施肥可分别将上层土壤(0~20 cm)的SOC储量提高10%、5%、5%和2%[58]。此外, 氮和磷肥增加了根系生物量和根系分泌物, 从而提高了SOC储量。不同土壤类型和种植制度的长期施肥试验表明, 矿物NPK肥料对SOC固存的影响取决于土壤参数, 主要是土壤有效氮含量和土壤pH[59]。除了提高初级生产力外, 磷肥还可以通过影响丛枝菌根真菌影响土壤固碳。与单独施用氮肥相比, 施用NPK可减少丛枝菌根真菌的定殖, 从而减少菌根介导的植物养分吸收, 进而对土壤碳固存产生负面影响[60]。然而, 丛枝菌根真菌在陆地碳循环中的作用在很大程度上被忽视, 尤其是其对碳固存潜力的影响[61]。总的来说, 土壤微生物群落在控制农业生态系统碳循环中起着重要作用[62]。碳利用效率决定了碳在分解过程中的去向, 与氮和磷的有效性密切相关, 并可能进一步受到钾和微量营养素可及性的影响[63]。这些联系需要通过进一步的研究加以解决。
2 土壤碳排放对土壤有机碳稳定性的响应机制
SOC是影响土壤质量和农业可持续性的主要土壤成分, 农田管理措施对土壤的碳固存有很大影响。保持和改善SOC对于缓解集约化种植系统中土壤结构恶化和CO2排放至关重要, 因为SOC被认为是一种重要的聚合结合剂和具有强大的碳汇功能[64]。土壤碳排放过程被认为与SOC稳定性密切相关, 一般来说二者表现出负相关, SOC稳定性减弱有利于土壤碳排放[65]。一方面, SOC稳定性降低代表矿化过程增强, 可产生更多有利于提高微生物活性的底物,促进土壤碳排放[66]; 另一方面, 为根系生长提供更多可利用元素, 促进根系活动, 增加土壤碳排放强度[67]。
土壤碳排放过程主要通过土壤呼吸来完成, 土壤呼吸主要由土壤微生物、动物呼吸组成的异养呼吸与根系、根际呼吸组成的自养呼吸构成(图2), 对农田生态系统土壤碳平衡有重要影响。目前, 关于土壤呼吸的研究主要集中于区分根呼吸和微生物呼吸两个生物学过程[68-69], 而上述过程在呼吸性质、呼吸主体、利用碳源、周转速率、时空变异性以及对环境因子的响应和适应方面差异显著[70]。土壤微生物呼吸是土壤对大气CO2浓度增加的主要贡献者,故只有区分根呼吸与土壤微生物呼吸才能准确探研SOC稳定性。在Zacháry等[71]的研究中指出通过测定轮作农田生态系统中不同作物生长时期的SOC的δ13C值, 可以定量描述SOC 的平均周转时间, 进而衡量SOC稳定性。SOC稳定性与δ13C关系的研究有很多[72-74]。然而, 近30年来, 利用实时原位稳定同位素技术区分土壤呼吸碳来源从而揭示SOC稳定性机制的研究尚不多见。因此, 以后的研究要注重典型农田生态系统的原位观测, 使用13C稳定同位素技术, 通过连续、实时地监测土壤中各SOC组分含量, 解析不同碳组分[易分解SOC、微生物生物量碳(MBC)、惰性SOC]对土壤呼吸的贡献, 以及SOC各组分碳的周转速率和内在稳定性机制, 为研究典型农田生态系统SOC不同组分稳定机制及固持效应提供科学依据。
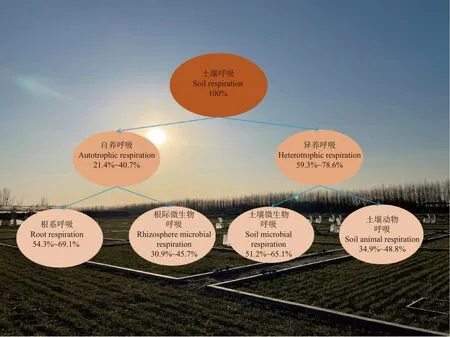
图 2 典型农田生态系统土壤呼吸组分结构图(华北平原)[69,79-81]Fig.2 Structure of soil respiration components in a typical farmland ecosystem (North China Plain)[69,79-81]
此外, 土壤呼吸各组分对环境因子响应不同, 对土壤活性碳和惰性碳的利用和分解也不一致[75]。因此, 区分土壤CO2不同组分的来源和贡献, 对理解不同碳组分的周转效应及SOC稳定性非常关键。目前为止, 许多方法用来区分土壤CO2, 如根系去除法、壕沟法、环剥法、物理分割组分法、直接测量方法及同位素技术[76]。不同组分的碳同位素值差异可定量不同呼吸组分的比例及组分之间的净交换, 自然丰度δ13C常用来区分土壤CO2通量, 其具有破坏小且不改变环境状况等优势而得到广泛应用[70,77], 使得土壤呼吸组分分离技术和相关机制的研究取得了较大进展。然而, 由于农田生态系统受人为活动的强烈影响, 农田水、肥的短期变化会严重影响土壤呼吸及SOC的稳定性[78], 运用现有的土壤呼吸分离方法来探索短期内农田土壤碳循环特征时, 表现出一定的局限性。深入分析农田生态系统中土壤呼吸的源、呼吸主体和呼吸过程, 亟需长期连续观测、整合利用现有土壤呼吸观测技术, 改进现有组分分离法和根分离法, 强化土壤呼吸组分和环境因子的同步观测, 以准确地评估农田碳收支。利用土壤呼吸各组分的实时、定位观测, 研究不同土壤呼吸组分对不同环境因子(如孔隙度、团聚体、温度、湿度、养分、腐殖质)、气候因子及生物因子的交互响应机制, 是重构现有全球碳模型中碳过程和土壤呼吸过程机理的重要手段。
3 13C 同位素技术在土壤有机碳组分固持中的定量识别
以SOC形式固持的碳库是减缓大气CO2浓度上升的全球重要碳蓄积库[16], 其含量达1400~1600 Pg,是大气碳库的3倍, 约是陆地生物量的2倍[82]。农田生态系统占陆地生态系统40%左右, 过去150年, 农田管理方式对土壤的扰动导致全球40~90 Pg的碳损失[83-84], 而通过增加深层土壤碳固持, 每年可固定5.5~6.0 Gt CO2当量碳[85]。因此, 土壤碳库的微小变化将影响大气CO2浓度及全球碳循环过程[86]。另外,未来30年SOC库提升是开展有机农业、提升粮食产量以满足全球90亿人口粮食需求的重要措施, 其碳库提升对减缓气候变化、维持农田生产力和粮食安全具有重要意义[87]。目前, 大量研究评估了不同气候环境和农田管理方式对SOC的固持效应, 而中国农田SOC储量占全球平均水平的1/3左右[88]。因此,提升我国农田土壤碳库水平和土壤肥力状况, 降低农田碳排放, 是目前亟需解决的科学问题之一。
当前, 不合理的农田管理措施致使大量的SOC损失, 合理的实践措施会增加土壤SOC含量以及增强碳汇功能。寻求一种或多种耕作措施之前理解其内在机制是关键, 而13C同位素技术是解决此问题的有力工具。13C同位素技术是研究限制全球碳循环不确定成分的最有效手段之一, 如陆地生物圈和大气储层之间的净碳交换以及农业生态系统碳储存大小和分配[89]。
目前利用13C稳定同位素研究主要集中于不同组分碳(易分解SOC、MBC、惰性SOC)和不同粒径团聚体(大团聚体和微团聚体)碳含量的周转速率和固持效应等。Dou等[11]使用δ13C同位素技术研究长期(25年)秸秆还田后农田不同等级土壤团聚体SOC含量, 发现秸秆还田配合无机氮肥施用, <250 μm团聚体中的SOC含量减小, 而>250 μm的SOC量逐渐增加, 显示轻组有机碳向大团聚体中颗粒有机碳的转变。
稳定性同位素技术适合研究从年到百年尺度的土壤碳循环过程, 能有效地阐明地下碳动态变化和土壤碳储量的微小迁移与转换, 以及定量化评价新老SOC对碳储量的相对贡献[90]。13C自然丰度法同位素技术在农田SOC的周转时间和不同碳组分分配上已有广泛研究[91-92]。农田生态系统中主要包括C3和C4两种光合作用途径的植物类型, 其中C3植物δ13C处于-35‰至-20‰范围之间, C4植物δ13C为-19‰至-9‰[92]。通过转换种植C3和C4作物, 秸秆还田后测定SOC不同组分δ13C值, 可进一步解释秸秆还田对土壤碳循环过程的影响, 推断土壤碳组分的周转时间[73]和SOC库周转速率[93]。通过标记秸秆研究作物秸秆、残茬或作物根系在土壤中的分解动态或对土壤有机质的贡献, 可为阐明土壤碳转化过程及土壤肥力演变提供新的技术支撑[94-95]。稳定碳同位素还可利用脉冲标记法研究碳在植株地上部、地下部以及土壤中的迁移和转化过程。脉冲标记法分为连续脉冲和间断脉冲两种, 可视研究需要进行选择。利用4次13C脉冲标记研究光合碳在互花米草(Spartina alterniflora)中分配和迁移过程发现, 互花米草各组分固定13C量呈现茎>叶>根>根际土壤>土体的趋势, 叶分配比例为32.4%~39.2%, 茎分配比例为39.8%~47.6%, 根分配比例为18.0%~20.3%, 根际土壤分配比例为0.3%~0.4%, 土体分配比例为0.08%~0.20%[96]。研究冬小麦(Triticum aestivum)生长向地下部分的输入对土壤固碳和作物生产具有重要意义。有研究分别在分蘖期、拔节期、花期和灌浆期, 用13CO2对冬小麦进行脉冲标记7 h, 发现在整个生育期内, 冬小麦净吸收的光合碳分配到地上部、根部及SOC和根际呼吸的比例分别为78.5%、6.0%、3.1%和12.4%[97]。此外, 注射法也可用于研究稳定碳同位素在植株体内及土壤中迁移和周转过程。注射法是指定期向植株体内注入稳定碳同位素肥料溶解液, 可通过叶片或茎部进行注射。然而, 大多研究基于室内培养的短期研究, 侧重于通过外源碳素添加来探索不同碳组分及其周转的贡献, 而对于野外耕作措施结合秸秆还田等农田管理方式对碳库的稳定机制研究还较少。
利用多种稳定碳同位素模型研究SOC稳定性也是未来一大趋势。已有国外研究利用WATEM模型模拟遭受侵蚀景观中的有机碳循环[98]。因为SOC是由不同组件组成的复合体, 所以在诸如CENTURY[99]、ICBM[100]等模型中, 它通常由关于碳输入和分解率的各种池来表示。Bouchoms等[101]还开发了模型来研究能量消耗、作物生产力和有机碳循环之间的关系。在农田尺度上, 结合SOC侵蚀再分布和SOC动态的模型现在能够重现土地利用下农田SOC储量的空间异质性、侵蚀区SOC耗竭、沉积区SOC富集[102-103]。碳同位素也被包括在有机碳循环模型中, 以约束模型参数或探索控制因素。Baisden等[104]利用碳和氮同位素模拟了SOC沿深度的周转和迁移, 表明水文条件在控制SOC的垂直迁移中起着重要作用。此外,整合碳同位素识别的有机碳循环模型被用于探索有机碳分解和物理混合对δ13C值随土壤深度垂直增加的影响[105]。Wynn等[89]建立了一个关于δ13C (SOC)的主要气候控制模型, 该模型植根于C3和C4植被的优化水利用效率的假设, 并且可以用现成的环境数据推断到大陆或全球数据。Ahrens等[106]使用14C特征来约束使用贝叶斯方法的多池有机碳模型的参数, 并且该模型被进一步应用于量化吸附、溶解有机碳迁移和微生物相互作用在确定土壤剖面δ14C值中的贡献[107]。虽然野外研究已经确定了土壤再分布对有机碳同位素剖面的影响[108], 但相关模型尚未开发。包含碳同位素的SOC模型适用于农田生态系统, 将有助于全面了解碳同位素剖面以及SOC同位素组成在农田生态系统中的空间变异性, 更好地揭示SOC稳定性机制, 为减缓气候变化、加快实现碳中和目标做出积极贡献。
实时监测时空尺度CO2排放是今后研究土壤呼吸组分的趋势(图3)。近期报道, 结合δ13C示踪技术在不同时间尺度(日、季和年)[109]、不同土壤剖面实时自动监测CO2浓度变化[110], 可揭示全年土壤呼吸组分来源变化, 表明稳定同位素示踪技术作为区分农田土壤关键碳过程的重要手段, 已得到广泛运用。然而, 短时间尺度监测并不能完全区分土壤呼吸组分来源以及不同组分碳固持速率。运用δ13C同位素探索土壤碳周转和稳定性固持的内在机制研究尚未完全开展。加强长期实时监测不同组分土壤呼吸及碳固持动态变化, 是精确评估农田生态系统碳过程及碳源/汇功能的关键。

图 3 土壤CO2排放实时原位监测系统Fig.3 Real time in situ monitoring system for soil carbon dioxide emission
4 展望
SOC稳定性对维持农田生态系统碳库、作物的稳产高产以及全球碳循环具有重要意义。农田土壤CO2排放是SOC耗散的主要方式, 不同农田管理措施将对SOC稳定性产生重要影响。农业生态系统在固碳减排方面发挥重要作用, 寻求合理的管理实践方式的关键是揭示SOC稳定性对农田管理措施的响应机制。SOC固持和碳排放的过程非常复杂, 响应机制的揭示必须清晰理解其中的过程, 先进的技术支持必不可少。
过去主要集中在用传统方法(如取样法)对土壤表层SOC稳定性进行研究, 很难反映有机碳在土壤中的周转和迁移过程, 不具有代表性。同时, 以往的手段不能识别土壤碳循环过程中不同有机碳组分含量的变化, 这对精确理解SOC稳定性的机制带来极大困难。对土壤碳排放的研究过去主要集中在室内小范围模拟试验且不连续, 与实际农业生态系统碳排放过程差距较大。13C同位素技术具有稳定性高、安全性强的特点, 在土壤碳固持和排放过程中能够精确识别不同有机碳组分的含量变化, 不仅能够根据单位时间内有机碳含量变化计算出有机碳稳定性,还能有效区分土壤呼吸组分、来源及贡献。结合现有的农田管理措施, 可以很好地揭示SOC稳定性的响应机制。
当前利用13C同位素技术的标记示踪和室内培养的方法, 揭示了其中重要的机理, 但与野外实时的状况仍有差距。土壤碳固持和排放的复杂过程受诸多环境因素的影响, 如大气温度、湿度、辐射、气压等, 土壤质地、结构、基础理化性质、微生物群落等以及人为因素。由于土壤碳循环的复杂性和易干扰性, 单纯的室内模拟试验很难揭示其内在过程和机理。
因此, 未来研究应加强农田生态系统土壤CO2排放的原位观测和实时动态观测, 并通过结合δ13C同位素丰度和环境要素的实时观测, 结合采样后的土壤和秸秆理化分析, 可以实现土壤CO2排放的实时分解, 从而揭示SOC稳定性对不同农田管理措施的响应机制, 为典型农田生态系统SOC、农田地力、土壤碳固持潜力的提升提供科学依据, 更好地实现农业可持续发展和国家碳中和目标。
