长枝木霉菌对水稻耐盐碱特性的影响
2023-02-11崔皓钧王文奕李岩刘琳马书荣
崔皓钧,王文奕,李岩,刘琳,马书荣
(东北盐碱植被恢复与重建教育部重点实验室(东北林业大学),哈尔滨 150040)
0 引言
在世界范围内盐碱土的面积约占陆地总面积的10%,盐碱地分布在干旱的荒漠和半荒漠地区,还存在于沿海及灌溉地区[1]。我国具有9 913万hm2的盐碱地,在东部的沿海地区、西北干旱地区、东北平原地区均有分布[2-3]。土壤盐碱化限制农业发展,威胁生态环境[4-5]。盐碱土中大量的盐离子毒害植物,使植物生长发育停滞,植物萎蔫加速。土壤中过量的盐碱离子,也会使土壤的肥力减弱[6-7]。现如今,如何高效地利用盐碱地,发挥最大的生态效益,促进农林业经济发展,是首要解决的任务。
木霉菌(Trichoderma)是世界上广泛分布的丝状真菌[8]。木霉菌可以提高植物抗病性,拮抗植物病原菌,还可以在盐碱胁迫下,减弱盐碱离子对植物的毒害作用,提高植物耐盐性,促进植物生长。木霉菌介导的植物耐盐性已在多种植物中得到证实,如Zhao等[9]发现棘孢木霉(Trichodermaasperellum)Q1对盐胁迫下黄瓜生长的促进作用,结果表明该菌株具有很好的溶解磷酸盐能力,使黄瓜幼苗叶片中吲哚乙酸(IAA)、赤霉酸(GA)和脱落酸(ABA)的水平增加,同时植株的根系生长和根系活力也增加。Contreras等[10]发现绿色木霉(Trichodermavirens)和深绿木霉(Trichodermaatroviride)在正常和盐条件下都能促进拟南芥(Arabidopsis)生长,使拟南芥的脱落酸、L-脯氨酸和抗坏血酸水平增加,同时还可以提高拟南芥的IAA水平以及抗氧化能力。可见,通过分离筛选获得木霉菌,作用于植物,可以作为提高植物抗盐碱性、促进植物生长的重要手段。
本研究旨在从盐碱地蒙古柳(Salixlinearistioularis)侧根分离出长枝木霉菌,通过盐碱处理及接菌实验,分别从生物量、生理生化指标及光合作用等几个方面,探讨了长枝木霉菌对水稻种子萌发和幼苗生长的影响,分析了长枝木霉菌对水稻的解盐促生功能,为进一步研究和利用长枝木霉菌缓解盐碱胁迫促进植物生长提供理论基础。
1 材料与方法
1.1 材料
供试植物:本试验所采用的水稻品种为长白9号。
长枝木霉菌株:从黑龙江省松嫩平原盐碱地上自然生长的蒙古柳侧根分离、纯化获得,并进行分子鉴定,GeneBank登录号NR120298.1。
供试土壤:园艺土与蛭石以3∶1的比例混合,装入灭菌袋,121 ℃,60 min。
1.2 试验方法
1.2.1 孢子悬浮液的制备
将长枝木霉菌株接种于PDA培养基上,28 ℃,黑暗培养7 d至完全产孢,用枪头刮取培养基上的菌丝和孢子,放入无菌水中,用无菌纱布过滤,制成孢子悬浮液。血球计数板计算孢子浓度,使浓度调至2×108cfu/mL。
1.2.2 种子萌发处理
挑选籽粒饱满,大小合适的水稻种子,先用75%酒精消毒8~10 min,无菌水多次清洗,再用5%的次氯酸钠溶液消毒10~15 min,无菌水多次清洗,得到无菌水稻种子。将无菌处理种子放入2×108cfu/mL的孢子悬浮液中,浸泡6 h,对照组用无菌水浸泡6 h。之后分别将种子放在无菌水、100 mmol/L NaCl溶液、30 mmol/L NaHCO3溶液[11]中,每板20粒种子,3个重复。在第2天时,测定水稻种子的发芽势。在第3天时,测定水稻种子的发芽率。培养后第7天,每个处理取5粒正常发芽的种子测定其胚根长和胚芽长。
发芽率(%)=(发芽第3天正常发芽种子数/供试种子数)×100;
发芽势=发芽前2天正常发芽的种子数/供试种子数。
1.2.3 盆栽试验
将无菌处理后的水稻种子转移至无菌培养皿中催芽3 d。3 d后挑选长势相同的种子,移入盆钵中,每盆种植5粒种子。试验组每5 d,用20 mL孢子悬浮液浇灌于幼苗根部,对照组和试验组均每3 d浇一次20 mL的盐碱处理液,剩余2次浇灌蒸馏水20 mL[12]。
1.2.4 幼苗生物学指标的测定
盐碱胁迫处理水稻45 d后,将水稻植株连根取出,分别测量水稻的株高、地上部分干鲜重、地下干鲜重、根长和叶绿素含量。
1.2.5 生理指标的测定
过氧化氢酶的测定采用过氧化氢还原法。过氧化物酶的测定采用愈创木酚法。超氧化物歧化酶采用氮蓝四哇法测定。丙二醛(MDA)采用硫代巴比妥酸(TBA)比色法测定[13]。将水稻叶片擦净剪碎,依次放入三氯乙酸溶液,获得MDA提取液,加入硫代巴比妥酸溶液,沸水浴,离心。在比色皿中,用紫外分光光度计分别在450、532、600 nm波长下测定吸光度值并计算含量。
1.2.6 光合参数的测定
采用便携式光合作用测定系统(LI-6400)测定水稻叶片的净光合速率(Pn)、气孔导度(gs)、蒸腾速率(Tr)和胞间CO2浓度(Ci),设置光照强度为1 000 μmol/(m2·s),固定系统内CO2浓度(体积分数,下同)为405 μL/L。
1.2.7 数据处理
用Excel 2016对水稻的各项胁迫指标的数据进行处理作图,取5次重复的平均值,用SPSS19.0采用单因素方差分析法进行显著性分析。
2 结果与分析
2.1 种子萌发的影响
在盐碱条件下,施用长枝木霉菌孢子悬浮液处理水稻种子进行发芽试验,处理3 d,统计发芽率。如图1所示,与对照组相比,无菌水条件下,长枝木霉菌显著提升水稻种子的发芽率,提升32%。盐碱胁迫能够显著降低水稻种子发芽率,抑制水稻种子发芽,发芽率仅达到75%和53%。接菌后,在碱胁迫下,长枝木霉菌提升水稻种子的发芽率为18%。而在盐胁迫下,种子发芽率显著提高,提升25%。证明长枝木霉菌能够有效缓解盐碱胁迫对种子发芽的抑制。处理2 d,统计发芽势。无菌水条件下,长枝木霉菌提升水稻种子的发芽势为0.06。而在碱胁迫下,长枝木霉菌提升水稻种子的发芽势为0.03。而在盐胁迫下,长枝木霉菌提升水稻种子的发芽势为0.2。由此可见,在盐碱胁迫下,采用长枝木霉菌孢子悬浮液处理后可减轻盐碱离子对种子萌发的抑制,提高发芽率和发芽势。
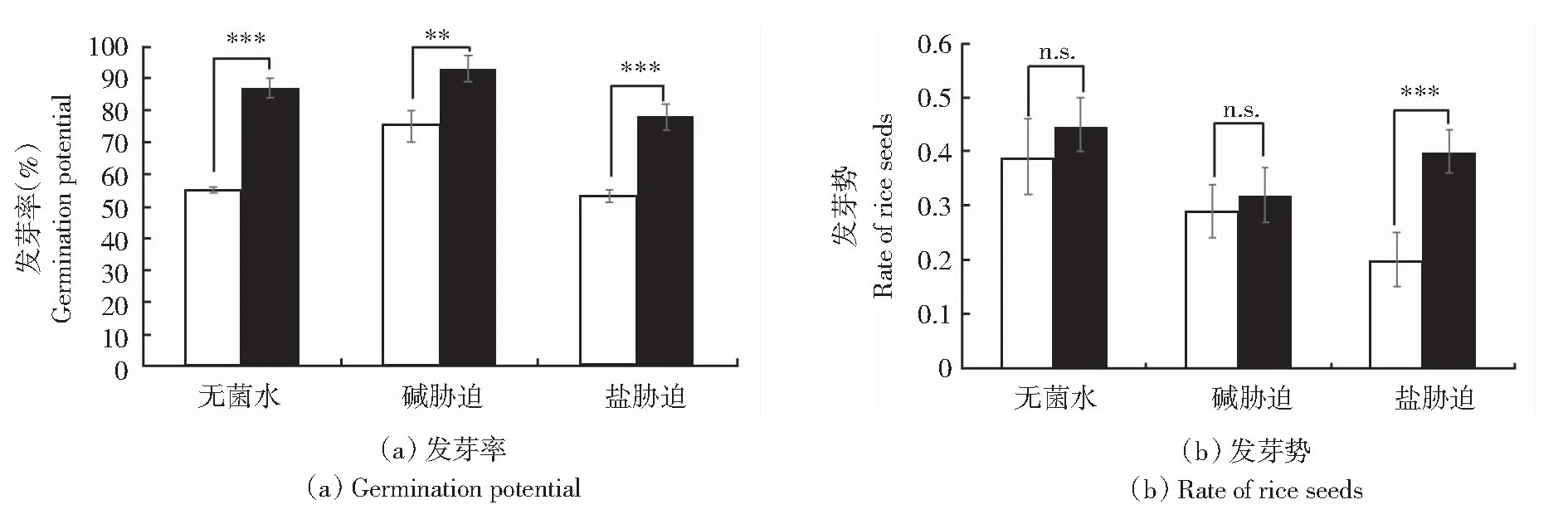
**表示P<0.01;***表示P<0.001;n.s.表示差异不显著。白色柱表示未接种对照;黑色柱表示接种长枝木霉菌株。
水稻种子培养7 d,测定水稻的胚根、胚芽。接种菌株后,促进水稻种子胚芽和胚根的生长,结果如图2和图3所示,无菌水条件下,胚芽增长14.2%,胚根增长38.9%。在盐条件下,胚芽增长50%,胚根增长50%。碱条件下,胚芽增长33.3%,胚根增长50%。与CK相比,种子的胚芽、胚轴显著提高。综上可知,施用长枝木霉菌孢子悬浮液可以有效缓解盐碱胁迫对水稻发芽带来的抑制作用,促进水稻生长。
2.2 生物量的影响
在盐碱胁迫条件下,长枝木霉菌对水稻幼苗生物量的影响,见表1。处理45 d后,在盐碱胁迫条件下,植株受到盐碱的作用,生长受到抑制。但接菌后,水稻的株高、根长、鲜重和干重均有所增加。无菌水条件下,接菌后,水稻的株高增加10.4%,水稻的根长增加21.2%,水稻的地下干重增加15.3%,水稻的地下鲜重增加17.4%,水稻的地上干重增加23%,水稻的地上鲜重增加17.4%。在碱胁迫条件下,接菌后,水稻的株高增加13.7%,水稻的根长增加6.7%,水稻的地下干重增加30%,水稻的地下鲜重增加6.7%,水稻的地上干重增加17%,水稻的地上鲜重增加10.8%。在盐胁迫条件下,水稻的株高增加2.3%,水稻的根长增加14%,水稻的地下干重增加42%,水稻的地下鲜重增加14.7%,水稻的地上干重增加17%,水稻的地上鲜重增加5.4%。试验结果表明,接种长枝木霉菌孢子悬浮液后,均不同程度地促进幼苗的生长,对水稻地上部分促生效果较为显著。

第一行为无菌水、NaHCO3、NaCl; 第二行为水+长枝木霉菌、NaHCO3+长枝木霉菌、NaCl+长枝木霉菌。
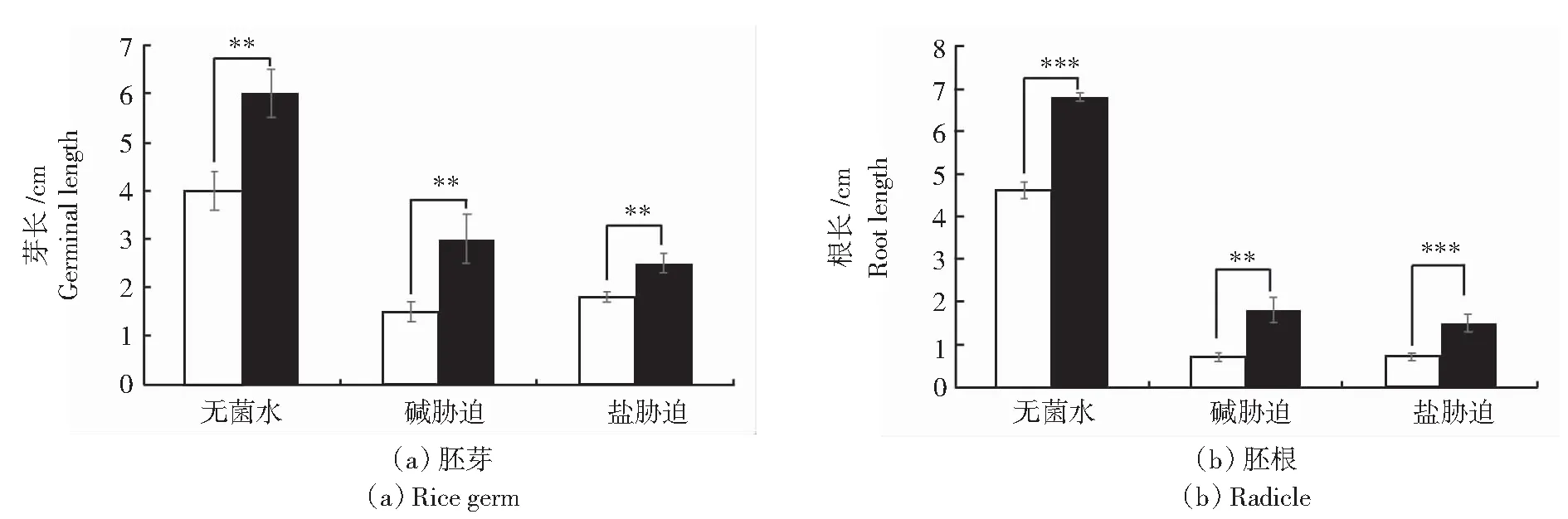
**表示P<0.01;***表示P<0.001。白色柱表示未接种对照;黑色柱表示接种长枝木霉菌株。
2.3 叶绿素含量的影响
光合作用是植物重要的生命活动,植物的光合作用为植物的生长发育提供养分和信息。盐碱胁迫会降解叶绿素,降低植物的光合作用效率[14]。如图4所示,在盐碱胁迫条件下,接菌后,叶绿素a、叶绿素b、叶绿素总量均升高。接菌后,在无菌水条件下,水稻叶片叶绿素a升高18.9%,叶绿素b升高31.8%,总叶绿素升高21.6%。在盐胁迫条件下,水稻叶片叶绿素a升高25.3%,叶绿素b升高10.3%,总叶绿素升高18.4%。在碱胁迫条件下,水稻叶片叶绿素a升高29.6%,叶绿素b升高16.3%,总叶绿素升高22.9%,综上所述,接种长枝木霉菌可增加水稻叶片中的叶绿素含量。

表1 长枝木霉菌对盐碱胁迫下水稻幼苗生物量的影响


***表示P<0.001;**表示P<0.01;*表示P<0.05。白色柱表示未接种对照;黑色柱表示接种长枝木霉菌株。
2.4 水稻叶片丙二醛含量的影响
丙二醛(MDA)的存在加剧膜的损伤程度,常作为植物受到逆境胁迫的指标。接菌后,水稻幼苗叶片内丙二醛含量的变化,如图5所示。在盐碱胁迫下,水稻叶片丙二醛的含量上升。在无菌水条件下,降低效果最明显,下降47.5%。在盐胁迫下,下降11.9%。在碱胁迫下,菌株使MDA上升2.5%。

n.s.表示差异不显著;*表示P<0.05。白色柱表示未接种对照;黑色柱表示接种长枝木霉菌株。
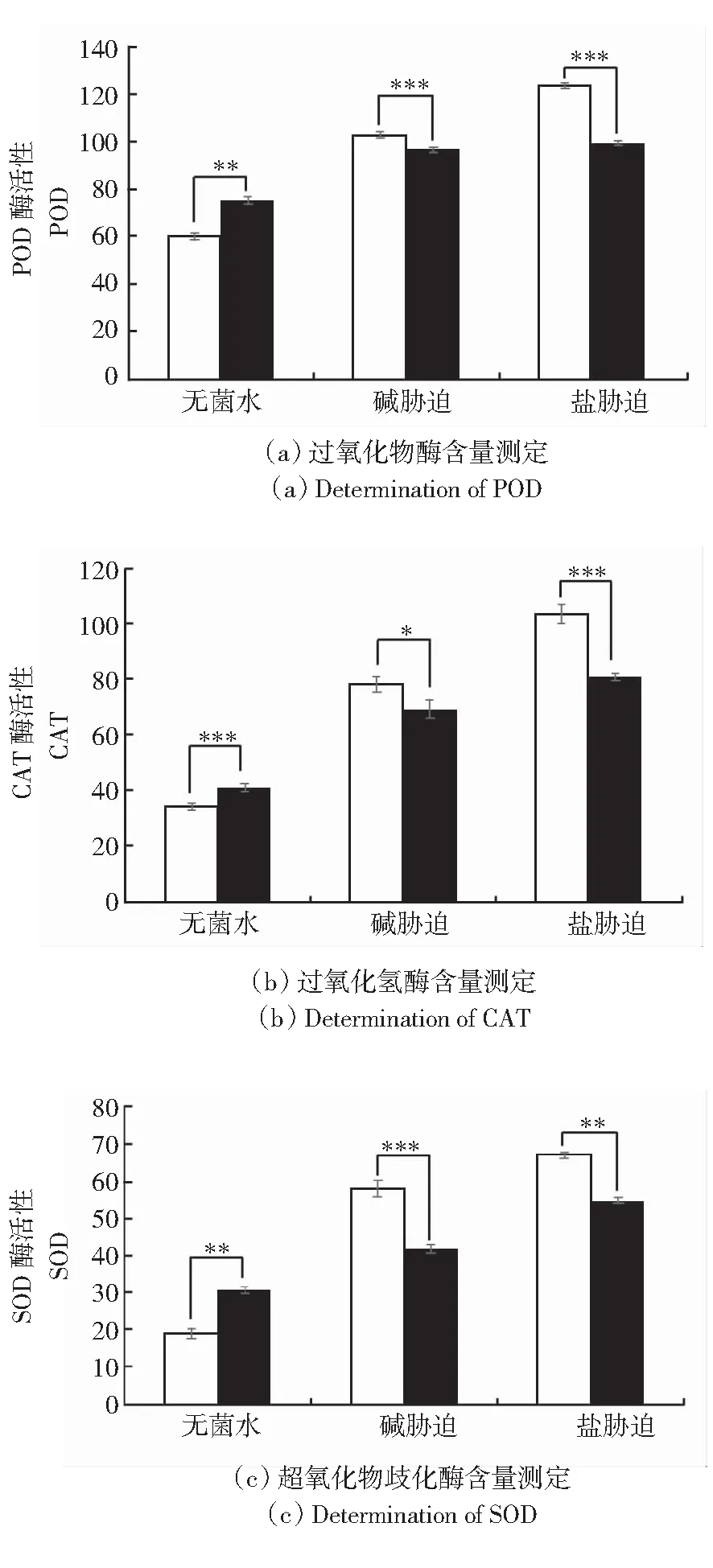
2.5 盐碱胁迫对水稻酶活性的影响
当植物受到盐碱胁迫时,会分泌大量的抗氧化物质保护自身,防止活性氧的积累[13]。如图6所示,在盐碱胁迫下,与对照相比,接菌后水稻叶片内酶含量发生变化。在无菌水条件下,接菌后水稻叶片过氧化物酶(POD)的含量上升38.7%,过氧化氢酶(CAT)的含量上升19.9%,超氧化物歧化酶(SOD)的含量上升20.2%。在盐胁迫条件下,水稻叶片过氧化物酶含量下降17.9%,过氧化氢酶含量下降11.2%,超氧化物歧化酶含量下降5.7%。在碱胁迫条件下,水稻叶片过氧化物酶含量下降27.9%,过氧化氢酶21.7%,超氧化物歧化酶下降19.2%。综上所述,在无菌水条件下,接菌后可使酶的活性提高。在盐碱胁迫条件下,接菌后酶的活性下降。
2.6 水稻叶片光合参数的影响
长枝木霉菌对水稻幼苗光合参数的影响,如图7所示,盐碱胁迫,使水稻的光合参数下降。在盐碱胁迫条件下,接菌后,净光合速率、蒸腾速率、气孔导度、叶片胞间CO2总量均有不同程度的升高。接菌后,在无菌水条件下,水稻叶片光合速率上升13.2%,蒸腾速率上升31.8%,气孔导度上升6.8%,胞间CO2的含量上升22.2%。在盐胁迫条件下,净光合速率含量上升13.8%,蒸腾速率含量上升41.8%,气孔导度含量上升24.6%,胞间CO2含量上升9.4%。在碱胁迫条件下,净光合速率含量上升13.5%,蒸腾速率含量上升17.9%,气孔导度含量上升65.3%,胞间CO2含量上升18.4%。
*表示P<0.05;**表示P<0.01;***表示P<0.001。白色柱表示未接种对照;黑色柱表示接种长枝木霉菌株。

**表示P<0.01; ***表示P<0.001。白色柱表示未接种对照;黑色柱表示接种长枝木霉菌株。
3 讨论
木霉菌是一种有益真菌,普遍存在于土壤、植物根际和植物组织中,是重要的微生物资源[15-16]。木霉菌能够产生一系列的防御反应,提高植物抗病性,同时也能产生植物激素,促进植物生长[16-17]。目前人们对木霉菌的研究主要集中在生物防治方面,而在盐碱条件下,木霉菌促进植物生长展开的研究较少。本研究从松嫩平原盐碱地蒙古柳根部分离出长枝木霉菌,并分析长枝木霉菌对植物的解盐促生能力。发现长枝木霉菌在生物量、生理生化、光合参数等方面增强了水稻幼苗对盐碱胁迫的耐受性,能够促进水稻生长。
本研究中,进行3组6个处理,分别为水、水+菌、盐、盐+菌、碱、碱+菌。以水、盐、碱分别作为每组的对照。在无菌水和盐碱条件下,长枝木霉菌接种水稻种子后,显著提高水稻种子发芽率和发芽势,且在盐碱条件下经长枝木霉菌处理后水稻种子的发芽率接近于无菌水条件下,能够基本消除盐碱胁迫对水稻种子萌发的抑制作用,提高水稻种子的发芽能力。接种长枝木霉菌后水稻地下部分和地上部分的干鲜重都得到了显著的提升,说明长枝木霉菌缓解了盐碱离子对水稻幼苗生长的抑制作用;还发现盐碱胁迫显著降低水稻幼苗中的叶绿素含量,但在接种长枝木霉菌后,水稻幼苗中的叶绿素含量上升。由此可见,长枝木霉菌显著促进水稻幼苗的生长,并帮助其在盐碱胁迫下积累光合产物,为植株自身的生理活动提供营养物质。 Mastouri等[19]研究发现,在非生物胁迫下,哈茨木霉(Trichodermaharzionnum)T22通过减少渗透胁迫和脂质过氧化物的含量,从而缓解氧化性损伤,同时还可以提高种子的萌发率。张树武等[20-21]研究发现,在盐胁迫下,长枝木霉T6菌株处理小麦种子7 d后,小麦种子的发芽率和发芽势提高,其种子胚芽长度的增长率为9%~34%,胚根长度的增长率为9%~31%。同时发现浓度100倍和50倍的深绿木霉发酵液对白三叶幼苗的叶绿素、抗性相关酶活性有较为明显的提升,且不同浓度的发酵液对白三叶草幼苗的影响不同。廉华等[22]发现合适浓度的棘孢木霉菌剂能促进黄瓜幼苗的形态建成和物质积累,也可增加叶片中总叶绿素含量和根系活力等指标。陈伯清等[23]将木霉菌孢子悬浮液作用于番茄幼苗根际后,发现番茄幼苗叶片中叶绿素含量上升,促进番茄光合作用。这些研究与本研究一致。
盐碱胁迫会使细胞内积累大量的活性氧(ROS),氧化损伤细胞内的膜脂、蛋白质等。为了抵御盐碱胁迫对植物的伤害,植物进化出有效的抗氧化系统,防止植物受到氧化损伤。人们常用抗氧化酶来评价植物的氧化还原状态。本研究发现,在无菌水条件下,长枝木霉菌使各抗氧化酶的活性提升,有利于植物体内活性氧的清除。在盐碱胁迫下,长枝木霉菌使各种抗氧化酶的活性下降,说明其有效缓解盐碱离子对幼苗的毒害作用。丙二醛含量反映细胞质膜受损伤的程度。本研究发现,接菌后,丙二醛明显下降,说明受氧化胁迫的幼苗生长得到缓解。李艳娟等[24]通过试验证明哈茨木霉与绿色木霉2种木霉菌混合作用于杉木幼苗,可提高杉木幼苗过氧化物酶、过氧化氢酶活性,降低了丙二醛含量,显著提高幼苗的抗逆性。尹大川等[25]发现绿木霉T43接种樟子松苗木后,过氧化物酶、过氧化氢酶、超氧化物歧化酶的活性显著上升,表现出较高的酶活性,可以提高植物的抗逆性。这些研究结果与本研究一致。
本研究中,在盐碱条件下,水稻幼苗用长枝木霉孢子悬浮液处理后,与对照组相比,光合参数指标得到提升,促进植物光合作用。此结果与他人研究一致,周晓馥等[26]发现哈茨木霉菌液灌根处理黄瓜幼苗,黄瓜植株的总根长、根尖数、分支数明显增加。叶片的净光合速率、气孔导度等明显提高,促进植物的光合作用。陈双臣等[27]发现木霉菌可以提高番茄的光合作用,可提高番茄叶片的净光合速率、气孔导度等指标。本研究发现长枝木霉菌可以提高水稻的光合指标等参数来增加光合作用,与他人的研究一致。说明长枝木霉菌通过提升水稻叶片的光合参数来促进水稻的生长发育。
4 结论
本研究从蒙古柳的侧根中分离出长枝木霉菌,并对其解盐促生能力进行深入研究。在盐碱条件下,长枝木霉菌提高水稻种子发芽率和发芽势,促进水稻种子胚根和胚芽的发育,提高水稻幼苗的株高、根长等各项生物量指标。在生理指标方面,长枝木霉菌使水稻叶片中丙二醛(MDA)、超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)的含量下降。在光合参数方面,接菌后水稻叶片净光合速率、蒸腾速率、气孔导度和胞间CO2浓度都得到了显著的提高。说明长枝木霉菌能减少盐碱胁迫对植物的伤害,增强细胞抗氧化体系,提高水稻对盐碱胁迫的抵御能力。
