黄菖蒲花部特征及繁育习性研究
2023-02-10范少茹符佳豪庄秋荣张永侠王银杰刘清泉田松青原海燕
范少茹,符佳豪,庄秋荣,张永侠,王银杰,张 婷,刘清泉,田松青,原海燕*
(1.江苏省中国科学院植物研究所,江苏 南京 210014;2.苏州农业职业技术学院,江苏 苏州 215008)
鸢尾属(IrisL.)植物是一类具有较高观赏、生态和经济价值的多年生草本植物,全球约300个种,主要分布于亚欧大陆和北美洲的温带地区[1-2]。据中国植物志记载,我国产鸢尾属植物约60种、13个变种及5个变型,占全球资源的1/5多,主要分布于我国西北、西南及东北[3-4]。近年来,随着国内对鸢尾属植物研究与开发利用的逐渐重视,鸢尾属植物在园林应用、生态修复、传统医药、香水和化妆品等行业产生了显著的经济和社会效益[5-7]。
黄菖蒲(IrispseudacorusL.)为鸢尾科鸢尾属无附属物亚属的多年生草本植物,原产于欧洲、北非和西亚等地,目前在我国各地都有引种栽培。黄菖蒲花期为4—5月份,花色黄艳,花姿秀美,观赏价值极高,主要应用于城市绿化以及水体美化和净化[3]。目前对黄菖蒲研究主要集中在水体净化[8-9]、生理抗性[10-11]、杂交育种[12]、种子萌发特性[13]、遗传多样性[14]等方面,对黄菖蒲花部特征及繁育习性等方面的研究还鲜有报道。黄菖蒲作为一类在我国广泛引种栽培的鸢尾属植物,长久以来被应用的黄菖蒲花色均为金黄色,花色单一,因此,了解黄菖蒲生殖生物学相关基本特征对利用遗传育种进行花色改良以进一步丰富黄菖蒲的花色和提高应用价值具有重要的意义。
植物的繁育系统为能够影响到后代遗传组成及适应性的因素总和,主要影响因素包括花器官特征及寿命、开花动态、传粉者的类型及繁育习性等,其中植物的花器官特征与繁育习性是其主要影响因素[15-16]。目前国内外已有相关研究的鸢尾属植物有鸢尾(Iristectorum)[17]、溪荪(Irissanguinea)[18]、马蔺(Irislactea)[19]、喜盐鸢尾(Irishalophila)[20]、小花鸢尾(Irisspeculatrix)[21]、膜苞鸢尾(Irisscariosa)[22]等,为鸢尾属植物的栽培及育种提供了理论基础。黄菖蒲花部特征与繁育习性在国内外尚未见报道,为此,该研究拟通过观察记录与授粉试验等方法,对黄菖蒲的花器官特征、开花动态、花粉形态及活力、柱头可授性、繁育习性与访花昆虫进行了观察与研究,为黄菖蒲生殖生物学特征提供依据,同时为后期杂交育种、品种改良与资源利用提供参考。
1 材料与方法
1.1 研究材料
以江苏省中国科学院植物研究所鸢尾种质资源圃中的黄菖蒲居群为研究对象,该居群于1994年自新疆乌鲁木齐植物园引种,种植20余年,面积约50 m2,共有黄菖蒲200余株。经江苏省中国科学院植物研究所黄苏珍研究员鉴定,为鸢尾科植物黄菖蒲(Irispseudacorus)。研究地处北纬32°05′79″,东经118°83′61″,海拔48 m,属亚热带季风气候,年平均气温14.7℃,相对湿度76%,年平均降水量1 000.4 mm,年平均日照为1 933 h,无霜期237 d。2022年4—5月,随机选择生长健壮、无病虫害的黄菖蒲植株进行观测。
1.2 试验设计
1.2.1生殖生物学性状观察 将研究材料中所选居群设置4个面积为3 m×3 m的区组。花部特征性状观察为盛花期时每个区组测量3朵花,共12朵花;花粉活力及柱头可授性测定为每天随机取3朵花测定,共测定3天;花粉/胚珠比共测定20朵花,每个区组测定5朵;杂交指数共测定40朵花,每个区组10朵。
1.2.2授粉实验 选取即将开放的花蕾进行授粉试验,设置T1—T9共9个处理(包括对照),每个处理选择20朵即将开放的花蕾进行授粉。T1处理为花蕾直接套袋;T2处理为花蕾去雄后套袋;T3处理为花蕾去雄后不套袋,自然授粉;T4处理为花蕾去雄后套袋,次日9∶00—11∶00人工异株授粉;T5处理组为花蕾套袋,次日9∶00—11∶00人工同株同花授粉;T6处理组为花蕾去雄后套袋,次日9∶00—10∶00人工同株异花授粉;T7处理组为花蕾去除外轮花被片后不套袋,自然授粉;T8处理组为花蕾去除全部花被片后不套袋,自然授粉;对照组(CK)为花蕾自然授粉。在授粉14 d后统计各组的种子数、坐果率及结籽率,计算公式为:种子数=(某组所有种子数/该组果实数)×100%;坐果率=(某组的果实数/处理个数)×100%;结籽率=(某组所有果实结籽率总和/该组的果实数)×100%。根据授粉试验所得结籽率计算黄菖蒲在自然状态下是否存在花粉限制和繁殖保障。
1.3 测定方法
1.3.1开花物候及花部特征观察 对区组内黄菖蒲植株的花仔细观察,记录该居群的花期、单株花期(单株上第1朵花开花到最后1朵花闭合的天数)、单花花期(一朵花从开放到闭合的时间)及开花进程(始花期为区组内10%的植株同时开花,盛花期为区组内超过50%的植株同时开花,末花期为区组内仅剩10%以内的植株仍在开花)[23]。使用游标卡尺测量开花当天的花直径、花纵径、垂瓣长与宽、旗瓣长与宽、柱头长与宽、花丝长及花药长。
1.3.2花粉形态检测 在黄菖蒲盛花期收集自然开裂花药中的花粉,烘箱40℃低温烘干,取干燥花粉抖落在贴有导电胶的样品台上,使用洗耳球吹净多余的花粉,将样品台放至在离子溅射镀膜仪(日本EIKO公司)中喷金镀膜,之后用Hitachi S 3000 N扫描电镜(日本Hitachi公司)进行观察。选取代表性的视野观察花粉粒及其外壁纹饰并进行拍摄,对花粉的极轴长和赤道轴长进行测量,以极轴长/赤道轴长(P/E)表示花粉粒形状:P/E>1.6为长椭球形;P/E处于1.3~1.6之间为椭球形;P/E<1.3为类球形[24]。
1.3.3花粉活力与柱头可授性检测 选取未开放的花蕾挂牌,从开花前一天开始采集,至花朵萎蔫,每天采集3朵花的花粉与柱头,测量花粉及柱头可授性。使用0.5%的TTC(2,3,5-氯化三苯基四氮唑)溶液测定不同时期的花粉活力。具体操作步骤如下:取少量花粉置于离心管上,加200 μL TTC溶液,搅拌均匀,避光染色20 min后制片。在光学显微镜低倍镜下观察,每个片子取3个视野,每个视野至少观察50个花粉粒,具有较强生活力的花粉粒呈深红色,微弱活力的花粉粒呈淡红色,无活力或不育的花粉为无色。花粉活力=(视野内染色的花粉粒数/视野内花粉总数)×100%。
采集到的柱头浸入滴有联苯胺-过氧化氢反应液(1%联苯胺∶3%过氧化氢∶水=4∶11∶22,体积比)的凹面载玻片中,观察并拍照记录,柱头周围呈棕色并且有大量气泡产生表明其具有可授性,反之则无[25]。
1.3.4花粉/胚珠比(Pollen/Ovule,P/O)与杂交指数(Outcrossing index,OCI)检测 随机选取花药尚未开裂的黄菖蒲花蕾,每个花蕾随机选取1枚雄蕊用1 mol·L-1HCl于60℃水浴下水解去除药壁后,蒸馏水定容至10 mL,振荡器上震荡制成花粉粒悬浮液,吸取花粉粒悬浮液10 μL置于载玻片上,在显微镜下观察并统计花粉数,取其平均值X,则每朵花的花粉数为3×1 000X;将随机选取到的黄菖蒲花蕾子房在解剖镜下用解剖刀剖开心皮,观测并记录其胚珠数,取其平均值。计算出花粉/胚珠比(P/O)判定其繁育习性。花粉/胚珠比(P/O)=平均每朵花的花粉数/平均每朵花的胚珠数。P/O为2.7~5.4时,繁育习性为闭花受精;P/O为18.1~39.0时,繁育习性为专性自交;P/O为31.9~396.0时,繁育习性为兼性自交;P/O为244.7~2 588.0时,繁育习性为兼性异交;P/O为2 108.0~195 525.0时繁育习性为专性异交[26]。
杂交指数OCI值为以下三者相加:(1)花朵直径≤1 mm记为0,1~2 mm记为1,2.1~6 mm记为2,>6 mm记为3;(2)雌雄蕊同时成熟或雌蕊先熟记为0,雄蕊先熟记为1;(3)雌蕊与雄蕊为同一高度记为0,空间分离记为1。当OCI=0时,繁育习性为闭花受精;OCI=1时,繁育习性为专性自交;OCI=2时,繁育习性为兼性自交;OCI=3时,繁育习性为自交亲和,有时需要传粉者;OCI=4时,繁育习性为部分自交亲和,异交,需要传粉者[27]。
1.3.5花粉限制及繁殖保障 花粉限制(Pollen limitation,PL)是落在柱头上的花粉数量不足导致的结实受限现象[28],SS(Seed setting rate)为结籽率,以杂交授粉(Cross-pollination,CP)和自然授粉(Open-pollination,OP)结籽率的差值评估花粉限制:PL=SSCP-SSOP;若PL<0,说明不存在花粉限制;若PL>0,且在统计学上差异显著(P<0.05),则存在花粉限制。
繁殖保障(Reproductive assurance,RA)是在缺少传粉者或配子的情况下以自花授粉来保证繁衍的一种现象[29],以去雄后自然授粉(Pollination after emasculation,PE)和自然授粉结籽率的比值来判断繁殖保障的有无及高低[30]:RA=1-SSPE/SSOP;若RA<0,说明不存在繁殖保障;RA>0,存在繁殖保障现象。
1.3.6访花昆虫行为观察 在黄菖蒲盛花期,每天9∶00至17∶00观察样方内的所有访花昆虫,记录观察到的访花昆虫的种类、数量、单次访花时间并拍照记录不同访花昆虫。根据昆虫在访花过程中身上是否携带花粉来判断是否为潜在的传粉者。
1.4 统计分析
使用SPSS 26.0(IBM Corp.,Chicago,IL)通过单因素ANOVA和独立样本t检验进行数据分析;图片使用Photoshop 2020软件处理。
2 结果与分析
2.1 黄菖蒲花部特征
观察结果表明,黄菖蒲苞片绿色,3~4枚,花为两性花,花被片两轮排列,辐射对称,子房三棱状柱形,心皮三枚,中轴胎座。黄菖蒲花平均直径10.88 cm,平均纵径9.76 cm;外花被3枚,花色黄色并伴随黑褐色条纹,长6.87 cm,宽3.66 cm,先端卵圆形或倒卵形,开放时自然下垂(图1A),转折处与柱头接触,基部狭楔形,两侧向上翻折(图1C),宽度略小于柱头;内花被裂片较小,3枚,黄色,倒披针形,长2.45 cm,宽0.57 cm(图1B);花柱分枝3个,淡黄色,略弯曲,与外花被裂片形成可让授粉昆虫进入的空间,长3.71 cm,宽1.23 cm,顶端裂片半圆形,边缘呈稀疏牙齿状,柱头成熟后向上翻折(图1C);雄蕊长2.75 cm,花丝细长,花药紫色,向外开裂;花药长1.19 cm,位于柱头下方,花粉黄色(图1A)。

图1 黄菖蒲花部特征
2.2 黄菖蒲开花物候
黄菖蒲4月下旬现蕾,始花期为04/20—04/27,盛花期为04/28—05/10,末花期为05/11—05/20,总体花期约为30 d,其中始花期7 d,盛花期13 d,末花期9 d,单花花期2~3 d,单株花期6~8 d。花朵晴天时早上8∶00~10∶00陆续开放,单花开放时,黄色的花蕾开始伸长(图2A),接下来各个花器官绽开(图2B),外花被片缓慢舒展并下垂,内花被片保持直立舒展的状态,随后柱头慢慢弯曲向外花被片靠近(图2C)。花朵开放的第2 d,外花被片开始萎蔫并向内卷曲(图2D),花被片及柱头开始向内抱合(图2E),直至最后所有花被片及柱头萎缩卷曲在一起(图2F)。黄菖蒲在开花过程中,雄蕊始终短于柱头,开花前1 d,雄蕊已经成熟;开花第1 d,柱头具有可授性,雄蕊散出花粉,大多数花粉在散出几小时后落在外花被片上;开花第2 d,花粉散完,外花被萎蔫卷曲。

图2 黄菖蒲花朵开放动态
2.3 花粉形态、活力及柱头可授性
扫描电镜观察结果表明,黄菖蒲的花粉形状为长椭球形,萌发沟数量为一个(图3A),外壁纹饰为显著粗网状,网眼内为凹凸不平的凸起(图3C),极轴长约130.23 μm,赤道轴长42.35 μm,极轴长和赤道轴长之比(P/E)约为3.08(图3B);花粉活力检测结果表明,黄菖蒲花粉在开花前一天花粉未散出时就具有很强的活力(91.71±5.81)%,此时柱头还没有可授性(图4A,图5A);开花当天的花粉活力达到最高(92.62±2.75)%,柱头可授性也最强(图4B,图5B),此时为授粉的最佳时间;开花第二天,花粉活力下降至(51.50±12.84)%,柱头可授性也显著降低(图4C,图5C)。总体来看,黄菖蒲的花粉活力及柱头可授性呈现先升高,后降低的趋势(表1)。

图3 黄菖蒲花粉形态、外壁纹饰

图4 黄菖蒲花粉活力

图5 黄菖蒲柱头可授性
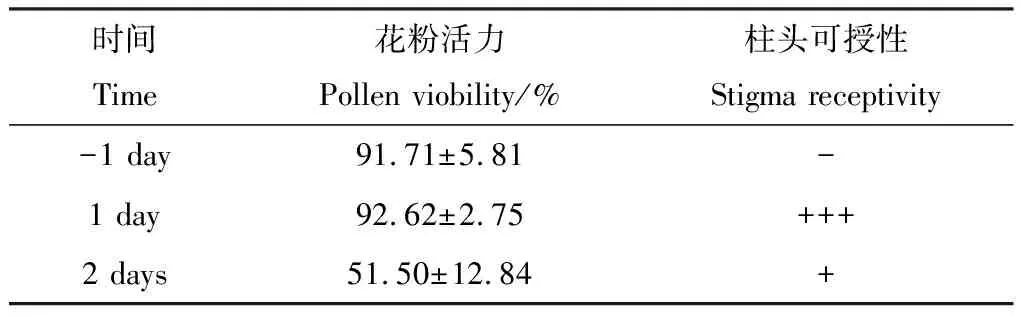
表1 黄菖蒲花粉活力及柱头可授性
2.4 花粉/胚珠比(P/O)及杂交指数OCI值的估算
黄菖蒲单花平均总花粉数为47 700.0,平均胚珠数为125.4,花粉/胚珠比(P/O)为380.4,判定黄菖蒲的繁育习性为兼性自交或兼性异交;黄菖蒲单花直径为107.90 mm,记为“3”;雄蕊先熟,记为“1”;雌蕊与花药在空间上分离,记为“1”,黄菖蒲的OCI值为5,判定其繁育习性为以异交为主,部分自交亲和,需要传粉者。
2.5 繁育习性
2.5.1授粉试验结果 如表2所示,T1组种子数及结籽率为0;T2组坐果率为100%,结籽率为0;T3组的种子数与结籽率近乎为0;T4组及T6组的种子数与结籽率显著高于对照(P<0.05);T5组种子数与结籽率与对照差异不显著;T8组的种子数与结籽率均为0。以上结果表明:黄菖蒲自交亲和,去雄套袋后子房自然膨大,但胚珠并没有发育,因此不存在无融合生殖的现象,去掉外轮花被片后不结实。

表2 黄菖蒲授粉试验结果
2.5.2花粉限制及繁殖保障 根据授粉试验的结籽率来计算黄菖蒲在该引种地自然状态下的花粉限制及繁殖保障程度。结果表明,CK组与T4组的结实率相差较大,与T5组结籽率相近,黄菖蒲在该引种地存在较高的花粉限制(PL=0.22,P<0.05)(图6A,图6B)及繁殖保障(RA=0.88)(图6C)现象。
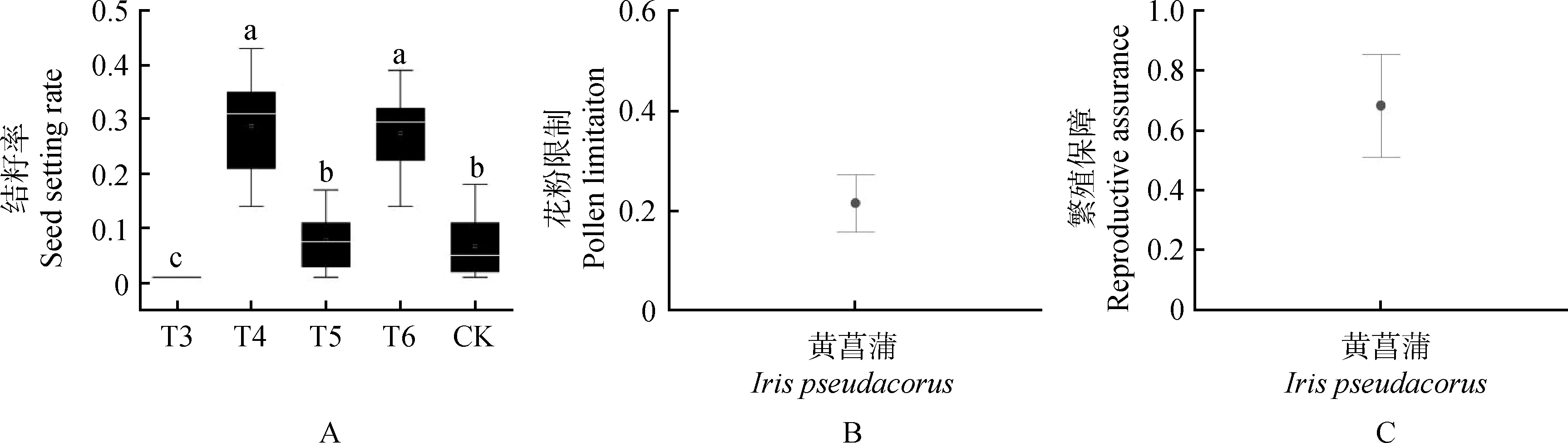
图6 黄菖蒲花粉限制及繁殖保障
2.6 访花昆虫种类及访花行为
黄菖蒲的主要访花昆虫有黑纹食蚜蝇(Episyrphusbalteatus)、淡脉隧蜂(Lasioglossum)与蓟马(ThripsvulgatissimusHaliday),三者的访花时间分别为30 s~1 min,5 s~13 s,1 h~6 h。黄菖蒲生境为湖畔或沼泽,叶片上常常散生有蓟马的卵,孵化后成虫就在柱头下取食和活动,虽然蓟马的访花时间远远长于其他两者,但是由于蓟马行动距离短、访花数少、花粉携带量少[31],判断黑纹食蚜蝇为潜在的传粉昆虫。访花过程中,黑纹食蚜蝇会停落在垂瓣上取食上面的花粉,跟随蜜导的指引进入柱头与外花被之间的访花通道,因为柱头的可授面与外花被形成了访花通道的半闭合入口,因此黑纹食蚜蝇的身体会与柱头的授粉处接触,传递花粉,随着黑纹食蚜蝇进入传粉通道,雄蕊上的花粉会黏在黑纹食蚜蝇的头部与背部(图6 A-D)。淡脉隧蜂采集花蜜时直接在柱头基部或花冠筒上伸入口器吸取花蜜;采集花粉时并不直接进入传粉通道,而是在柱头两侧的缝隙中探入身体采集花粉,且在观察过程中,中华蜜蜂及淡脉隧蜂的数量较少,因此判断二者并不是黄菖蒲的有效传粉昆虫。在观测过程中还发现了少量的蜻蜓与蜘蛛,可能是访花昆虫的捕食者。
3 讨论
植物的花部构造往往与传粉者相适应[23],而鸢尾属中不同物种的传粉者与其柱头与垂瓣之间传粉通道的大小具有相关性[20]。例如鸢尾的花瓣结构较为松散,花药隐藏于柱头之下,因此适合黄胸木蜂(XylocopaappendiculataSmith)等大型昆虫传粉[17];喜盐鸢尾的花冠紧凑,传粉通道狭窄,因此有效传粉昆虫只有条蜂(Anthophorasp.)[20]。而本研究对象黄菖蒲柱头弯曲,传粉通道的入口狭窄,因此授粉昆虫黑纹食蚜蝇体型较为扁平。

图7 黄菖蒲主要访花昆虫
花蜜与花粉是植物吸引昆虫的两种主要手段[32],鸢尾花蜜腺为部分心皮不愈合,在子房室内反卷,表皮细胞分化为产蜜组织发育而成[33]。本研究发现黄菖蒲的蜜腺位于心皮边缘,花蜜由孔道流至花瓣基部,与研究结果相吻合。除此之外,鸢尾垂瓣同样具有吸引及引导授粉昆虫的作用,研究发现,垂瓣在鸢尾花授粉过程中起着至关重要的作用,如果去除垂瓣,授粉将不能进行[34-35],本研究结果也证实了这一点。黄菖蒲去除外花被或全部花被片后,种子数与结籽率为0,说明了外花被片在黄菖蒲吸引授粉者中起着决定性的作用。此外,黄菖蒲外花被片为黄色,对昆虫有着天生的吸引力;外花被片上还有从基部延伸的棕黑色条纹蜜导,蜜导是一类对授粉昆虫有着吸引和指示作用的花图案或花结构[20];黄菖蒲的外花被片还有承载授粉昆虫,与柱头构成授粉通道的作用;黄菖蒲花药开裂后花粉也多数散落在外花被片上吸引昆虫。因此,黄菖蒲的外花被片的结构与功能已完全特化,是用来吸引授粉昆虫的主要手段。
本研究授粉试验结果中T1组(花蕾直接套袋)坐果率及结籽率为0%;T2组(花蕾去雄后套袋)坐果率、结实率为100%和0%;T3组(花蕾去雄后不套袋)坐果率及结籽率为10%和0.8%,推测T2组花蕾去雄后套袋的坐果率为100%而结籽率却为0%的原因可能是花蕾去雄套袋后温度和湿度变化协同作用触发了黄菖蒲子房的膨大,但由于没有花粉完成受精过程,胚珠并没有膨大,也说明黄菖蒲不存在无融合生殖现象。有研究者在对马铃薯(SolanumtuberosumL.)的孤雌生殖研究中也发现,环境的温度与湿度是影响马铃薯坐果率和结籽率的重要因素[36]。此外,自然界中两性花的结实率通常受花粉限制、资源限制、传粉昆虫限制等影响,而自交亲和可能是植物应对传粉限制的适应机制,因为专性异交的植物过于依赖传粉者,当传粉者活动较弱时就会面临较高的灭绝风险,而专性自交可能会导致近交衰退,因此,自然界大多数植物的繁育习性为混交[37-42]。在本研究中,黄菖蒲的繁育习性为以异交为主,部分自交亲和,但授粉实验发现其受到了较高的花粉限制,繁殖保障现象明显,推测是由于授粉昆虫的缺少导致。此外,黄菖蒲与多数鸢尾属植物相同也可以通过根茎无性繁殖作为一种生殖补偿机制来保证繁衍。但是,长期的自交会降低物种的遗传多样性,进而引发近交衰退[43]或种子折损[44-45],因此近交带来的繁育保障优势需要与其带来的近交衰退相抵消或权衡,我们推测本研究所用黄菖蒲资源的繁育习性是与引种地气候、昆虫等相互适应的结果。因此,对于引种驯化的黄菖蒲应该种植于传粉昆虫较多的水边,避免近交衰退,保持物种多样性。
4 结论
黄菖蒲4—5月开花,雌雄异熟、雌雄异位,单朵花期2~3天,单株花期6~8天。黄菖蒲为典型的虫媒花,繁育习性以异交为主,部分自交亲和,需要传粉者,自然状态下存在花粉限制,繁殖保障明显,不存在无融合生殖现象,黑纹食蚜蝇为潜在传粉者。
