双相情感障碍与抑郁症DLPFC脑区转录组差异基因表达数据挖掘分析
2023-02-04刘芳杨萍王枭冶肖剑英石璐
刘芳,杨萍,王枭冶,肖剑英,石璐
双相情感障碍(bipolar disorder,BD)与抑郁症(depression)是最常见的慢性、反复发作的情感类精神疾病[1,2]。流行病学显示我国BD加权年患病率为0.5%,大多数国家抑郁的终身患病率在8%~12%之间[3,4],给家庭及社会带来了沉重的负担。然而目前BD与抑郁症发病机制仍处在探索阶段。在临床实践中,BD患者处于抑郁期时,有部分患者容易被确诊为单向抑郁[5]。BD与抑郁症具有共同的遗传危险因素[6]。
近年来,随着影像学发展,很多学者认为背外侧前额叶皮质(dorsal lateral prefrontal cortex,DLPFC)在负性认知、情绪加工的过程中可能起重要作用[7,8]。BD和抑郁症发病具有相似或者重叠的脑区,DLPFC是大脑的执行网络的重要组成部分,与多种认知控制过程有关[9]。DLPFC在两种疾病的病理和生理中起关键作用[10,11],BD患者和抑郁患者在处理情绪面孔时导致边缘系统激活,BD患者在识别情绪面孔时,DLPFC活性下降[12]。抑郁患者DLPFC的局部脑血流量灌注较低,抑郁的严重程度与DLPFC的灰质体积呈负相关[13]。然而不管是结构影像还是功能影像都难以确定DLPFC活性改变是两种疾病的素质指标还是状态指标。最近已有对BD与抑郁症的DLPFC的测序方面研究,故本研究将基于数据挖掘分析BD与抑郁症DLPFC转录组差异性分子表达,分析其与免疫、代谢的关系,为BD、抑郁症的潜在病因学研究和临床治疗提供新方向。
1 资料与方法
1.1 一般资料
检索Gene Expression Omnibus(GEO)数据库,检索时间为2016年1月1日至2021年12月31日,检索词为“bipolar disorder(BD)”、“depression”、“dorsal lateral prefrontal cortex(DLPFC)”、“Peripheral blood”、“sequencing”,总共检索到8组相关数据集。数据纳入标准:研究对象为BD、抑郁症患者;样本需进行DLPFC组织转录组测序(RNA-seq)或进行外周血RNA-seq并具备对照组织样本的测序;计算了不同个体的基因表达水平;具备完整的临床信息。排除标准:合并神经系统疾病史或重大躯体疾病史的测序研究;有酒精或药物滥用或依赖史的测序研究;使用神经兴奋性药物的测序研究。经过筛选后确定GSE78936、GSE101521、GSE124326数据集为本研究可用数据,共纳入552例转录组数据。其中包括6例对照和7例BD患者(GSE78936)、29例对照和30例抑郁症患者的DLPFC转录组数据(GSE101521);240例健康对照和240例BD患者的外周血转录组数据(GSE124326)。
1.2 方法
1.2.1 分组 总共纳入552例,按检索式为“BD+DLPFC sequencing”,GSE78936纳入对照组6例,BD组7例;按“MDD depression+DLPFCsequencing”检索式,GSE101521纳入对照组29例和抑郁症组30例患者;按检索式为“BD+Peripheral blood+sequencing”,GSE124326纳入对照组240例,BD组240例。
1.2.2 差异表达基因计算及统计学处理 根据以上分组数据,从基因表达数据集GSE78936患者寻找BD组与对照组DLPFC脑区的差异表达基因,从基因表达数据集GSE101521寻找抑郁组与对照组DLPFC脑区的差异表达基因;从基因表达数据集GSE124326寻找外周血基因差异表达。使用R语言中的DESeq2包(版本号为1.26.1)鉴定BD、抑郁患者DLPFC脑区相较于对照组差异表达的基因,用火山图展示差异基因的情况,用热图展示上下调top50的差异表达基因(基于gene level),DEgenes代表患者组与对照组中表达水平显著差异的基因,Unchanged代表患者组与正常组中表达水平差异不显著的基因。FoldChange为患者组中平均表达水平/对照组中平均表达水平,差异基因阈值设置FoldChange≥1.5,且P≤0.05。
1.2.3 差异基因功能注释 将BD、抑郁症组较对照组计算出的差异表达基因进行功能富集分析,将外周血差异表达基因进行功能富集分析。通过使用R中的clusterProfiler包(版本号为3.14.3)采用功能注释基因富集的生物学过程(Biological Process,BP)[9],从基因学角度对BD、抑郁发病原因进行探索。
1.2.4 基因交并集分析 对抑郁症患者和BD患者的差异基因进行交并集分析,统计BD和抑郁患者中差异表达基因与代谢和免疫相关基因的交集(免疫相关基因列表从ImmPort数据库下载,共包括2 498个免疫相关基因;代谢相关基因列表从KEGG下载,共包括1 660个代谢相关基因)。
1.2.5 BD患者代谢相关差异性基因验证 从GSE124326纳入BD外周血转录组数据,验证BD是否与代谢相关;差异基因鉴定及功能注释方法同上,使用GSVA包(版本号为1.34.0)计算各代谢通路在BD患者样本中的富集分数。
2 结果
2.1 BD和抑郁症患者DLPFC脑区的差异表达基因
通过DESeq2算法计算后发现与对照组相比,BD和抑郁患者DLPFC脑区具有不同的差异表达基因。BD患者DLPFC脑区显著高表达基因总共有384个,其中 排 名 前 十:IL1RL1、SERPINA3、NTF3、FPR1、ATP5MGL、OR10A4、PLN、IL18R1、EXO1、ANGPTL4;显著低表达基因总共有117个,其中排名前十:VNN1、LILRA4、GLYATL2、IGF2、ADAMTS14、SGK2、NGFR、FOXD4L6、CEL、ADAT3。抑郁患者DLPFC脑区显著高表达基因总计165个,其中排名前十:MTRNR2L8、CTAG1B、DNASE1L3、HLA-DRB6、CYP4B1、MIR3178、LINC02010、FAM47C、MIR4730、MIR30C1,显著低表达基因总计316个,其中排名前十:HSPA6、CXCL8、IL6、MMP8、SERPINH1、CXCL10、CSF3、IL1B、CCL2、LBP,见图1、2。
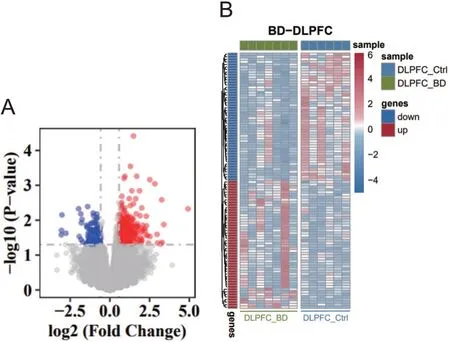
图1 BD组与对照组DLPFC脑区差异表达基因火山图及热图
2.2 BD、抑郁的DLPFC脑区差异表达基因富集分析
通过GO功能分析,得到BD、抑郁的DLPFC脑区差异表达基因BP富集结果。BD患者DLPFC脑区中上调基因富集在mRNA分解代谢、蛋白质定位、氧化磷酸化、ATP代谢、核糖核酸分解代谢等物质和能量分解代谢相关通路上面;下调基因富集到有机酸/核苷酸、磷酸核苷分解代谢和髓鞘形成、轴突、神经元等神经系统相关通路上面。抑郁患者DLPFC脑区中上调基因富集在髓鞘形成、神经元、中枢神经系统、少突细胞等神经相关通路上面;下调基因富集在细胞趋化作用、白细胞游走、粒细胞迁移等免疫炎症相关通路上面,并且这些下调基因与白介素1引起的细胞反应蛋白显著富集,见图3、4。
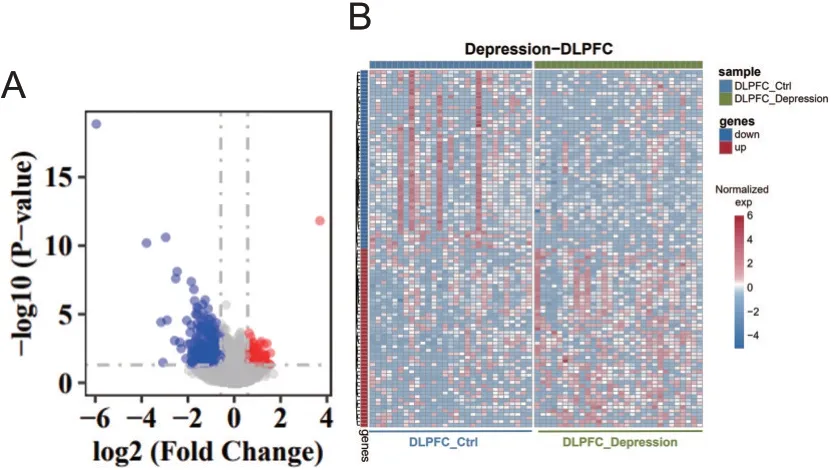
图2 抑郁症组与对照组DLPFC脑区差异表达基因火山图及热图

图3 BD患者DLPFC脑区差异表达基因富集图
2.3 基因交集分析
对BD和抑郁症组DLPFC脑区差异基因进行交集分析,发现2组间有15个基因交集。接着分析2组差异表达基因与代谢和免疫相关基因的交集显示,抑郁患者差异基因中有74个免疫相关基因和20个代谢相关基因;而BD患者差异基因中有42个免疫相关基因和51个代谢相关基因,见图5。
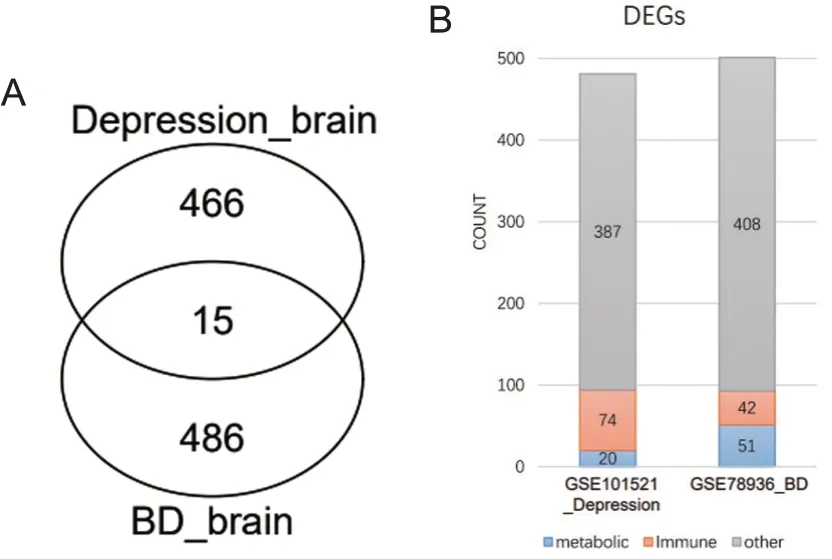
图5 BD和抑郁患者基因交集图
2.4 BD与代谢相关的差异表达基因富集
对BD患者的外周血转录组数据进行基因表达差异分析,并对差异表达基因进行GO富集分析,结果显示,BD患者组中上调表达基因富集到氧气运输、过氧化氢分解代谢、活性氧代谢、一氧化氮代谢等细胞能量代谢相关通路;下调表达基因则富集到膜电位调节、突触信号传递等神经活动相关通路,见图6。
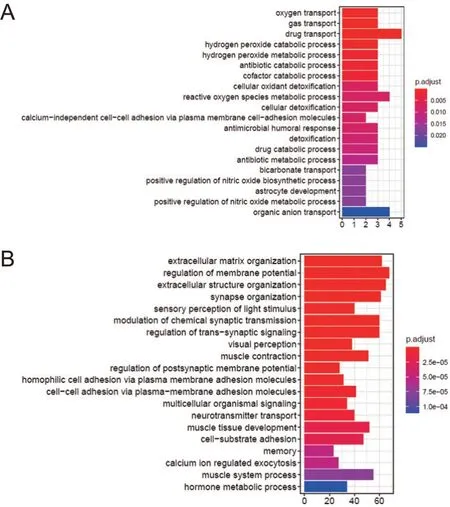
图6 BD与代谢相关的差异表达基因富集图
2.5 BD患者DLPFC脑区、外周血与代谢相关差异性基因分析
比较BD患者脑区与外周血差异基因,结果发现有51个在BD患者DLPFC脑区中显著差异表达的代谢相关基因,在BD患者外周血中也呈现出表达显著差异。基于BD患者DLPFC脑区数据对其进行代谢通路富集分析,结果显示在BD患者中富集程度更高的通路包括脂肪酸生物合成、氧化磷酸化及亚油酸、丙酮酸代谢等多条分解供给能量通路。而健康对照中更富集的通路包括多糖降解、各种糖类、不饱和脂肪酸、类固醇等生物合成通路,见图7。通过BD外周血转录组数据对上述代谢通路富集结果进行进一步验证,结果显示脂肪酸生物合成、泛醌等萜醌生物合成、硒化合物代谢、其他聚糖降解、其他类型的O.聚糖生物合成等通路在BD外周血与健康对照外周血中依然存在富集差异,见图8。
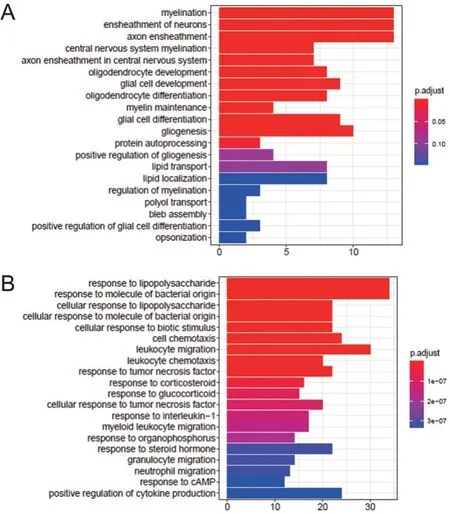
图4 抑郁患者DLPFC脑区差异表达基因富集图
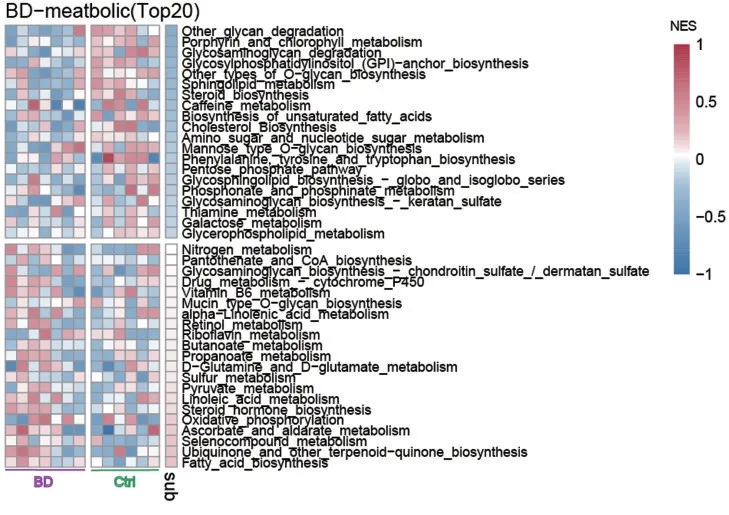
图7 BD患者代谢特征分析(排名前20)
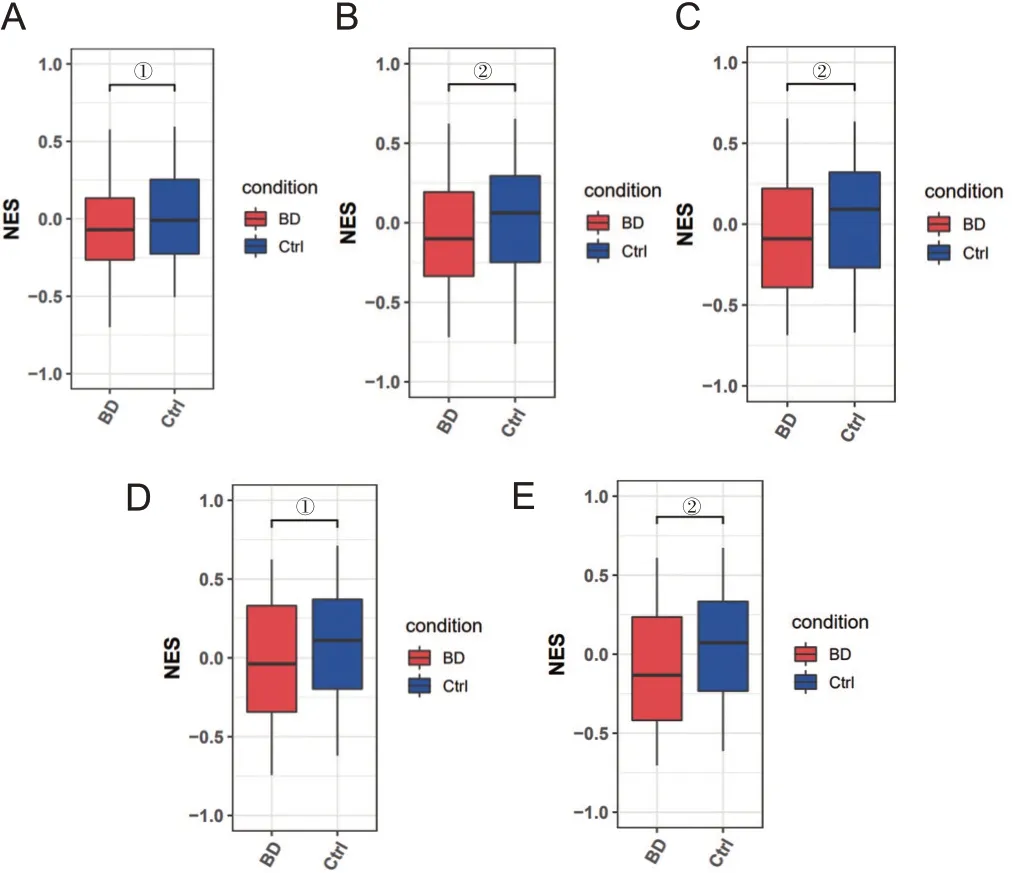
图8 BD外周血转录组数据验证代谢通路富集结果(排名前5)
3 讨论
DLPFC作为额顶网络(fronto-parietal network,FPN)的重要节点,在BD、抑郁症等精神疾病的发生发展机制中起重要作用[14,15]。以往均围绕功能影像学进行研究,鲜少从分子水平进行探讨。本研究基于数据挖掘对BD、抑郁的DLPFC脑区差异表达基因进行功能富集分析,发现BD、抑郁患者中可能会发生神经系统相关功能基因的异常,证实两者发病具有相似或重叠的脑区。本研究从分子角度,发现BD中多发生基因下调,而抑郁多发生基因表达量上调。对两者差异表达基因交并集分析发现两者只有15个交集,说明BD患者和抑郁患者呈现不一样的分子异常。此外,抑郁患者中差异表达基因功能富集与免疫相关,例如差异基因中CXCL8、IL6、IL1B、CXCL10可能通过诱发免疫炎症反应与抑郁发病相关[16-18]。而BD患者则主要富集于代谢通路相关,例如ATP5MGL(ATP Synthase Membrane Subunit G Like)是一个蛋白质编码基因,其相关途径包括嘌呤核苷酸从头合成,PLIN参与脂质代谢,其蛋白水平与TG储存水平相关,能促进脂肪沉积[19]。故抑郁患者可能免疫功能异常为主,BD患者主要表现为代谢异常。以上差别可能与BD表现情绪调控障碍,DLPFC可能参与BD情绪加工和调节过程的神经环路[20],而抑郁的产生与前额叶认知控制不足,不能抑制大脑对负性情绪信息进行偏向加工有关[21]。
抑郁患者中DLPFC脑区多见于免疫相关基因异常,这与本课题组前期研究中的抑郁与免疫反应紊乱紧密相关结果一致[22]。BD患者中代谢相关基因表达异常,可见代谢系统在BD的发生和发展中起到重要作用,代谢相关基因可能参与BD的调控过程。因此本研究进一步下载BD患者外周血转录组数据进行分析,发现BD患者脑区差异基因富集结果一致,进一步证明BD患者除了神经系统的分子异常之外,也表现出代谢系统紊乱。且51个在BD患者中DLPFC脑区中差异表达的代谢相关基因在BD患者血液与正常对照血液中表达有显著差异,因此,差异表达的代谢相关基因可能参与BD患者的发病调控。从代谢特征上分析,BD患者与对照组比代谢系统表现紊乱,主要表现为生物合成通路富集减弱,分解供给能量通路富集增加。既往研究多关注在BD抗精神病治疗相关糖脂代谢紊乱,鲜少对BD代谢进行研究。最近Fürtjes等[23]认为BD患者可能存在心脏代谢异常,与BD过早死亡密切相关。本研究代谢通路排名前五:脂肪酸生物合成、泛醌等萜醌生物合成、硒化合物代谢、其他聚糖降解、其他类型的O.聚糖生物合成,且其他聚糖降解、其他类型的O.聚糖生物合成的富集相对趋势与BD脑区结果一致,提示应关注BD代谢紊乱相关的问题。
因BD、抑郁症脑组织难获取性,本研究仅针对公共数据库中有限的DLPFC脑区测序数据进行挖掘分析,可能会忽略与其他脑区的联系性与差异性比较,且难以进行验证,有待进一步探索。
