不同干旱胁迫及亚精胺复水对南方红豆杉生理特性的影响
2023-02-03温家康邢长杰王亚萍李文杨
温家康,邢长杰,王亚萍,杨 乐,郑 伟,李文杨
(1.信阳农林学院林学院信阳市林木遗传育种重点实验室,河南信阳 464000;2.新疆农业大学林学与风景园林学院新疆教育厅干旱区林业生态与产业技术重点实验室,乌鲁木齐 830052)
0 引言
南方红豆杉(Tɑxus chinensisvar.mɑirei),也叫紫杉,属于紫杉科(Taxaceae),红豆杉属植物(Tɑxus),主要分布于中国南方,其植物中提取的紫杉醇是公认的珍贵抗癌药物,具有极高的利用价值和研究潜力[1,2]。近年来,自然灾害突发频率增大,在各种自然灾害中,旱灾具有持续时间长、影响范围广、频率高等特征,给幼苗期的红豆杉带来极大的危害[3,4]。据报道,干旱对植物的影响是全方位的,往往能够导致细胞结构发生改变,从而对植物的光合作用、生理活性产生影响,导致植物生长受限,严重时甚至会导致植物死亡[5]。利用外源物质提高植物抗逆性是简便、有效、可行的方法之一[6]。
多胺(PAs)属于在植物体广泛存在的一类脂肪族含氮碱化合物,是植物内源性成长调节剂之一,主要有腐胺(Put)、精胺(Spm)和亚精胺(Spd)等[7,8]。由于亚精胺(Spd)的多聚阳离子性能最好,在逆境胁迫条件下起到了保护性物质和信号分子的作用,它的调节作用主要体现在植物的抗逆性上,因此,亚精胺与逆境胁迫之间的关系研究最为广泛[9,10]。由于它能与酶分子直接结合,使单位酶活力增加,过氧化氢含量下降,可以在一定程度上减轻植物受干旱胁迫的损害[11]。已有研究发现外源Spd起到缓解干旱胁迫对甜瓜幼苗伤害的作用,增强甜瓜幼苗对干旱逆境的适应性[12];魏晓凯等[6]研究发现喷施最佳浓度Spd能够有效提高烤烟幼苗的抗旱能力;胡胜男等[13]发现喷施Spd显著影响闽楠幼苗各项生理、光合指标,缓解了闽楠幼苗因干旱受到的损伤,并从光合角度筛选出适合闽楠幼苗的最佳Spd浓度。目前对于外源亚精胺与植物逆境胁迫虽有一定研究,但在南方红豆杉的领域研究还较为匮乏,且对南方红豆杉干旱胁迫后喷施最佳Spd浓度也未有研究。鉴于此,本试验通过设置不同干旱梯度胁迫以及不同浓度亚精胺复水处理,探究南方红豆杉叶片生理活性和功能物质对干旱胁迫的协同变化,比较不同浓度外源亚精胺复水处理对南方红豆杉生理特性的影响,以期为南方红豆杉的高产栽培和应用推广提供科学依据。
1 试验材料与方法
1.1 试验材料和试验地概况
2022年6-8月在河南省信阳市平桥区信阳农林学院林学院试验基地(114°06′E、31°12.5′N)进行南方红豆杉干旱胁迫试验。试验材料选用设施大棚中培养的3 a生南方红豆杉幼苗,植株生长一致、健康、无病虫害,平均树高0.7 m,胸径1.5 cm。棚内平均气温34~37℃,平均相对湿度60%~65%,利用一层遮阳网遮阴,通风良好,控制设施大棚内光照、温度和湿度,使所有植株均始终处于相同外界条件。
1.2 试验处理
于6月中旬选取48株3 a生幼苗期南方红豆杉,采用盆栽控水法(口径40 cm、高50 cm),进行干旱胁迫处理。调整土壤湿度一致后,根据正常生理需水量,设置3个处理:轻度胁迫T1处理(50 mL/d)、中度胁迫T2处理(33 mL/d)、重度胁迫T3处理(0 mL/d),设置正常浇水(100 mL/d)为空白对照(CK),单株处理,每处理12个重复,分别在处理的第0、7、14、21和28 d对南方红豆杉成熟叶片取样,液氮速冻后-80℃冰箱保存后留待备用。
亚精胺复水试验:干旱胁迫处理28 d后,因部分重度胁迫(T3组)可能导致植株死亡,且轻度胁迫(T1组)相比中度胁迫(T2组)各项指标影响较小,为使复水效果更为显著,对中度干旱胁迫的南方红豆杉进行复水试验。试验共分4个处理。N1处理:正常复水(100 mL/d)+喷施0.5 mmol/L外源亚精胺溶液;N2处理:正常复水(100 mL/d)+喷施1.0 mmol/L外源亚精胺溶液;N3处理:正常复水(100 mL/d)+喷施1.5 mmol/L外源亚精胺溶液;以正常复水不喷施外源亚精胺的中度干旱胁迫南方红豆杉为EGCK处理,单株处理,重复3次。喷施时靠近植株叶片的正反面,喷施量以叶片叶面水珠凝结滴落为标准,处理后的第3 d进行南方红豆杉成熟叶片的采集,使用液氮速冻后放-80℃冰箱进行保存,留待备用。
1.3 测定方法
本研究以乙醇浸出法、考马斯亮蓝染色法、氮蓝四唑光化还原法以及愈创木酚法分别测定了南方红豆杉叶片中的叶绿素总量、可溶性蛋白含量、SOD活性和过氧化物酶活性,具体操作参考刘家尧等[14]编著的《植物生理学实验教程》。
1.4 数据统计
通过Excel 2019进行标准曲线绘制与实验数据的统计,利用SPSS 24.0数据分析软件进行方差分析(ANOVA)以及Duncan多重比较分析,使用Origin 2021作图。
2 结果与分析
2.1 不同干旱胁迫对南方红豆杉叶绿素含量的影响
不同干旱处理下南方红豆杉叶片的叶绿素总含量如图1所示,在土壤持续干旱过程中,与CK相比,3个胁迫处理叶片的叶绿素总含量均呈增加趋势,整体上呈“升——降——升——降”的趋势,在第7 d和第21 d达到峰值,胁迫至第7 d,处理T3的叶片叶绿素总含量达到最高值,比CK叶片叶绿素总含量高126.1%,其次为T2处理,高出CK含量125.0%,最低为T1处理,高出CK叶绿素含量的77.2%;随着干旱时间的延长,在短暂的下降后,在第21 d叶绿素总量达到第2次峰值,此时处理T3的叶片叶绿素总含量最高,显著高于T2和T1处理(P<0.05),且比CK高出139.8 %,其次为T2和T1处理,分别比CK高52.3%、23.9%;胁迫至第28 d叶绿素含量均呈下降趋势,此时重度胁迫T3组显著低于CK组(P<0.05)。在叶绿素含量曲线波动中,在28 d之前,相比CK的叶绿素总含量总是T3>T2>T1,且短期干旱胁迫处理对南方红豆杉叶绿素含量呈正向促进作用。

图1 不同持续干旱胁迫对南方红豆杉生理活性的影响Fig.1 Effects of different persistent drought stresses on physiological activity of T.chinensis var.mairei
2.2 不同干旱胁迫对南方红豆杉丙二醛含量的影响
图1结果所示,随胁迫天数的增加3组叶片中丙二醛(MDA)含量均呈上升趋势,胁迫至21 d到达峰值,随后趋于平缓,且处理T1、T2南方红豆杉叶片中MDA含量上升趋势相比T3较为平缓,处理至21 d,T1、T2、T3组南方红豆杉叶片中MDA含量均达到最高含量水平,此时T3处理的MDA含量最高,且显著高于T2和T1处理(P<0.05),相比CK高出145.2%;其次为T2,比CK高出48.9%;而胁迫较轻的T1处理MDA含量最低,比CK高出39.1%。综合来看,3个不同胁迫程度的MDA含量均显著高于CK(P<0.05),且叶片MDA含量与胁迫梯度以及胁迫时间呈正相关。
2.3 不同干旱胁迫对南方红豆杉SOD活性的影响
南方红豆杉受到干旱胁迫后,其叶片中的保护酶活性均发生不同程度的变化。随着处理时间的增加,酶活性整体呈上升趋势,其中0~7 d各处理SOD活性变化较为平缓,胁迫至第14 d,此时3个处理的SOD活性显著增加且高于CK(P<0.05),此时T3处理的SOD达到最高活性水平,随后T2和T1分别在21 d和28 d达到峰值,且T3的峰值显著高于T2和T1处理(P<0.05),T1处理SOD活性上升较为缓慢。T3处理在胁迫14 d后,呈现下降趋势,胁迫至28 d,T3处理的SOD活性显著低于CK(P<0.05),此时T2处理最高,其次为T1。综合来看,T3处理的SOD活性上升最快,且峰值来临时间最早,其次为T2以及T1处理,且峰值过后叶片SOD活性持续高于对照组,说明T3受干旱胁迫的影响较大,且干旱胁迫对胁迫中前期的SOD活性呈正相关。
2.4 不同干旱胁迫对南方红豆杉可溶性蛋白的影响
由图1可知,南方红豆杉受到干旱胁迫后,其叶片中的可溶性蛋白含量均发生不同程度的变化。可溶性蛋白峰值水平显著高于同时期CK(P<0.05),总体呈现先升后降的波动趋势,其中0~7 d各处理可溶性蛋白含量变化较为平缓,胁迫至第14 d,此时3个处理的可溶性蛋白含量显著增加且高于CK(P<0.05),T3处理可溶性蛋白含量达到最高水平;随后T2和T1可溶性蛋白的含量分别在21 d和28 d达到最高水平,且T3的峰值含量显著高于T2和T1处理(P<0.05),T3处理在胁迫14d达到最高水平后,呈现下降趋势,胁迫至28 d,T3处理的可溶性蛋白含量显著低于CK处理(P<0.05),T2在胁迫21 d达到最高水平后,呈现下降趋势,T1在0~21 d期间可溶性蛋白变化较为平缓,胁迫至28 d可溶性蛋白含量到达最高水平。综合来看,短期干旱可以使南方红豆杉叶片的可溶性蛋白含量上升,与胁迫处理程度呈正向促进作用,并且与叶片SOD活性呈协同变化趋势。
2.5 外源亚精胺复水对南方红豆杉生理活性的影响
由图1可知,重度胁迫T3处理在第28 d时,叶绿素总含量、SOD活性和可溶性蛋白含量均低于正常组CK,可能达到生理阈值,不能作为复水最佳选择。为使复水效果更为显著,对中度胁迫T2处理进行亚精胺复水,生理活性变化如图2所示。在干旱复水后,4个不同处理叶绿素总含量水平均不同程度地高于复水前,且N2处理的南方红豆杉叶片叶绿素总含量最高,为EGCK的140.5 %,与复水前以及EGCK呈显著差异(P<0.05),其中N3处理叶绿素总含量仅次于N2处理,且喷施亚精胺处理均高于EGCK处理。复水后4个处理的MDA含量均有不同程度下降,且组间差异显著(P<0.05),其中N2处理的南方红豆杉叶片MDA含量最低,相比EGCK降幅最大,为EGCK的31.6%,且N3处理MDA含量降幅仅次于N2处理。复水后3个亚精胺处理的SOD活性均有不同程度上升,且组间差异显著(P<0.05),其中N2处理的南方红豆杉叶片SOD活性最高,相比EGCK上升程度最大,为EGCK的23.4 %,且N3处理SOD活性升高程度仅次于N2处理。复水后4个处理的可溶性蛋白含量均有不同程度升高,且组间差异显著(P<0.05),其中N2处理的南方红豆杉叶片可溶性蛋白含量最高,相比EGCK上升程度最大,为EGCK的34.5 %,且N3处理可溶性蛋白含量升高程度仅次于N2处理。综合来看,喷施3种浓度的亚精胺相比单一复水处理均有较强的促进作用,其中N2处理组对南方红豆杉的生理活性影响最大,N3处理的复水促进作用仅次于N2处理,其次为N1处理。
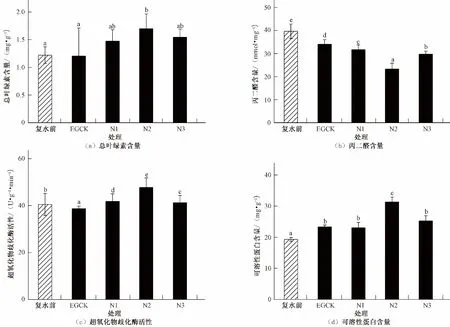
图2 不同浓度外源亚精胺复水对南方红豆杉生理活性的影响Fig.2 Effects of different concentrations of exogenous spermidine rehydration on physiological activity of T.chinensis var.mairei
3 讨论
干旱胁迫是植物在生长过程中遭受到主要的非生物胁迫,可导致植物生理代谢活动双向波动,且适度的干旱胁迫也可促进植物的生长量[15,16]。其中叶绿素在植物光能吸收和转化中起重要作用,当环境发生剧烈变化时,植物可以通过光合色素的动态变化来保证正常的光合作用,叶绿素总含量是直接反映其受逆境胁迫程度的重要指标之一[17]。本研究表明,相比对照组不同持续干旱胁迫均显著升高了叶绿素总含量(P<0.05),叶绿素含量首次出现高峰表明,南方红豆杉幼苗在短时间和低强度干旱胁迫下,会首先发生应激反应,提高叶片的叶绿素总含量,以适应干旱胁迫引起的植物代谢产物急剧下降,干旱胁迫下植物叶绿素总含量变化代表植物光合能力和对水分胁迫的敏感度[18]。胁迫至21 d,叶片中的叶绿素含量经历了短暂的下降,达到了第2个峰值,推测此时随着干旱时间持续增加,土壤干旱胁迫程度不断加深,南方红豆杉幼苗开始适应胁迫环境,从而产生了相应的生理反应,这与张玉玉等[19]研究结果相似。持续胁迫至后期,重度胁迫的叶绿素总含量骤然下降且低于对照,推测此时植物生理调节能力可能已达极限,造成不可逆失活,这与阮凌暄等[20]研究结果一致。本试验中适度干旱使叶绿素含量升高,叶绿素又是植物光合作用的重要因素,因此适度的干旱胁迫能够提高南方红豆杉的光合能力,增加植株的生物量[21]。
丙二醛是机体内脂质过氧化的最终产物之一,其含量高低可一定程度地反映植物受氧化伤害的程度[22,23]。本研究表明,3种不同持续干旱胁迫相比对照显著增加了丙二醛含量,且随胁迫梯度的增加,MDA含量的累积呈较高水平,其中持续重度胁迫处理的叶片MDA含量显著高于其他2组处理,说明持续重度胁迫对南方红豆杉的损伤最大,且随胁迫梯度的增加,MDA含量的累积呈较高水平。
植物对逆境胁迫的生理反应是以酶的催化活动实现的,SOD的活性水平已被证实与机体自身的抗性密切相关,是植物细胞内抵御活性氧(ROS)伤害的主要保护性酶类,对清除和阻止ROS的产生起到重要作用[24,25]。本研究结果显示,与CK相比3种不同持续干旱胁迫处理显著提高了SOD活性,且胁迫越严重越为显著。干旱胁迫越严重,酶活性峰值到来越早,持续胁迫至后期,重度胁迫条件下酶活性达到峰值过后呈下降趋势,推测在低强度和短时间干旱胁迫情况下,能诱导南方红豆杉叶片保护酶活性提高,并能清除部分活性氧,从而减轻干旱对植物机体的损害[26]。但持续的重度干旱胁迫打破了植物细胞自由基产生和清除的动态平衡,进而出现活性氧过量累积,加剧膜脂过氧化而造成整体膜的损伤,导致植物死亡[27]。由于在短期的干旱胁迫中,SOD活性水平与干旱胁迫呈正相关,因此适度干旱胁迫锻炼,也可以增加南方红豆杉的抗逆性,使植物在逆境环境中性状更为稳定。
可溶性蛋白的含量是植物组织里重要的渗透调节物质和营养物质,亦是植物抗旱能力的重要指标之一[28]。在干旱条件下,细胞会通过积累渗透调节物质来维持膨压,以保持植物正常生理活动,植物细胞缺水会促使植物主动累积可溶性蛋白等渗透物质,以降低细胞渗透势,增加细胞持水量[29]。本研究结果显示,南方红豆杉叶片可溶性蛋白含量总体呈先升后降的趋势,干旱胁迫越严重,可溶性蛋白含量峰值到来越早,此时可溶性蛋白含量越高。南方红豆杉为适应一定范围内的干旱胁迫,短时间内依靠合成更多的可溶性蛋白来调节自身的渗透势,这与徐澜等[30]研究结果一致。但植物依靠合成渗透调节物质蛋白质等来适应胁迫的能力是有限的,随着胁迫程度的增加,渗透调节系统逐渐紊乱,组织中的可溶性蛋白质含量也随之降低[31]。并且在干旱胁迫过程中,可溶性蛋白与SOD活性变化趋势呈现协同性,因此适度干旱锻炼,可溶性蛋白的含量对南方红豆杉抗旱性具有重要意义。
为探究南方红豆杉有效缓解干旱胁迫的方法,本研究对南方红豆杉持续干旱胁迫后进行亚精胺复水试验。结果显示,喷施外源亚精胺处理相比单一复水处理均有较强的促进作用,其显著提高了叶绿素含量、SOD活性以及可溶性蛋白,显著降低了MDA含量,有效提高了干旱胁迫下南方红豆杉的光合能力,加速了活性氧的清除,增加了南方红豆杉的渗透调节能力,缓解了因长期干旱胁迫而膜脂过氧化造成的损伤,这与孙天国等[12]研究结果一致。通过对比不同浓度的外源亚精胺复水,结果显示,1.0 mmol/L的外源亚精胺复水处理效果最为显著,相比0.5 mmol/L以及1.5 mmol/L,喷施1.0 mmol/L的外源亚精胺复水处理对南方红豆杉的干旱恢复作用最强。在实际生产中,对南方红豆杉干旱林区适当喷施1.0 mmol/L的外源亚精胺,也具有一定的缓解作用。本试验仅对幼苗期南方红豆杉叶片干旱胁迫以及外源亚精胺复水第3 d的生理活性变化进行了分析,只观测短期变化,对南方红豆杉生长发育的影响需长期观察继续研究。另外,外源亚精胺提高植物抗旱能力途径有很多种,如增强干旱信号的传导,提高南方红豆杉某些保护酶(主要是POD)的活性,直接参与氧自由基的清除,以及增强南方红豆杉的渗透调节能力等,这些途径之间的关联还尚未完全清楚,外源亚精胺对干旱胁迫更深层次的抗旱机理与作用途径还有待进一步研究。
4 结论
通过对幼苗期南方红豆杉持续干旱胁迫表明,前期的干旱处理对南方红豆杉起正向积极作用,干旱胁迫可显著提高南方红豆杉叶片的叶绿素总含量、MDA含量、SOD活性以及可溶性蛋白含量,并呈现不同的波动变化,特别是重度干旱胁迫下,其各项数值的变化幅度更大,且SOD活性、可溶性蛋白含量的峰值来临时间最早。其中对幼苗期南方红豆杉复水+1 mmol/L外源亚精胺的组合,能有效缓解干旱胁迫带来的损伤,并提高幼苗期南方红豆杉对干旱胁迫的恢复能力。由于适度的干旱胁迫导致生理代谢活动双向波动,从而促进植物的生长量,因此在干旱缺水地区,把干旱胁迫锻炼和喷施亚精胺结合,不仅可以降低植株耗水量、节约灌溉用水、维持作物生长和养分吸收,还可以提高叶片和植株水平的水分利用效率,增强植株的抗逆性。
