常绿阔叶林米槠叶片功能性状对不同海拔梯度的响应与适应
2023-01-31仲小茹潘昕昊叶雄英黄献峰孙荣喜
仲小茹,张 露,潘昕昊,叶雄英,黄献峰,柯 叮,孙荣喜*
(1.江西农业大学 林学院/江西省森林培育重点实验室,江西 南昌 330045;2.江西交通职业技术学院,江西 南昌 330032)
【研究意义】植物功能性状是指能够反映植物碳获取、水分传递、养分循环等重要生命活动的属性,直接或间接影响植物生长、存活和死亡,包括植物形态、生理等方面的特征[1]。叶片是树木地上部分物质交换最频繁的器官,也是植物光合作用的重要器官,对环境变化具有敏锐的捕捉力,在植物资源获取利用和竞争中起着关键作用[2]。叶片功能性状可以反映植物采取的生态策略。例如,叶经济型谱一端具有快速生长策略的物种表现为较高的光合速率、叶片氮磷含量、比叶面积和叶片导水率,另一端慢速防御策略的物种则具有较低的光合速率、叶片氮磷含量、比叶面积和叶片导水率[3]。目前,国内外学者有关叶功能性状的种内变异研究非常有限,大量研究关注种间变异。例如,叶片功能性状的耦合和权衡关系主要是基于物种间的关系而推断的,主要前提假设是种间变异大于种内变异[4]。然而,许多优势树种分布范围广,可以分布在不同的温度、降水和海拔梯度上,适应环境变化采取的生态策略较为复杂,在不同的环境中协调各功能性状而采取的措施有所不同,这也意味着该物种功能性状的变异也是巨大的。因此,了解植物种内功能性状的差异对于理解该物种的分布特征以及如何适应和响应环境有重要意义。而选择单一物种来研究环境梯度下叶片性状的变化,可以弱化系统发育的影响,凸显环境对功能性状变化的影响[5]。植物叶片性状是植物与环境长期互作的结果,海拔所带来的生境差异会对植物生长和分布产生很大影响。首先,温度是最重要的影响因素,一般海拔每减少100 m,温度增加0.6 ℃,这直接限制了土壤湿度和温度耐受性;其次,土壤养分可能存在差异,但不起决定性作用。虽然理论上在高海拔山地,土壤淋溶较多,土壤全氮随海拔增加而增加,土壤全磷随海拔的增加则先减少后增加,然而在热带亚热带森林土壤养分的异质性是巨大的,养分主要受到林分类型、坡度和坡位的影响。温度-植物生理假说(temperature-plant physiological hypothesis)认为:植物体内不同生理代谢速率受到温度调控,在低温条件下,与氮磷相关的光合酶活性和RNA 活性会降低,造成叶绿素含量降低。同时,由于土壤氮含量随海拔的升高而上升,因此,叶片中更多的氮分配到结构物质(细胞壁)中,导致比叶重增大[6-7]。还有研究显示,叶片功能性状在海拔梯度上的分布不尽相同,并没有随海拔的变化而变化。因此,研究海拔梯度下植物叶片功能性状的变化,有助于了解植物对环境变化响应的生理生态机制,同时也可以作为全球气候变化的指南针[8-9]。【前人研究进展】米槠(Castanopsis carlesii)作为我国亚热带地区常绿阔叶林的典型植被类型之一,主要分布在海拔1 300 m以下的山地或丘陵地带的常绿或落叶阔叶混交林中[10],对环境的变化具有较强的适应性。国内外学者有关叶功能性状的研究大量集中在种间变异,种内变异研究非常少。叶片性状对环境梯度的响应研究主要集中在群落水平上,但由于植物适应环境变化采取的生态策略较为复杂,在不同的环境中协调各功能性状而采取的措施有所不同,以植物群落为整体来研究会忽视不同物种之间的差异性。【本研究切入点】目前国内有关米槠研究主要集中在繁殖技术[11-12]、群落分布格局[13-14]、森林凋落物[15-16]、根系研究[17-18]和氮磷响应[19]等方面,但对其叶性状沿海拔梯度的响应机制鲜有研究。【拟解决的关键问题】本研究以九连山自然保护区常绿阔叶林不同海拔群体的米槠为对象,分析米槠叶片形态性状和生理性状随海拔梯度的变化规律,探讨叶片功能性状对环境变化的响应,为预测气候变化下植物物种分布、群落动态和森林生态系统结构功能的变化提供依据。
1 材料与方法
1.1 试验地概况
九连山自然保护区(24°29′~24°38′N,114°22′~114°31′E)位于江西省的南部,为南岭东部地区的核心部位,保存有较大面积的原生性亚热带常绿阔叶林,植被类型多样,被誉为“生物基因库”。境内海拔自300 m至主峰黄牛石1 430 m,地势南高北低,为中亚热带与南亚热带过渡地带。九连山气候具有大陆性、海洋性,受海拔高低以及地形变化的影响,形成复杂多样的小气候。年平均气温16.7 ℃,年均降水量2 070.4 mm,年均相对湿度85%;保护区内地质构造错综复杂,成土母质多样,按海拔高度自下而上依次分布有山地红壤、山地黄红壤、山地黄壤和山地草甸土,pH值4.5~6.5。
1.2 样品采集
根据米槠在九连山自然保护区的主要分布范围,从其分布下限开始采样,分为3个海拔梯度,按海拔由低到高分别记为:低(389~512 m)、中(514~626 m)、高(638~794 m)。每个海拔梯度内选取20 株左右米槠,株间距离要大于50 m。每株米槠在向阳面选取成熟且无病虫害叶片30 片,叶片摘取后用密封袋装好并放入便携式小冰箱内,带回实验室进行各种指标的测定。
1.3 米槠叶片性状的测定
1.3.1 叶片形态 采用Epson4990 型平面扫描仪对米槠叶片进行扫描,结合加拿大Regent Instrument 公司的WinSeedle 软件分析叶长、叶宽、叶长宽比和叶面积等指标。采用1/10 000 电子天平测量叶片鲜重,随后将叶片置于烘箱内(80 ℃)烘至恒重,测量叶片干重。计算比叶重(leaf mass ratio area,LMA,g/cm2)及叶干物质含量(leaf dry matter content,LDMC,mg/g)。计算公式:比叶重=叶片干量/叶面积;叶干物质含量=叶片干重/叶片鲜重×100%。
1.3.2 叶片生理 采用蒽酮比色法测定可溶性糖(mg/g)和淀粉含量(mg/g);参照Lichtenthaler 等[20]的方法测定叶绿素含量;非结构性碳水化合物(NSC)=可溶性糖+淀粉。
1.4 数据分析与处理
采用SPSS 21.0 对不同海拔梯度米槠群体的叶性状进行单因素方差分析,同时为了研究米槠叶片性状之间的相关性,对叶片性状数据进行Pearson 相关性分析。变异系数主要是用于说明不同海拔梯度内叶片性状的变异程度,其计算公式为:变异系数=标准差/平均数。
2 结果与分析
2.1 米槠叶性状对海拔的响应特征
2.1.1 海拔高度对叶片形态指标的影响 米槠叶片形态指标随海拔梯度变化的分析表明(图1),在调查的海拔梯度范围内叶面积、叶长和叶宽随着海拔的递增均呈减少的趋势,叶片长宽比和比叶重随着海拔的递增呈增加的趋势,叶片干物质含量总体呈现相对稳定的状态;叶面积、长宽比和比叶重在低海拔与高海拔之间存在显著差异(P<0.05),叶长和叶片干物质含量随海拔梯度变化差异不显著(P>0.05),叶宽在高海拔的含量显著低于其他海拔梯度(P<0.05),但在低、中海拔梯度间差异不显著(P>0.05)。
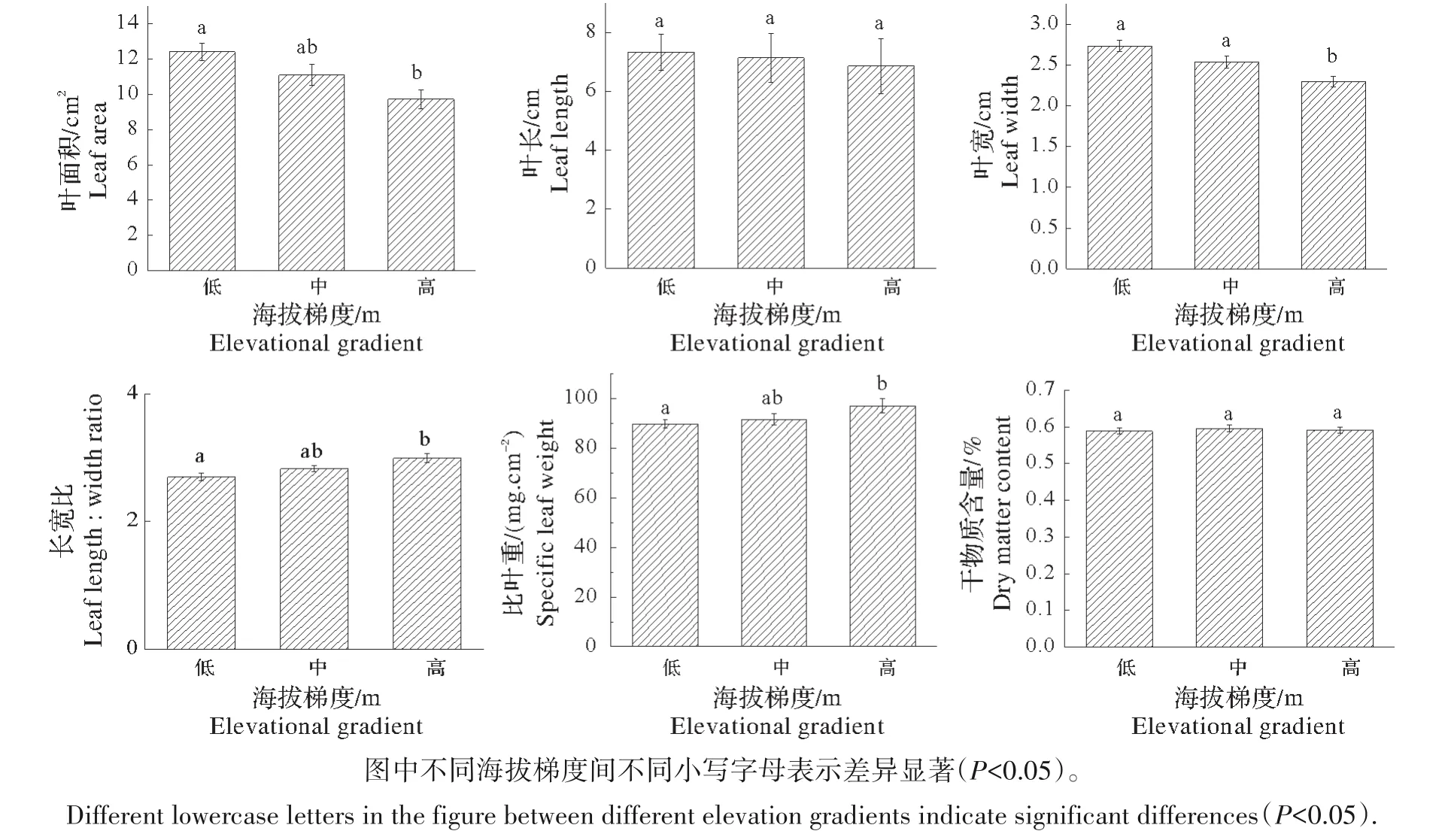
图1 米槠叶片形态指标随海拔梯度的变化Fig.1 Variations of leaf morphological indices with altitude gradient for Castanopsis carlesii
2.1.2 海拔对叶绿素a、叶绿素b和总叶绿素含量的影响 由图2可知,叶绿素a、叶绿素b和总叶绿素含量均随海拔升高而降低,叶绿素a、总叶绿素含量在低海拔与高海拔之间存在显著差异(P<0.05),但叶绿素b含量在各海拔梯度间差异不显著(P>0.05)。
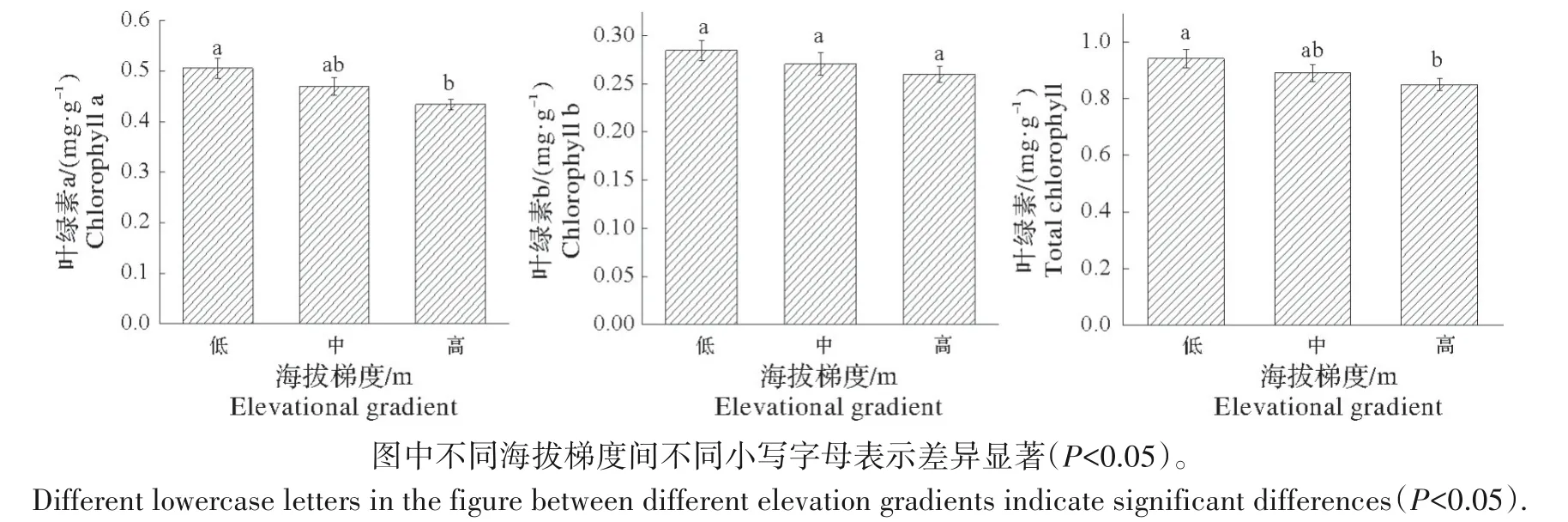
图2 米槠叶片叶绿素含量随海拔梯度的变化Fig.2 Changes of chlorophyll content in leaves of Castanopsis eyrei with elevation gradient
2.1.3 海拔对淀粉、可溶性糖和NSC 含量的影响 由图3 可知,淀粉、可溶性糖和NSC 含量均随海拔升高而增加,可溶性糖含量在低海拔与高海拔间存在显著差异(P<0.05);淀粉、NSC 含量在低海拔的含量显著低于其他海拔梯度(P<0.05),但在中海拔和高海拔间含量变化不显著(P>0.05)。
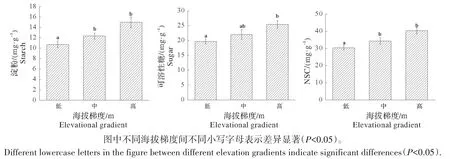
图3 米槠叶片淀粉、可溶性糖和NSC含量随海拔梯度的变化Fig.3 Changes of starch,soluble sugar and NSC contents in leaves of Castanopsis carlesii with altitude gradient
2.2 米槠叶片性状沿海拔梯度的变异规律
本研究比较了12 个叶片功能性状的变异系数,其总体的变异顺序由大到小依次为淀粉、可溶性糖、NSC、叶面积、叶绿素b、叶绿素a、总叶绿素、叶宽、叶长、比叶重、叶长宽比和干物质含量,其中淀粉、可溶性糖的变异系数最大,分别为30.16%和27.56%,干物质含量的变异系数最小,为6.73%(表1)。
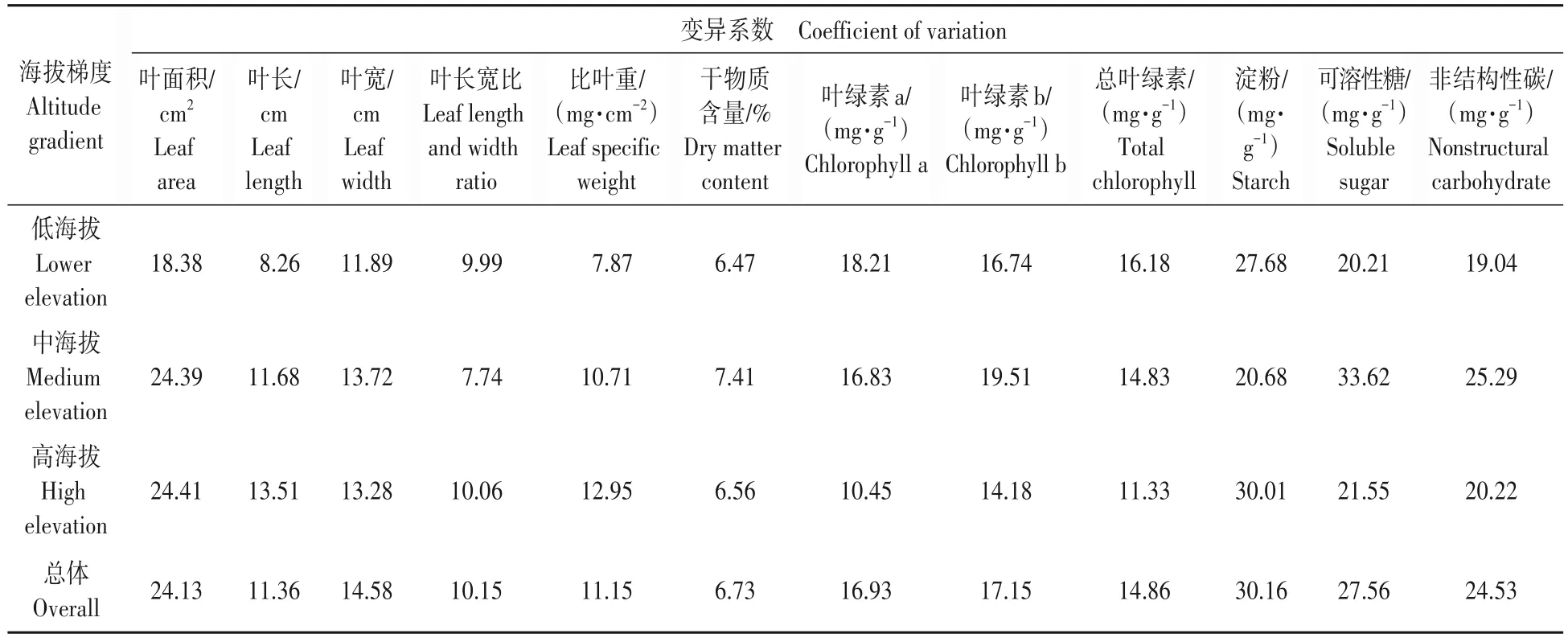
表1 不同海拔梯度米槠叶片性状数据统计Tab.1 Statistical analysis of leaf traits of Castanopsis carlesii at different elevations
不同海拔梯度内叶面积的变异范围为18.38%~24.41%,叶干物质含量的变异范围为6.47%~7.41%,叶长、叶宽、叶长宽比和比叶重的变异范围分别为8.26%~13.51%、11.89%~13.72%、7.74%~10.06%和7.86%~12.95%;与干物质含量、叶长、叶宽、叶长宽比、比叶重相比,叶片形态指标中叶面积变异系数相对较大,且在中高海拔的变异系数与总体的变异系数趋于一致,大于其在低海拔的变异系数。
本研究表明,叶绿素a 与总叶绿素含量在低海拔的变异系数最大,分别为18.21%和16.18%,在高海拔处的变异系数最小。叶绿素b 在中海拔的变异系数最大为19.51%,大于其在低海拔与高海拔的变异系数;淀粉含量在低海拔与高海拔处的变异系数大于在中海拔的变异系数,但可溶性糖在低海拔与高海拔处的变异系数小于在中海拔的变异系数,说明米槠各个叶性状在不同的海拔梯度范围存在不同程度变异。
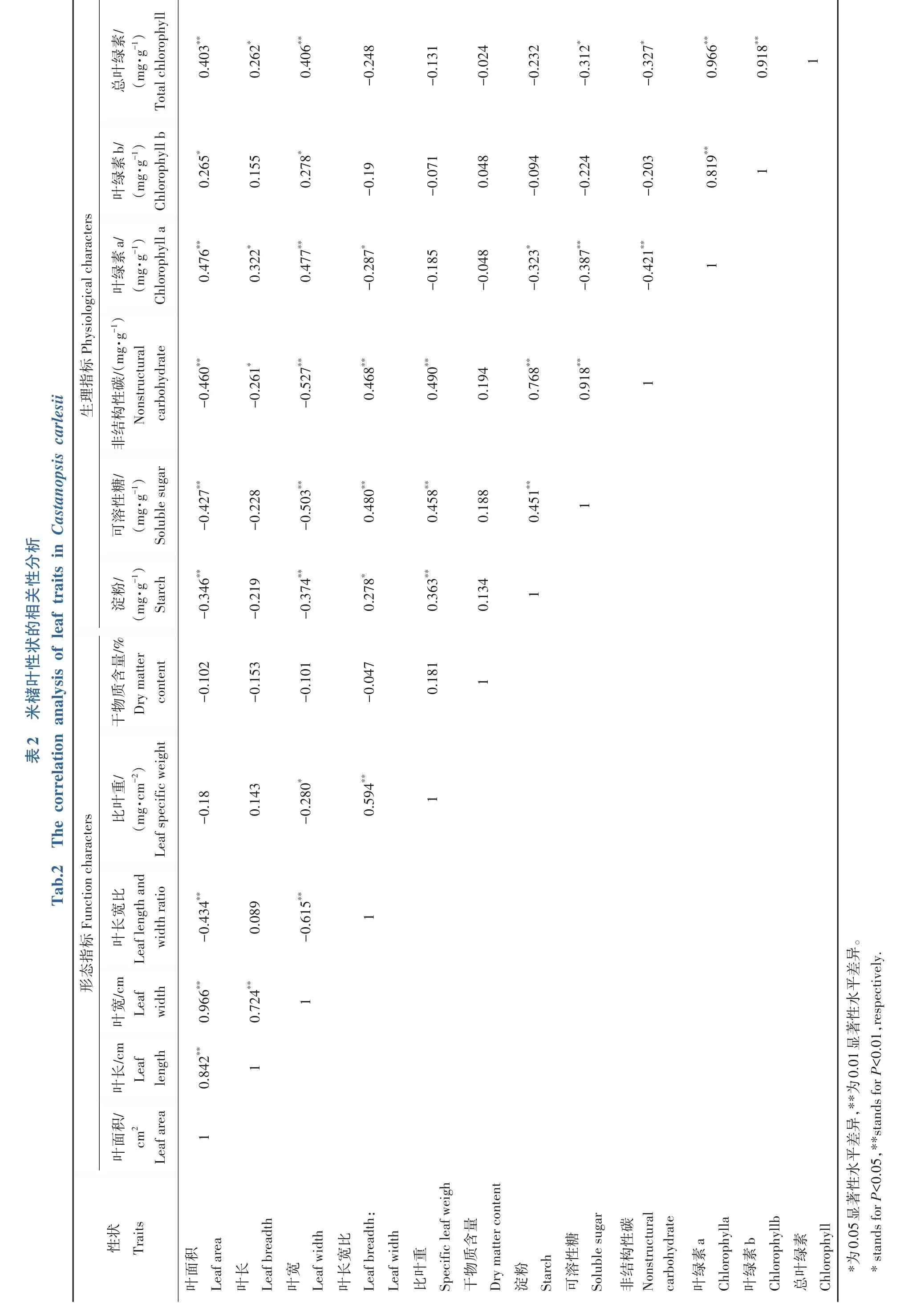
2.3 米槠叶片性状相关性分析
由各性状指数间的Pearson 相关分析结果(表2)可知,米槠叶片的各项指标之间存在着相关关系。叶片形态指标的相关分析表明:叶面积与叶长、叶宽呈极显著正相关(P<0.01),与叶长宽比呈极显著负相关关系(P<0.01),叶长与叶宽存在极显著正相关关系(P<0.01),叶长宽比与比叶重呈极显著正相关(P<0.01)。米槠叶片生理指标的相关分析表明:淀粉与可溶性糖、NSC 呈极显著正相关(P<0.01),叶绿素a 与可溶性糖以及NSC 呈极显著负相关(P<0.01)。除此之外,叶面积与淀粉、可溶性糖和NSC 呈极显著负相关(P<0.01),与叶绿素a、叶绿素b 和总叶绿素呈极显著正相关(P<0.01),比叶重与淀粉、可溶性糖、NSC 呈极显著正相关,叶宽与6 个生理指标均存在显著相关关系(P<0.01)。
3 结论与讨论
3.1 米槠叶性状对海拔的响应特征
3.1.1 叶面积、比叶重及叶干物质含量 叶面积和比叶重作为植物碳收获策略的重要叶特征之一[21],其大小直接与植物光合反应及环境适应相关。随着海拔升高,气温急剧下降,是影响植物生长繁殖及其分布的主要环境因子[22]。研究区域内随着海拔的升高,米槠叶面积逐渐变小,比叶重增加,并且在高低海拔梯度间存在显著差异,出现这种现象的原因是在低海拔地区种间竞争较为激烈,为满足生长需求,米槠通过增加叶面积捕捉更多光能。但在高海拔地区受低温、强紫外线等不利条件的影响,可供给植物利用的资源相对较少,植株通过减少叶面积、增加叶厚来降低叶片呼吸和蒸腾速率,以减少自身能量损耗并降低强光辐射对叶片的损害,因此较高的比叶重有助于植物对高海拔生境的适应[23]。在一定程度上,叶片干物质含量主要反映的是植物对养分元素的保有能力,易受水分的制约但不易受到其他叶片性状的影响[24],与环境的变化联系紧密。相关学者[25]研究表明高海拔处的植物更易遭受光温水分的胁迫,植物更倾向于保有自身养分来抵抗外界压力,高干物质含量的叶片相对坚韧,更有利于抵抗外界压力。但在本研究中,叶干物质含量总体呈现相对稳定的趋势,在不同海拔梯度并没有表现出显著的海拔差异,这可能与不同树种的生理结构不同及其所处环境存在差异有关。先前有学者在分析九连山米槠的群落特征时发现群落内米槠大乔木具有板状根系,表明该群落各海拔梯度内降雨量充沛,水分不是植物生长的主要限制因素,与同为壳斗科植物的蒙古栎[26]研究结果相一致。
3.1.2 叶绿素a、叶绿素b 和总叶绿素 叶绿素与植物的光合作用密切相关,直接参与光能的吸收、传递和转化,其含量大小影响植物的光合能力和光合器官的保护[23]。在本研究中,随着海拔的升高,叶绿素a、叶绿素b、总叶绿素含量均呈减少的趋势,这与以往研究[27]认为高山植物叶片具有相对低的叶绿素含量及对光的强反射性的结论相同。叶绿素的合成是一系列的酶促反应,受温度影响很大,因此高海拔的低温胁迫会降低酶活性进而抑制了叶绿素的合成,促进了叶绿素的降解。另一方面伴随着海拔的升高光照增强通过减少叶绿素的含量可减少因捕捉过度的光能而对叶面造成的伤害。
3.1.3 淀粉、可溶性糖及NSC 非结构性碳水化合物是植物新陈代谢过程中能量的主要储存形式,主要由可溶性糖和淀粉组成[28]。NSC 含量的变化能较好地反映出植物体碳收支情况以及对外界环境变化做出的响应与适应策略,可有效调节植物抵抗外界环境的胁迫[29]。在本研究中,米槠叶片淀粉、可溶性糖、NSC含量随海拔的反应趋势基本一致,均随海拔升高而增大。相关研究[30-31]认为可溶性糖作为植物体内活跃的碳储存形式,在植物抗低温、渗透交界、增强抗病性等方面发挥重要作用,低温和干旱环境可促进其积累。因此有可能是高海拔地区的低温胁迫促进了NSC 含量的积累来降低自身的渗透势和冰点,以此来提高米槠的抗寒适应性,起到冰冻保护剂的作用。植物组织中NSC 含量的高低被用来检验生长在高海拔的林木是否碳供应不足或生长受限制,在本研究中高海拔的NSC含量高于低海拔的含量,不支持关于林线形成的“碳受限假说”[32],并由此可以推测,随全球气候变暖米槠林线有可能向更高海拔移动。
3.2 不同海拔梯度下米槠叶性状的变化差异
变异系数体现的是叶片对环境因子的适应潜力,植物性状的变异系数高说明植物对环境有更高的适应性[33]。一般情况下,变异系数>50%为强变异,中等变异表现为20%<变异系数<50%,而变异系数<20%属于弱变异[34]。米槠作为样地的优势树种,对环境的变化具有敏锐的捕捉力,各叶片性状表现出较大的可塑性。其中淀粉、可溶性糖、NSC、叶面积具有较高的变异系数,变异系数范围在20%~50%,属于中等变异,这是由于叶片作为与环境接触面积最大的器官对环境变化的响应较为敏感,表现出较大的表型可塑性来适应不同海拔梯度的光照环境[35],以此来维持植株的水分平衡;另一方面,可溶性糖与淀粉含量是一个持续的动态转化过程,较高水平的可溶性糖含量有助于植株抵御低温环境的胁迫,因此在存在温度差异的不同海拔梯度间表现出较大的可塑性。与其他性状相比,干物质含量变异系数最小,表现出较强的内稳性,与多数研究结果一致[36]。
3.3 米槠叶性状间的相关关系
植物叶片的功能性状间联系紧密,在应对环境变化时并非调节某个单一性状而是通过各性状间的协同与权衡来适应环境变化[37]。研究植物性状间的耦合关系,可更好地理解植物在不同环境的生存策略。在本研究中,部分叶功能性状之间沿海拔梯度表现出显著的协同或权衡变化趋势。其中,叶面积与叶长、叶宽、叶绿素含量存在极显著的正相关关系,但与淀粉、可溶性糖、NSC 含量存在极显著的负相关关系,这是因为在低海拔地区植株更倾向于自身生长而非防御,较大的叶面积可促进植物的光合作用,同时植物的呼吸消耗大量能量。在高海拔地区,米槠受低温胁迫影响,更倾向于资源的保护而非快速生长,通过提高可溶性糖含量提高抗性并减少叶面积降低蒸腾。在植物处于胁迫状态时,叶绿素水解酶活性升高,叶绿素合成过程减弱,导致了叶绿素含量的降低[38]。
综上所述,本研究以不同海拔梯度的米槠为对象分析了叶片形态性状和生理性状随海拔梯度的变化规律。研究发现米槠叶片性状呈现出不同程度的变异,表明该物种在异质生境中具有较强的可塑性。在不同海拔梯度上米槠的生理生态适应策略也存在显著差异,在低海拔主要通过增大叶面积、光合速率、降低抗旱性来快速获取资源,但在高海拔为抵御低温胁迫,米槠产生一系列与防御相关的生态适应机制来适应环境,说明温暖湿润的环境更能促进米槠的生长,在全球未来气候变化下,该物种具有一定的适应能力,可能往更高海拔分布。本文为米槠的保护利用及预测未来气候变化下植物的空间分布提供了科学依据,但本研究仅从叶片的形态、光合、生理相关的指标来评价低温对米槠生长的影响,海拔梯度上众多生态因子发生变化,可能存在更复杂的适应机制,还需更深入的研究。
致谢:江西省科技厅项目(20202BABL215018)和江西省林业局林业科技创新专项(201801)同时对本研究给予了资助,谨致谢意!
