龙葵-镉抗性菌株修复镉污染土壤*
2023-01-30张文兴岳晓岚陈文祥张周位黄苑龄张安丰
张文兴 邓 强# 岳晓岚 陈文祥 张周位 黄苑龄 张安丰 何 海
(1.贵州省地质矿产中心实验室,贵州 贵阳 550018;2.贵州省贵金属矿产资源综合利用工程技术研究中心,贵州 贵阳 550018)
随着农药化肥的使用、工业废气废水的排放和金属矿石的冶炼等,重金属污染土壤问题日趋严重[1]。有研究表明,我国耕地土壤污染点位超标率为19.4%,其中无机污染物镉的污染点位超标率最大,达到7.0%,且镉依然是影响农用地土壤环境质量的首要污染物[2-3]。因此,研究适合镉污染土壤的修复技术尤为重要。
植物-微生物联合修复技术因原位修复、成本低、环境友好、修复效率高等优点,已成为目前治理重金属污染土壤主要的研究方向[4-5],[6]6,[7]。抗性微生物提高植物修复效率的机理主要包括两个方面[8-9]:一是抗性微生物通过合成一些金属载体(如铁载体)、固氮作用、溶磷作用、产吲哚乙酸等机制改善植物营养,以促进植物生长和发育,提高植物地上部分的生物量和重金属积累量;二是抗性微生物通过酸化土壤环境(产有机酸),提高土壤重金属的生物有效性,以促进植物对重金属的吸收和积累。例如,赵树民[10]的研究表明,向黑麦草根际土壤接种巨大芽孢杆菌(Bacillusmegaterium),可增加土壤有效磷、有效铁和有效态镉含量,同时提高了黑麦草的生物量和地上部分镉吸收量。张璐[11]77的研究发现,向甜高粱根系接种产铁载体细菌,可增加甜高粱地上部分和根部的铁和磷含量,同时提高其地上部分和根部生物量,且土壤中可溶出的镉浓度较高,使其地上部分镉浓度提高。江春玉等[12]的研究也表明,向印度芥菜和油菜接种伯克霍尔德氏菌(Burkholderiasp.)WS34(具有较高的脱氨酶活性和产铁载体能力,同时代谢分泌少量的生长素吲哚乙酸),可提高其生物量和镉积累量。由此可见,镉抗性微生物在促进植物生长、强化植物提取镉方面具有较大的潜力。
龙葵是一年生茄科草本植物,具有抗逆能力强、分布范围广的特点,已被证实对镉具有超富集作用,应用于镉污染土壤的修复研究中。例如,魏树和等[13]的研究发现,龙葵是一种镉超积累植物,在镉投加质量浓度为25 mg/kg时,龙葵茎和叶中镉分别为103.8、124.6 mg/kg,其地上部分镉富集系数为2.68。刘莉华[6]48-49向龙葵接种芽孢杆菌属(Bacillus)细菌,可使龙葵根干质量比对照高47.83%,地上部分镉达到125.21 mg/kg。可见,将龙葵应用于镉污染土壤的修复治理,有望通过提高龙葵根系发育并提高其生物量来增大龙葵对镉的吸收富集,从而提高土壤镉修复效率。
植物-微生物联合修复研究大多都是在未受重金属污染的土壤中添加有效态镉,利用温室盆栽进行试验,这与人类工业活动带来的土壤重金属镉污染源不符,与植物露天生长环境也存在差异,使得研究结果难以进行实际应用。此外,目前研究的镉抗性微生物主要从重金属污染区的土壤或植物里筛选,很少从未受重金属污染的宿主植物本身或其根系土壤筛选,缺少不同来源的镉抗性微生物对植物修复镉污染土壤的研究。本研究将从镉污染土壤、龙葵根际土壤和标准菌株筛选出的镉抗性菌株接种到龙葵根际土壤,通过原土室外露天盆栽试验,研究不同来源的镉抗性菌株对龙葵修复重金属镉污染土壤的强化作用,旨在为治理土壤中镉污染提供技术支持。
1 材料与方法
1.1 试验材料
镉污染土壤:在贵州省六盘水市某耕地(采样深度0~20 cm,该地区耕地受附近铅锌矿区人为活动影响),采用蛇形法、不锈钢小土铲取样10个点位,装入聚氯乙烯塑料袋中,共10袋,共计300 kg,带回室内风干、磨碎、混匀,过2 mm尼龙筛,制成综合试验样。土壤有机质为57.3 g/kg,全氮为2.84 g/kg,全磷为2.06 g/kg,全铁占10.72%(质量分数),pH为5.83,土壤含水率为44.24%,镉全量为13.87 mg/kg,有效态镉为3.27 mg/kg,土壤镉全量超过《土壤环境质量 农用地土壤污染污染风险管控标准(试行)》(GB 15618—2018)中风险管控值近7倍。
供试龙葵:种子采自贵阳市乌当区地矿新庄山体公园内,晒干,自封袋密封保存。
菌株分离样品:采自镉污染土壤综合样、龙葵根际土壤,新鲜土样装入牛皮纸袋于保温箱中放入冰袋保存,带回实验室于4 ℃冷藏;荧光假单胞菌(Pseudomonasaeruginosa)质控菌株[14]5246购自于广东某微生物科技有限公司。
产铁载体检测培养基按照文献[15]中CAS检测培养基进行配制。
1.2 主要试验试剂与仪器
牛肉膏蛋白胨培养基(营养琼脂);CdCl2·2.5H2O ,分析纯;GR60DA高压灭菌器;7000电感耦合等离子体发射光谱仪(ICP-OES)。
1.3 研究方法
1.3.1 镉抗性菌株的分离筛选与鉴定
土壤稀释液制备:分别称取10.0 g取自镉污染土壤和龙葵根际土壤的新鲜土样,置于装有90 mL无菌水且放有玻璃珠的250 mL锥形瓶中,150 r/min振荡培养10 min,静置30 min后用无菌吸管吸取1 mL上述土壤稀释液,置于9 mL的无菌水试管中,制成10-2的土壤稀释液,依次操作制备不同浓度梯度的土壤稀释液。
细菌纯化培养:用铂金丝接种环取1环制备好的土壤稀释液的上清液蛇形涂布于30 mL的牛肉膏蛋白胨培养基中,28 ℃培养72 h,挑选生长丰满、不同形态的单菌落进一步蛇形划线纯化培养得到单菌落。
镉抗性菌株筛选:在含镉(以CdCl2·2.5H2O配置)50、100 mg/L的牛肉膏蛋白胨培养基中(根据文献[15]的研究结果选择镉浓度),挑取上述经纯化培养的单菌落和荧光假单胞菌进一步蛇形划线培养,每种菌株每个镉浓度设置3个重复,28 ℃恒温培养48 h,挑选能够长出菌落的细菌即为镉抗性菌株。
产铁载体菌株筛选:将上述镉抗性菌株单菌落用铂金丝蛇形划线接种于CAS检测培养基中,28 ℃恒温培养箱中培养72 h,挑选出有橙色晕圈的菌株即为能分泌铁载体的菌株[16]。如此筛选出的抗镉且产铁载体细菌将作为强化龙葵修复镉污染土壤的特异微生物材料。
细菌鉴定:对菌株进行菌落和菌体形态观察、主要生理生化试验、16S rRNA基因序列同源性比对分析,以确定其分类地位。菌株形态及生理生化特性测定参照东秀珠等[17]的方法。抗镉产铁载体菌株制成冻干磁珠交由生工生物工程(上海)股份有限公司扩增、测序,将所得的扩增序列与GenBank数据库进行比对。与数据库中已有的16S rRNA基因序列进行同源性比较分析,可快速而准确地对细菌进行鉴定[18]。
镉抗性菌株分泌有机酸的测定参照《食品安全国家标准 食品中有机酸的测定》(GB 5009.157—2016)。
1.3.2 盆栽试验
龙葵种子首先在温室下育苗杯中萌芽育苗,待幼苗生长出7~8片叶时,挑选长势一致的幼苗移栽至装有1 000 g镉污染土壤的塑料盆(高11 cm,直径18 cm)中,每盆1株,置于楼顶(周围无其他污染源),露天盆栽,不施底肥,不加盖挡雨棚,根据盆中土壤缺水情况不定期浇水(去离子水),该试验地属于亚热带湿润温和型气候,年平均降水量1 449.7 mm。移栽两周,待植株生长良好后,在植物根部接入光密度(以600 nm波长处的光密度(OD600)计)为0.5左右的菌液10 mL,每周接种1次。设置以下7种处理:不接菌的空白对照(CK);接种镉污染土壤镉抗性菌(BV);接种龙葵根际土壤镉抗性菌(PH);接种荧光假单胞菌镉抗性菌(PA);接种镉污染土壤镉抗性菌和荧光假单胞菌镉抗性菌(BV+PA);接种龙葵根际土壤镉抗性菌和荧光假单胞菌镉抗性菌(PH+PA);接种镉污染土壤镉抗性菌、龙葵根际土壤镉抗性菌和荧光假单胞菌镉抗性菌(BV+PH+PA),每个处理设置3个平行样。
为防止重金属镉淋溶渗漏损失,在盆下放置塑料托盘并将渗漏液倒回盆中,植株生长120 d后收获,将植株轻轻拔起后,抖动根部,将龙葵根际土壤收集于布袋(或自封袋)中,带回实验室自然风干,手工磨碎筛分至小于2 mm混匀,分析样制成小于0.15 mm颗粒。将龙葵根部土壤清理干净,小心清洗根系,得到一株完整的植物,测量植物株高,编号拍照,将洗净的植株放入恒温干燥箱中105 ℃杀青5 min,70 ℃干燥48 h,称量植株地上部分和地下部分干质量,分别手工磨碎筛分至小于0.85 mm混匀,分析样制成小于0.43 mm颗粒。土壤样镉全量的测定参见《区域地球化学样品分析方法 第5部分:镉量测定 电感耦合等离子体质谱法》(DZ/T 0279.5—2016),有效态镉含量的测定采用二乙基三胺五乙酸浸提法;植物样镉全量的测定参见《食品安全国家标准 食品中镉的测定》(GB 5009.15—2014)。
1.4 数据处理与分析
计算富集系数,反映龙葵从土壤中吸收重金属的能力[19];计算转运系数,表征龙葵将重金属从地下部分向地上部分运输的能力[20];计算修复效率(镉去除率),表征土壤中重金属被活化、吸收、转化的能力[21-22]。
2 结果与讨论
2.1 镉抗性菌株筛选与鉴定
按照镉抗性菌株的分离筛选与鉴定方法,从镉污染土壤和龙葵根际土壤的系列稀释液中分别得到7、15株细菌。在50 mg/L镉污染的牛肉膏蛋白胨培养基中进行耐受性试验后,筛选出7株生长较好的菌株(镉污染土壤2株,龙葵根际土壤5株);在100 mg/L镉污染的牛肉膏蛋白胨培养基中筛选出5株生长较好的菌株(镉污染土壤2株,龙葵根际土壤3株)。在CAS检测培养基中进行细菌产铁载体检测,筛选出7株产铁载体菌株,分别是:100 mg/L 镉污染土壤1株(2020Z-4),50 mg/L镉污染龙葵根际土壤3株(2020Z-5、2020Z-7、2020Z-11),100 mg/L镉污染龙葵根际土壤3株(2020Z-6、2020Z-8、2020Z-12)。光密度在0.5左右的菌液对应的两株菌株分别是2020Z-4(OD600=0.443)和2020Z-8(OD600=0.449)。荧光假单胞菌经不同镉浓度耐受性试验、细菌产铁载体检测、菌悬液光密度测定,各筛选出1株抗镉产铁载体菌株,记为2020Z-PA1(耐受50 mg/L镉污染,OD600=0.510)、2020Z-PA2(耐受100 mg/L镉污染,OD600=0.619)。选择2020Z-4、2020Z-8和2020Z-PA1进行主要生理生化研究和细菌鉴定。
菌株主要生理生化特性见表1。对菌体总脱氧核糖核酸(DNA)进行提取,利用细菌16S rRNA基因通用引物(27F:AGAGTTTGATCMTGGCTCAG,1492R:GGTTACCTTGTTACGACTT)对供试菌株进行扩增、测序,结合生理生化特性,菌株鉴定结果见表2。

表1 镉抗性菌株的主要生理生化特性1)Table 1 Major physiological and biochemical properties of the cadmium-resistant strains

表2 镉抗性菌株鉴定结果Table 2 Identification results of cadmium-resistant strains
2.2 镉抗性菌株对龙葵修复镉污染土壤的效应
采用原土室外露天盆栽试验能更加真实地模拟龙葵野外生长状况,分别向龙葵根际土壤添加不同组合的镉抗性菌株,以研究镉抗性菌株对龙葵吸收镉的强化作用。
2.2.1 镉抗性菌株对龙葵生长的影响
从图1中可以看出,BV、PH、PH+PA和BV+PH+PA可促进龙葵地下部分干物质增加,说明接菌处理可促进龙葵根系的发育。其中,PH+PA的龙葵地上部分和地下部分干物质较CK均显著增加,分别提高了31.07%和75.59%,表明接菌处理可促进镉污染土壤中龙葵的生长。PA处理则阻碍龙葵生长。

注:同一指标不同字母表示处理间差异显著(P<0.05),图2至图8同。图1 不同镉抗性菌株接种下的龙葵地上部分和地下部分干质量Fig.1 Shoot and root dry weight of the Solanum nigrumwith different cadmium-resistant strains
张璐[11]76-77的研究认为,假单胞菌属(Pseudomonas)具有分泌铁载体的能力,可促进甜高粱对铁和磷等营养元素的吸收,从而促进甜高粱的生长。邓平香等[14]5252研究表明,荧光假单胞菌可分泌植物生长素吲哚乙酸和产铁载体,促进东南景天的生长;同时分泌大量有机酸,促进CdO溶解。PA+PH菌株的光密度均比BV大,推测在龙葵根际土壤中苍黄假棍状杆菌和荧光假单胞菌的丰度高于其他处理,其产铁载体的能力更强,从而促进龙葵生长。PA菌株为荧光假单胞菌,除了分泌铁载体能力强外,分泌有机酸的能力也强,从而溶解土壤中更多的重金属,导致龙葵吸收大量的重金属,阻碍自身生长。
2.2.2 镉抗性菌株分泌有机酸对土壤pH和有效态镉含量的影响
从图2中可以看出,BV菌株分泌乳酸较显著,PH菌株分泌苹果酸较显著,PA菌株分泌酒石酸和柠檬酸较显著;PA菌株分泌的有机酸总量高于BV和PH菌株,且显著高于BV菌株。与CK相比,所有接菌处理的土壤pH均显著降低,其中BV和PA土壤pH分别下降了0.14和0.13(见图3)。接菌处理后土壤有效态镉含量较CK有不同程度增加,其中PA土壤有效态镉含量显著增加,增加了26.65%(见图4)。菌株产酸能力的强弱可反映其活化重金属的能力。
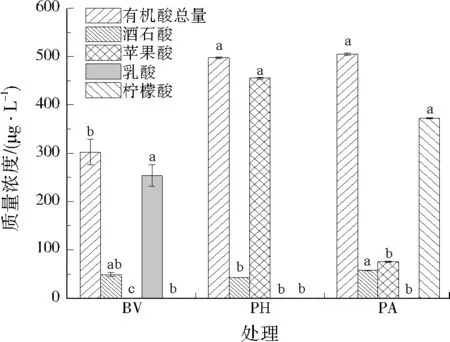
图2 3种单一菌株分泌的有机酸质量浓度Fig.2 The content of organic acids secreted by three single strains
邓月强等[23]研究认为,巨大芽孢杆菌在植物生长过程中代谢产生有机酸,对土壤镉有一定的活化作用,从而增加土壤有效态镉含量,该菌使伴矿景天对镉的积累量为95.40 μg/棵,土壤活化镉总量为186.21 μg/盆。江春玉[24]65研究表明,假单胞菌属使土壤中有效态镉含量较对照组提高16.63倍,土壤活化镉率为48.65%;这是由于菌株在代谢过程中产生了大量酸性物质,使原本处于沉淀态的重金属被活化成为可溶态的金属离子。本研究发现,PA菌株在培养液中分泌有机酸的总量高于BV和PH菌株,其土壤pH相比于CK显著降低,土壤有效态镉含量显著高于其他处理,且土壤活化镉总量为(1 480.00±400.37) μg/盆,高于文献[23]的研究,表明PA菌株在分泌有机酸和活化镉方面具有明显的优势。

图3 不同镉抗性菌株接种下的土壤pHFig.3 Soil pH under inoculation with different cadmium-resistant strains

图4 不同镉抗性菌株接种下的土壤有效态镉质量浓度Fig.4 Effective-cadmium content under inoculation with different cadmium-resistant strains
2.2.3 镉抗性菌株对龙葵镉积累量的影响
龙葵镉积累量为生物量与镉含量的乘积。与CK相比,PH、PH+PA和BV+PH+PA龙葵镉总积累量显著增加,分别提高了7.60%、30.28%和25.40%(见图5),其中菌株PH+PA龙葵的镉总积累量最高,达到184.11 μg/棵,低于文献[11]的研究结果(甜高粱的镉总积累为3.53 mg/棵)。究其原因,文献[11]是向洁净的土壤中添加有效态镉,且进行温室盆栽,不受淋溶的影响,植物对原有的有效态镉有更好的吸收能力;而本研究PH+PA的龙葵受到淋溶作用,使原本活化的镉部分随淋溶流失,龙葵吸收的有效态镉减少,导致最后龙葵镉总积累量较低。PA菌株虽然分泌有机酸能力强,但土壤有效态镉含量的增加阻碍了龙葵的生长,使龙葵生物量减小,因此龙葵镉总积累量最低。

图5 不同镉抗性菌株接种下的龙葵地上和地下部分镉积累量Fig.5 Shoot and root cadmium accumulation of the Solanum nigrum with different cadmium-resistant strains
PH+PA和BV+PH+PA均可以显著增加龙葵地上部分镉积累量,较CK分别增加了31.53%和25.22%;地下部分镉积累量有一定的增加,但相比CK无显著差异。可见,龙葵将吸收的镉大部分积累在地上部分,PH+PA对龙葵地上部分积累镉具有更明显的优势。
2.2.4 镉抗性菌株对龙葵镉含量的影响
与CK比较,PA龙葵地上部分和地下部分镉含量均显著增加,分别增加了129.29%和73.19%(见图6)。与CK比较,PA龙葵地上部分和地下部分富集系数增加显著,分别增加了128.40%和72.90%(见图7)。张璐[15]研究证实,假单胞菌属对土壤中镉具有活化作用,并能显著增加植物地上部分和根部的镉含量。江春玉[24]75-76的研究也认为,假单胞菌属对土壤中沉淀态镉具有明显的活化作用,可增加土壤中水溶态镉含量。本研究中,PA菌株为荧光假单胞菌,分泌有机酸总量大,对土壤镉具有较强的活化作用,并能增加龙葵对镉的吸收,从而提高龙葵镉含量和富集系数。
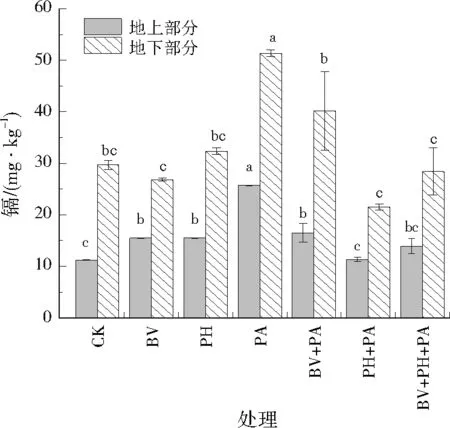
图6 不同镉抗性菌株接种下的龙葵地上部分和地下部分镉质量浓度Fig.6 Shoot and root cadmium concentration of the Solanum nigrum with different cadmium-resistant strains

图7 不同镉抗性菌株接种下的龙葵富集系数与转运系数Fig.7 Enrichment factor and transport coefficient of the Solanum nigrum with different cadmium-resistant strains
除BV+PA外,其他接菌处理的龙葵转运系数较CK均显著提高,BV、PH、PA、PH+PA和BV+PH+PA分别提高了52.63%、26.32%、31.58%、39.47%和31.58%,表明在镉污染土壤中接种镉抗性菌能提高龙葵地下部分镉向地上部分运输。究其原因,BV菌株是镉污染土壤土著菌;PA菌株为荧光假单胞菌,对土壤镉有较强的活化作用;BV+PA可将土壤镉大量活化,龙葵根系吸收大量的活化镉反作用于本身,阻碍龙葵生长,减少了龙葵地上部分对镉的吸收,导致转运系数不高。
2.2.5 镉抗性菌株对土壤镉含量和修复效率的影响
所有处理的土壤中镉均低于修复前(13.87 mg/kg),其中PH+PA土壤镉较CK下降了4.13%,修复效率较CK显著提高了67.83%(见图8)。可见,PH+PA对龙葵修复镉污染土壤具有强化作用,龙葵生物量和镉总积累量最大,对镉污染土壤的修复效率最好。这与MA等[25]的研究结论一致,即内源细菌能够帮助宿主植物提高其本身的重金属耐受性和生物量,以及提高植物对重金属的吸收,增加植物体内的重金属浓度来提高植物修复效率。PH菌株为龙葵根系土壤内源细菌,PA菌株虽然是外源细菌,但其产铁载体和有机酸能力强,使得PH+PA提高了龙葵对镉污染土壤的修复效率。

图8 不同镉抗性菌株接种下的土壤镉质量浓度和修复效率Fig.8 Soil cadmium content and remediation efficiency with different cadmium-resistant strains
淋溶镉量为修复前土壤镉总量减去修复后龙葵镉总积累量和修复后土壤镉总量所得的差值。试验中,尽管在盆下放置了塑料托盘,但遇到降雨量大的季节则会导致镉淋溶渗漏损失。结合SPSS统计分析得出,土壤修复效率和淋溶镉量的Pearson相关系数为0.98,表明土壤修复效率和淋溶镉量呈显著正相关。PH+PA土壤镉去除量的13.84%由龙葵吸收积累,而土壤镉去除量的86.16%随淋溶流失,反映出土壤修复受淋溶影响很大。如果在旱地或田间进行应用,由于镉淋溶量占比高,对深层土壤和地下水存在潜在风险;因此,后续应加强镉淋溶风险处置研究,例如,探索一种辅助材料,既能将土壤镉活化,又可稳定存在于土壤,同时还能被植物吸收。
2.2.6 土壤镉有效性与土壤修复效率的关系
土壤重金属有效性很大程度上决定了植物对重金属的吸收量,从而影响植物修复效率[26]。与CK相比,PA菌株分泌有机酸总量较大,处理后土壤pH降低程度较大,土壤有效态镉含量、龙葵镉含量和富集系数均较高,而土壤修复效率较低。究其原因,PA淋溶镉量与CK无明显差异,但有机酸的大量分泌促进土壤镉活化,土壤镉有效性增加到一定程度会阻碍龙葵的生长,龙葵生物量减小使得镉总积累量降低,导致土壤修复效率低于CK。
与CK相比,PH+PA土壤pH较低,土壤有效态镉含量和龙葵地上部分镉含量增加不明显,地下部分镉含量较低,但土壤修复效率显著增加。究其原因,一方面,PH菌株作为内源细菌能够提高宿主植物的重金属耐受性和生物量,PA菌株分泌有机酸能力强,可增加土壤镉有效性,PH+PA的龙葵生物量达到最大,镉总积累量最高,使得土壤镉含量降低;另一方面,淋溶镉量的增加也会使土壤镉含量降低,从而使土壤修复效率显著提高。
综合分析发现,土壤修复效率可由龙葵对镉的积累量和淋溶镉量共同决定。土壤镉有效性即为微生物分泌有机酸活化镉的能力,包含了龙葵镉积累量和淋溶镉量以及土壤中多余的活化镉量。因此,在尽量降低淋溶镉量的条件下,土壤镉有效性的增加和龙葵生物量的提高可增加龙葵镉积累量,以提高龙葵对镉污染土壤的修复效率。
3 结 语
从镉污染土壤中筛选出一株镉抗性微生物,鉴定为贝莱斯芽孢杆菌;从龙葵根际土壤中筛选出一株镉抗性微生物,鉴定为苍黄假棍状杆菌;荧光假单胞菌经筛选后得到具有同样的抗镉产铁载体功能的菌株。
原土室外盆栽龙葵试验表明,与CK相比,PH+PA可显著提高龙葵地上部分干质量、地下部分干质量、龙葵镉总积累量、地上部分镉积累量、转运系数和土壤修复效率,分别提高31.07%、75.59%、30.28%、31.53%、39.47%和67.83%。PH+PA对龙葵生长、富镉能力以及土壤修复效率具有一定的强化作用。PH+PA土壤镉去除量的13.84%由龙葵吸收积累,而86.16%随淋溶流失。
