长江干流轮虫群落结构的空间分布及其影响因素
2023-01-19钟可儿陈敏建田盼盼马吉顺周琼
钟可儿,陈敏建,田盼盼,马吉顺,周琼
1.华中农业大学水产学院/农业农村部淡水生物繁育重点实验室,武汉 430070;2.长江经济带大宗水生生物产业绿色发展教育部工程研究中心,武汉 430070
浮游动物在水生生态系统物质循环、能量流动等生态过程中扮演着重要角色,是天然水域食物链中关键环节之一[3]。轮虫是淡水浮游动物的重要组成部分,是食物网中维系生产者和次级消费者的关键组分,对环境变化的反应敏感,被作为水生态系统中的重要环境指示生物[4]。在不同水体中,环境因子(水温、溶氧、总氮、总磷等)对轮虫结构特征的影响程度亦有差别[5-6],轮虫的种类组成往往随水质的变化而发生改变,轮虫迁移能力较弱,对水质的响应比瞬时水质监测更为长效[7]。因此,轮虫的群落结构特征可以有效地反映水体健康状况[8]。轮虫的监测与研究已被广泛应用于河流、湖泊、水库等各种水生态环境的健康评价中[9-10]。
近年来,研究人员在长江流域各江段及其附属水体开展了较多基于浮游动物的水质评价及其环境驱动因素研究[11-13],但未见对整个长江干流水域的浮游动物群落进行系统性的调研与分析,亦无对长江干流轮虫空间分布的专门报道。2020年1月1日农业农村部在长江开始实施“十年禁渔”计划,本研究对长江干流(源头至下游)的轮虫群落结构特征与分布规律进行系统调查及分析,探讨影响长江干流水域轮虫群落结构的关键环境因素,旨在为长江水域生态保护提供理论依据,同时为长江“十年禁渔”提供饵料生物变化方面的基础资料。
1 材料与方法
1.1 采样区域概述
本研究的采样时间为2019年6-9月,将长江干流全流域自西向东划分为27个江段,各采样江段根据城市上、中、下游的地理形态特征及实际采样状况设置1~3个采样站点,各调查江段的分布情况如表1所示。

表1 长江干流各江段采样断面信息Table 1 Information of sampling sections in the mainstream of the Yangtze River
1.2 轮虫样品采集与鉴定及环境因子测定
轮虫样品与水质样品采集、水质物理因子的测定同步进行。
在每个采样点分别采集轮虫的定性与定量样品。轮虫定性样品使用25#浮游生物网在水体表层0.5 m处绕“∞”形反复缓慢拖拽后将样品收集至采样瓶内,加入4%福尔马林溶液进行固定。定量样品则在表层0.5 m处使用5 L采水器进行采集后使用25#浮游生物网过滤,将过滤后的轮虫转移至采样瓶内,加入鲁哥试液进行固定并贴好标签。将所有样品带回实验室静置24 h后将样品浓缩以待镜检,使用1 mL计数框在倒置显微镜下进行物种鉴定与全片计数,种类鉴定参考相关文献[14-15]。
使用便携式水质分析仪(YSI,美国)现场测定酸碱度(pH)、溶氧(DO)、盐度(Sal)、总溶解固体(TDS)、电导率(Cond)、水温(WT),使用塞氏透明度盘对水质透明度及水深进行测试。另采取1 L水样于4 ℃下避光保存,将其带回实验室后对总氮(TN)、总磷(TP)、硝态氮(NO3--N)、氨氮(NH4+-N)等指标进行测定,测定方法见参考文献[16]。
即便在最严酷的革命斗争中,列宁也严格遵守党内民主程序,利用党的政治纪律和政治规则妥善处理各项重大问题,积极维护团结稳定的政治大局,并未凭借自己的威望和地位以势压人。在病危时,列宁虽然指出斯大林性格粗暴,缺乏团结同志、总揽全局的政治素养,无法很好地胜任总书记一职,但是也仅仅“建议同志们想个办法”将他从总书记这个职位上调离,并诉诸党的最高权力机关按程序解决,从未越权、擅权、弄权。
1.3 数据处理与分析
① 采用优势度(Y)表示物种的优势程度,计算公式如下:
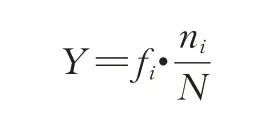
式中,fi表示第i物种的出现频率,ni表示第i物种的个体数,N表示同一样品中的轮虫总个体数。将Y≥0.02的物种视为优势种,使用自编程函数计算优势度[3],并划分污染指示类型[17-18]。
②采用Shannon-Wiener 多样性指数(H')、Pielou均匀度指数(J)对水质进行健康评价,各指数计算公式及评判标准如下。
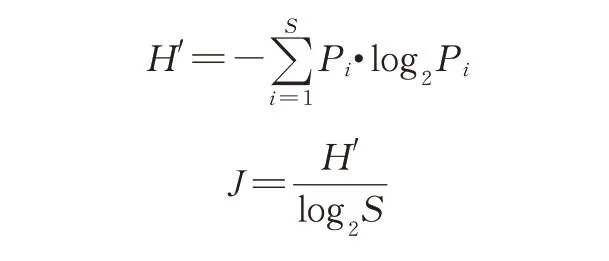
上述公式中,S为同一样品中轮虫的物种数之和,Pi表示第i物种的个体数(ni)与N(同一样品中的轮虫总个体数)的比值。
③使用 R软件4.2.1对各区域轮虫群落进行非度量多维尺度分析(NMDS)、聚类分析。
④使用R软件4.2.1 探究长江干流轮虫群落结构与环境因子的关系。物种的分布可能呈现单峰或者线性2种情况,利用除趋势对应分析(detrended corre‑spondence analysis, DCA)进行确定。典范对应分析(canonical correspondence analysis,CCA)适 用 于DCA排序轴最大梯度长度值大于4的情况,而冗余分析(redundancy analysis, RDA)适用于DCA排序轴最大梯度长度值不大于4的情况。pH以外的数据在统计分析前进行lg(x+1) 转换。
⑤使用数据分析软件SPSS 26.0进行不同江段间密度、生物量、多样性指数等的单因素方差分析,并进行Pearson相关性分析,以探究环境因子对长江干流轮虫群落结构与分布规律的影响。
2 结果与分析
2.1 长江干流各江段生境特征
根据27个采样江段的水体理化因子进行聚类分析,在聚类距离为2.5的水平上可将其划分为4个组群,其空间分布规律明显。第1个组群为高山区,包括自源头曲麻莱至丽江的6个江段(S1-S6),横跨青藏高原、横断山区,其生态环境恶劣,水温较低。第2个组群为峡谷区(S7-S17)主要为峡谷地带,其水流湍急、水量丰沛 ,建有大量水电站。第3个组群为平原区,包括中、下游的9个江段(S18-S26),平原区水面宽阔、水流缓慢。第4个组群为S27,启东江段位于河口,受咸水控制,与长江下游生境有较大差异。生境区域划分的聚类结果见图1。
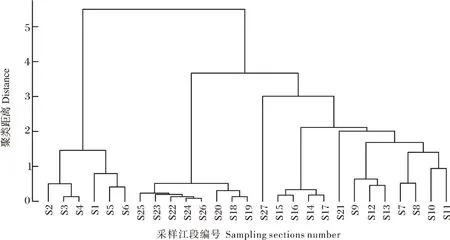
图1 长江干流27个采样江段的生境聚类Fig.1 Habitat clustering of the 27 sampling sections in the mainstream of the Yangtze River
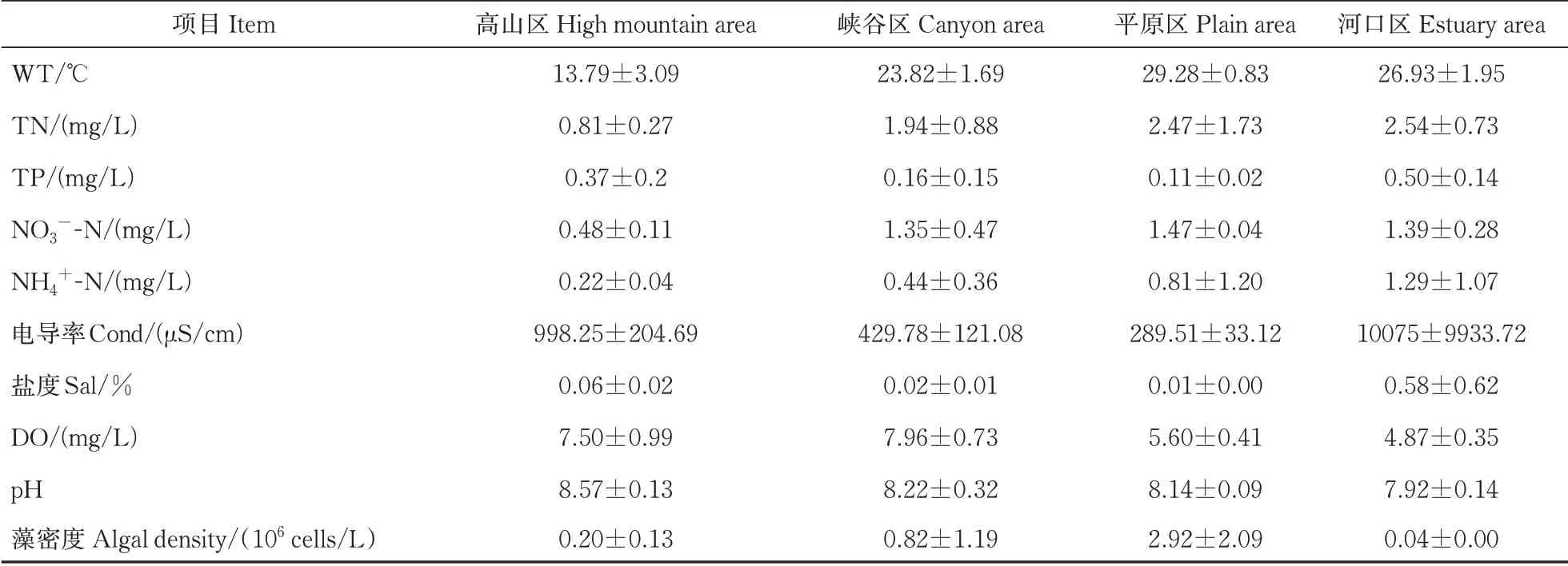
表3 长江干流各区域环境参数Table 3 Environmental parameters at each area in the mainstream of the Yangtze River
长江干流各采样江段TN质量浓度范围为0.67~7.08 mg/L,最高值出现在平原区的S21,属于劣Ⅴ类水体;最低值出现在高山区的S1。各采样江段NO3--N、NH4+-N质量浓度波动范围分别为0.32~1.96、0.15~3.98 mg/L,均值分别为1.20、0.54 mg/L。各采样江段DO的范围为4.87~9.24 mg/L,DO均值为6.96 mg/L。各采样江段水温变化范围为8.68~30.2 ℃,各区域中,高山区平均水温为13.79 ℃,峡谷区平均水温为23.82 ℃,平原区、河口区平均水温为29.28 ℃、26.93 ℃,整体上呈现由源头向下游递增的趋势。各江段电导率在219.5~10 075 μS/cm波 动,TDS在0.37~6 469.67波动,河口区电导率、盐度明显大于其他区域。
2.2 轮虫群落结构特征
1) 种类组成。研究期间长江干流各江段共鉴定出轮虫11科37属85种,主要为臂尾轮科(11属31种)、腔轮科(3属13种)、镜轮科(6属9种)、鼠轮科(1属7种)、腹尾轮科(3属6种)、疣毛轮科(3属6种)的种类。高山区、峡谷区、平原区、河口区分别检出轮虫20、56、80、3种,其中平原区的轮虫种数最多。
2) 密度与生物量。长江干流各江段轮虫密度与生物量空间分布如图2所示,密度分布范围在0.32~164.16 ind. /L,平均密度为19.66 ind. /L。高山区、峡谷区、平原区、河口区平均密度分别为9.24、27.23、19.44、0.75 ind./L,各江段中轮虫密度的最高值(S13)与最低值(S11)均出现在峡谷区。长江干流各江段生物量分布范围在8.64×10-5~7.51×10-2mg/L,平均生物量为0.11 mg/L,最高值出现在峡谷区的S13,最低值出现在S11。在各区域中,高山区、峡谷区、平原区、河口区轮虫平均生物量分别为6.46×10-3、1.54×10-2、1.38×10-2、1.44×10-4mg/L。
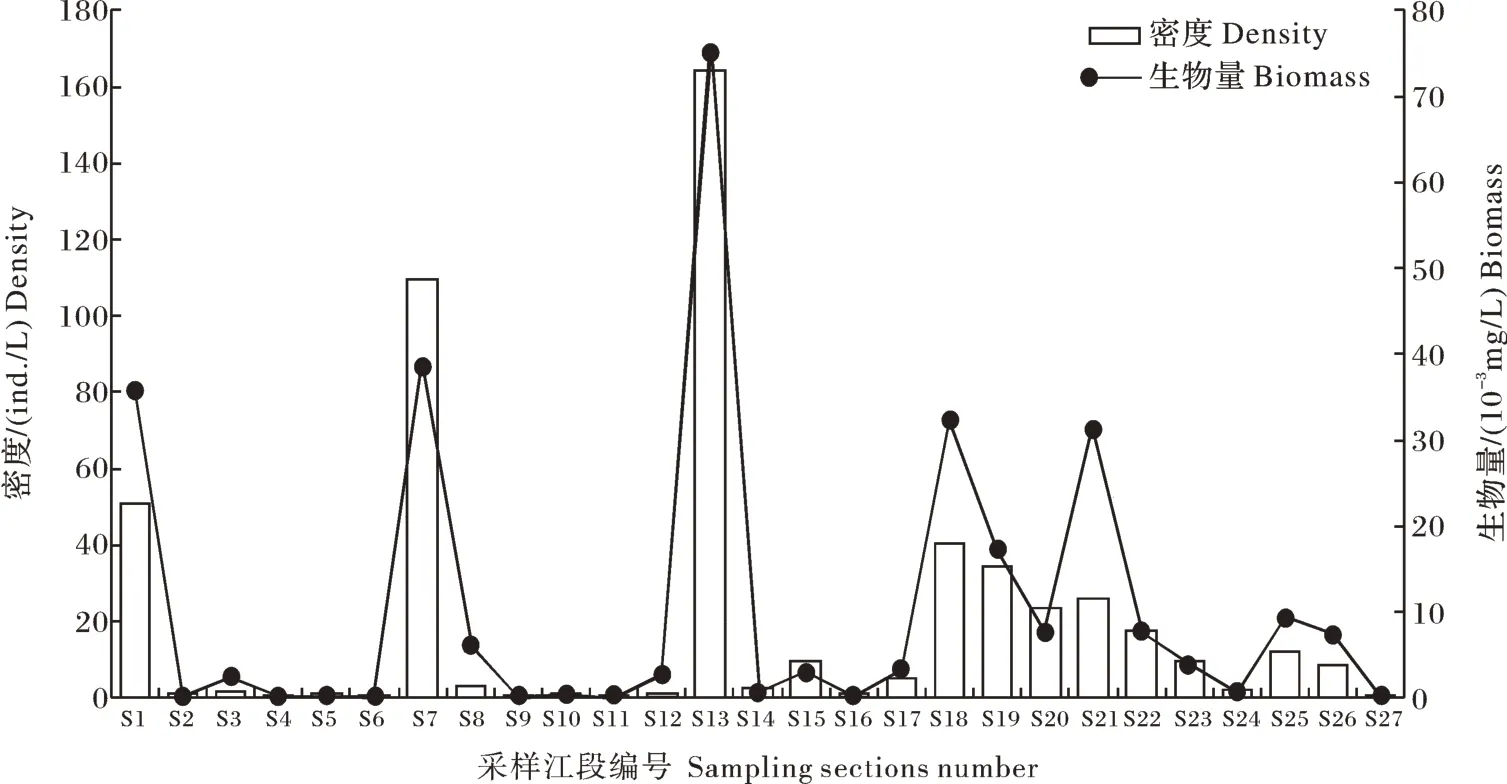
图2 长江干流各江段轮虫的密度与生物量Fig. 2 The density and biomass of rotifer in the mainstream of the Yangtze River
对长江干流各个区域轮虫密度、生物量进行差异性分析,结果显示:各区域密度(F=0.365,P>0.05)、生 物 量(F=0.280,P>0.05)差 异 不显著。
2.3 优势种与指示种
1) 优势种类。根据优势度公式计算各区域轮虫优势度,将Y≥0.02的物种视为优势种,结果如表4所示。各区域中,优势物种共12种,以臂尾轮科臂尾轮虫属为主(5种)。高山区轮虫优势种包括螺形龟甲轮虫(Keratella cochlearis)、矩形龟甲轮虫(K. quadrata)。螺形龟甲轮虫(K. cochlearis)是峡谷区最主要的优势种,曲腿龟甲轮虫(Keratella ualga)次之。平原区优势种以螺形龟甲轮虫(K. cochlearis)为主,其他依次为等刺异尾轮虫(Trichocerca similis)、曲腿龟甲轮虫(K. ualga)、角突臂尾轮虫(Brachionus angularis)、针簇多肢轮虫(Polyarthra trigla)等。河口区轮虫优势度最高者为螺形龟甲轮虫(K. cochle⁃aris),曲腿龟甲轮虫(K. ualga)次之。
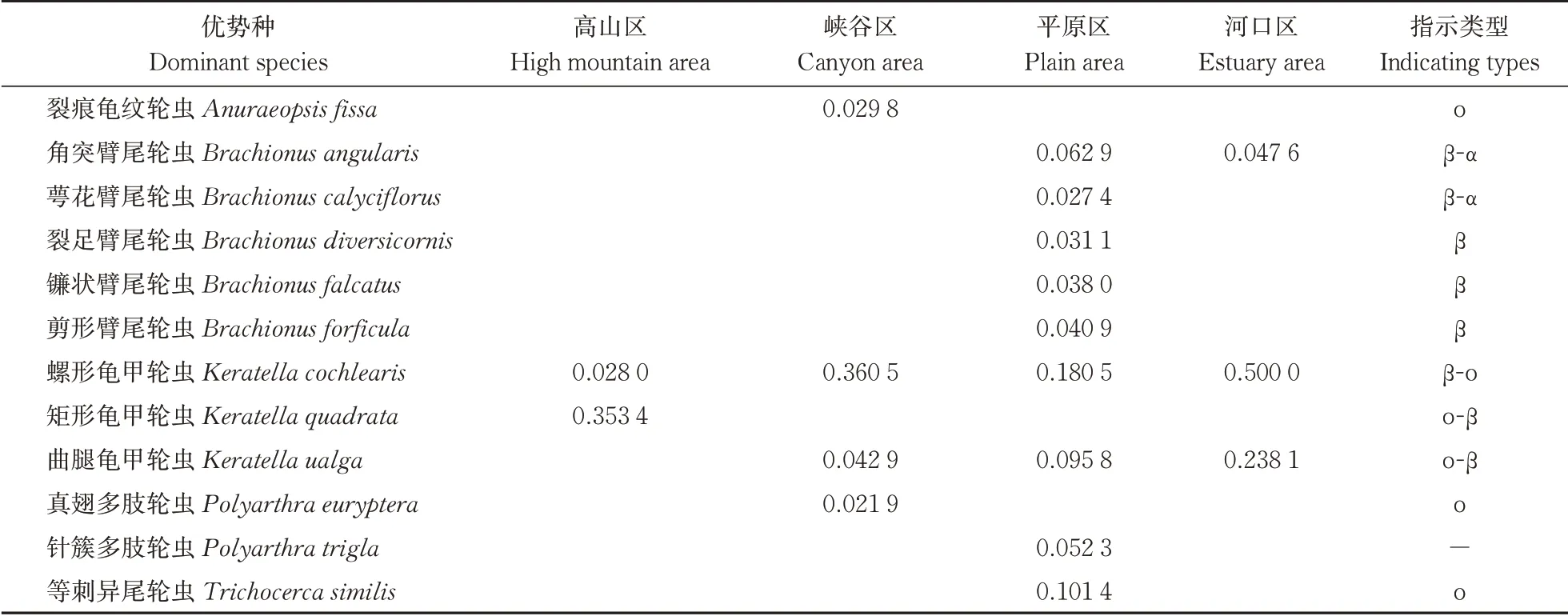
表4 长江干流各区域轮虫优势物种及优势度Table 4 The dominance (Y) and the dominant species of rotifer at each research area in the mainstream of the Yangtze River
2) 轮虫污染指示种类。共检测出污染指示种63种,其中臂尾轮科臂尾轮属种类最多(8种),鼠轮科异尾轮虫属种类数次之(6种)。所有的污染指示种包括寡污型种类23种,寡污至β-中污型17种,β-中污至寡污型2种,β-中污型15种,β-中污至α-中污型3种,α-中污型2种,多污型1种。长江干流各区域中,高山区以寡污型种类最多,占污染指示种类数的50%;峡谷区以寡污型种类最多,占40.82%,β-中污型次之,占26.53%。平原区指示种以寡污型为最多,占41.67%,寡污至β-中污型、β-中污型二者次之,占23.33%。河口区污染指示种类为寡污至β-中污型、β-中污至寡污型、β-中污至α-中污型(表5)。

表5 轮虫指示种的空间分布Table 5 Spatial distribution of rotifer indicator species
2.4 多样性指数
参考表2的评价标准对轮虫多样性进行评价,结果(图3)显示:研究期间长江干流各江段Shannonwiener值分布范围为0.65~4.19,H'指数均值为2.62(β-中污型)。其中,最低值出现在高山区的S2,最高值出现在平原区的S18。从各个区域来看,高山区、峡谷区、平原区、河口区的H'值与评价水平分别为:1.39(α-中污型)、1.89(α-中污型)、3.63(寡污型)、1.43(α-中污型)。各江段Pielou(J)指数范围在0.32~0.95,长江干流整体J均值为0.78(寡污型);对于各区域,高山区、峡谷区、平原区、河口区分别为0.80(清洁型)、0.76(寡污型)、0.76(寡污型)、0.90(清洁型)。
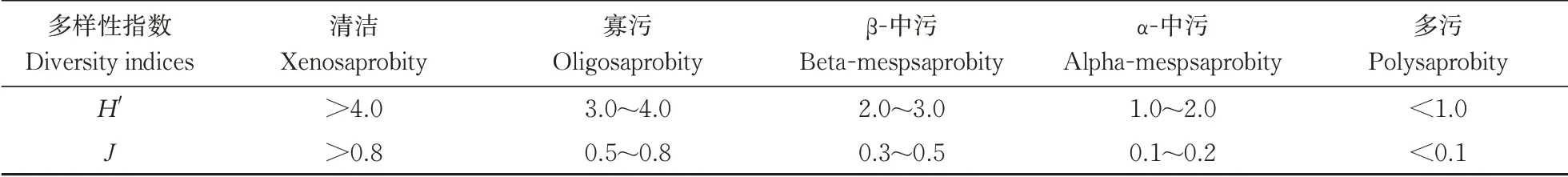
表2 基于轮虫多样性指数的水质评价标准Table 2 Criteria of water quality assessment based on the diversity indices of rotifer
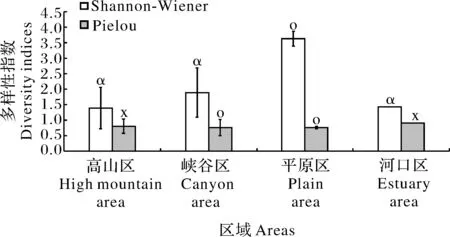
图3 长江干流各区域轮虫多样性指数及评价水平Fig. 3 The diversity indices of rotifer and evaluation levels at different research areas in the mainstream of the Yangtze River
对长江干流各个区域轮虫多样性指数进行差异性分析,结果显示,各区域之间Shannon指数(F=12.218,P<0.01)差异性极显著,Pielou指数(F=0.303,P>0.05)差异不显著。使用One-way ANO‑VA方差多重比较对Shannon-Wiener多样性指数进行分析,结果表明,高山区与峡谷区(P<0.05)、高山区与平原区(P<0.01)、峡谷区与平原区(P<0.01)均存在显著性差异。
2.5 轮虫空间分布模式
非度量多维尺度分析(NMDS)显示,峡谷区、平原区轮虫群落组成具有相似性,而其他区域间显示出明显区别(图4)。NMDS分析中的压力值(stress)为0.108,表明该二维分析具有一定的解释能力。
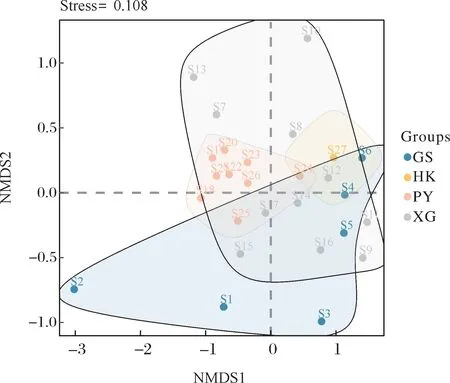
图4 非度量多维尺度分析排序结果Fig.4 Results of non-metric multidimensional scaling
2.6 轮虫群落结构与环境因子的相关性
1 )冗余分析。整合各区域共12个优势种,对其丰度进行RDA分析。长江干流RDA排序图第1轴与第2轴分别解释了轮虫变异程度的49.06%、24.14%(图5)。WT、藻密度Np(实验室未发表数据)、DO、TP是影响长江干流轮虫分布的主要影响因子。从轮虫分布来看,裂痕龟纹轮虫、萼花臂尾轮虫、裂足臂尾轮虫、镰状臂尾轮虫、剪形臂尾轮虫、等刺异尾轮虫受藻密度影响较大,与DO、TP呈负相关关系。角突臂尾轮虫分布受WT的影响较其他因素高。针簇多肢轮虫与TP呈现明显的负相关关系。曲腿龟甲轮虫与DO、藻密度无相关关系,螺形龟甲轮虫分布受DO、TP影响较高,与WT呈现显著的负相关关系。

图5 长江干流主要轮虫种类与环境因子、采样江段间的RDA排序图Fig. 5 The RDA diagram between main rotifer species and environmental parameters and sampling sections in the mainstream of the Yangtze River
2)群落结构特征与环境因子Pearson相关性分析。轮虫密度、生物量、Shannon(H')多样性指数、Pielou(J)均匀度指数与各环境参数、藻密度Np的Pearson相关性分析结果见表6。各江段轮虫密度、生物量与ORP呈显著正相关关系。多样性指数中,H’与WT、NO3--N、Np呈极显著正相关关系,与TP、DO呈显著负相关关系;J与pH呈现出显著正相关关系。寡污型种类相对密度与TN、NO3--N、NH4+-N呈显著正相关关系。
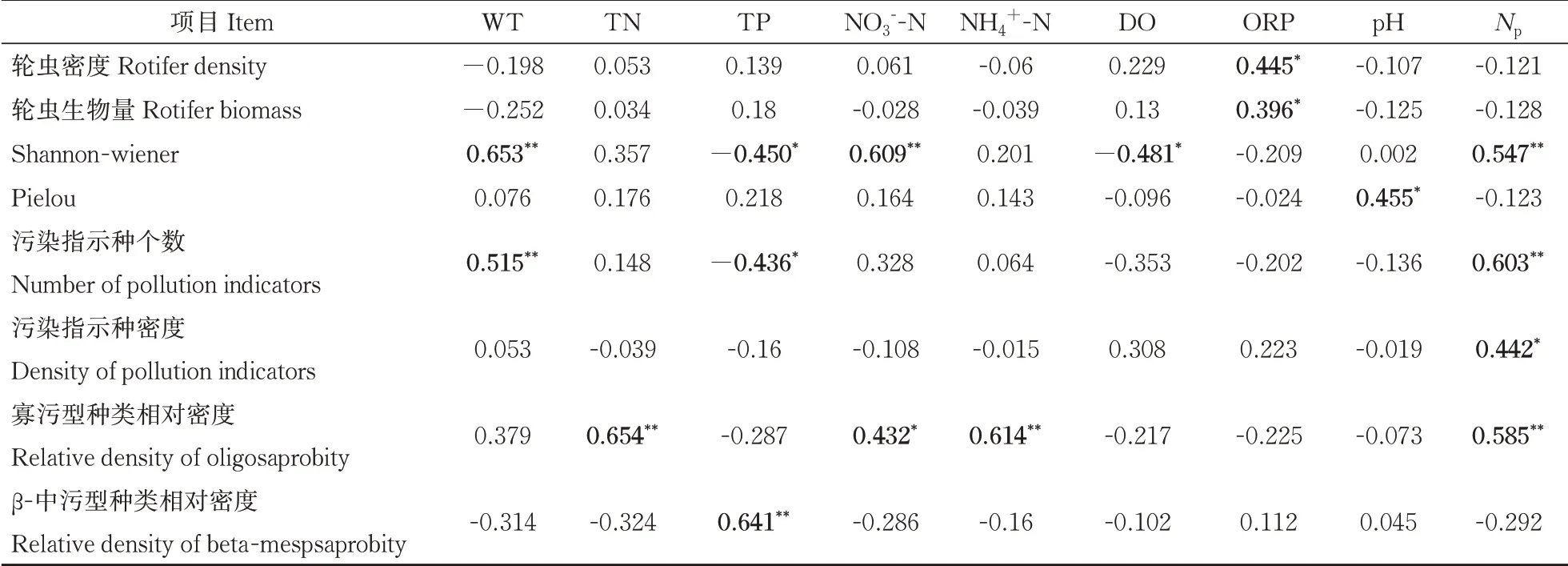
表6 轮虫群落结构特征与环境参数的相关性分析Table 6 Correlation analysis between community structure characteristics of rotifer and environmental parameters
3 讨论
多数研究揭示河流中的浮游动物以轮虫占比最高[19-20]。本研究中,臂尾轮科占轮虫种类的36.41%,该科为长江干流轮虫种类组成的主要部分,这与长江各江段已有研究结果一致[21-24]。轮虫在各种类型水域中均具有高度的适应性[25],长江干流大部分江段水流湍急,枝角类、桡足类等甲壳种类一般难以适应流动水体,这也是本研究中选择以轮虫为研究对象的原因之一。龟甲轮虫属、臂尾轮虫属种类在长江干流各个区域占据优势地位,这与罗丹等[13]在2016年对长江中游宜昌、岳阳等5个江段和郭欧阳[12]在2016-2017年对长江下游干流江段所作研究得出的优势种大体相同,表明长江干流轮虫优势种分布状况较为稳定,且龟甲轮虫属、臂尾轮虫属是长江干流水域中的广布种属。
受河流物理形态、人类干扰、河岸带等影响,长江干流存在复杂的生境类型[26]。长江干流27个采样江段呈现出规律性的空间聚集,区域之间Shan‑non-Wiener指数存在极显著差异(P<0.01),NMDS分析结果显示各区域间不同程度的差异,这表明长江干流各区域轮虫群落结构存在空间异质性。在各区域中,峡谷区、平原区平均密度、生物量、Shannon指数总体高于高山区、河口区。这可能与快速的城市化、工业化进程使武汉城市群、长三角城市群流域水质富营养化程度升高[27]有关。长江发源自青藏高原腹地,高寒且生态系统脆弱敏感,而峡谷区地理跨度大,建有大量梯级电站,过渡于四川盆地与中、下游平原[28],巨大的环境变化可能是各区域轮虫群落结构明显差异的重要原因。在高山区的6个采样江段中,S1江段呈现出较高的轮虫现存量,原因可能是S1江段的采样断面之一靠近雅西错(沱沱河左岸外流淡水湖),位于静水、流水交汇处,复杂的地形地貌与水交换使生态环境发生改变,而峡谷区的S13江段的轮虫现存量则呈现出所有采样江段中的最高水平,S13江段位于三峡大坝前,坝前缓慢的水流有助于营养盐积聚与悬浮物的沉降[6],也为轮虫的生长繁殖提供了有利条件,进而导致了S1、S13段异常高的轮虫密度与生物量。从水体理化指标来看,S7江段的营养盐指标的含量较高,水体富营养化的程度更高,因此,S7江段轮虫群落结构的异常可能与成渝城市群周围人类活动的影响有关[29]。
轮虫能对水体营养状态的变化做出快速的反应,具有污染指示功能。参照Sládeček[17]对轮虫指示种的划分,高山区优势种的污染指示水平为寡污至β-中污型,峡谷区为β-中污至寡污型,平原区介于寡污型到β-α型之间,河口区指示水平为寡污至β-中污型。不同区域指示种存在一定差异,这与长江干流水域存在着不同的生境的结果相符,表明本研究筛选出的优势物种对长江干流水质具有一定的指示作用。高山区优势种类与其他区域相差明显,具体表现为第1优势种矩形龟甲轮虫优势度高达0.35,且矩形龟甲轮虫仅在高山区占优势地位,矩形龟甲轮虫为寡污至β-中污型种类,建议当地进行水质监测及生态修复时应重点关注矩形龟甲轮虫种群的动态。而峡谷区、平原区、河口区需加强对其第1优势种螺形龟甲轮虫的监测。
以往的研究表明,多样性指数与水质呈现正相关,多样性指数越高,水质越好[30]。本研究中,高山区、峡谷区、平原区、河口区的轮虫多样性指数综合评价结果分别为β-中污、β-中污、寡污、β-中污,即平原区的水质最好,高山区水质最差。在使用传统方法对水质进行评价时,N、P等营养因子是水体理化指标中重要的部分[31],而单因素方差分析结果表明高山区在TN、NO3--N等指标均显著低于其他区域(P<0.05),表明仅用多样性指数对水质进行评价是存在误差的。这可能是由于多样性指数与物种数有关[32],物种数目越多则多样性指数越高,水质评价结果越好,但轮虫被划分为多个指示类群[17-18],因此轮虫的污染水平不能单由其数量与种类进行评定[33]。本研究中,长江干流平原区轮虫种类虽然较多,但多为污染指示种,故而仅利用多样性指数进行水质评价存在局限性。在对长江干流各区域水质生态健康进行监测评价时,将生物指标和理化指标相结合显得更为科学、准确。
水温、酸碱度、营养盐等环境因素对轮虫的分布具有重要的影响[34-35]。浮游动物对藻类的摄食是上行效应的关键环节,藻类的密度也与轮虫的生长繁殖有着密切的关系[36]。Pearson相关分析表明,ORP、WT、TP、NO3--N、DO、藻密度与长江干流轮虫多样性有显著相关性。RDA分析结果表明随着藻密度增加,多种臂尾轮虫(萼花臂尾轮虫、裂足臂尾轮虫、镰状臂尾轮虫、剪形臂尾轮虫)、裂痕龟纹轮虫、等刺异尾轮虫密度也增加;此外,TP与多种优势种密度呈现负相关关系。温度是影响轮虫生长、繁殖以及种类组成的重要因子[37]。本研究采样于6-9月,平原区、河口区的平均水温均超过25 ℃,浮游动物大量繁殖使水体中耗氧量增加,溶解氧含量减小[38],进而抑制了轮虫的生物量与多样性。水体中TP浓度的增加会导致浮游动物物种数减少[39],营养盐则通过影响浮游植物的分布来间接影响轮虫群落结构[40],这与本研究中长江干流轮虫分布与温度、藻密度、溶氧、总磷密切相关的结果一致。
浮游动物是鱼类优质的天然饵料,能通过上行效应影响鱼类群落结构,进而影响珍稀土著鱼类的摄食和分布特征[32,34],其现存量会影响渔业资源量和水生态系统整体的容纳量[30]。轮虫是淡水鱼类最主要的开口饵料[41],具有营养价值高、游动速度慢、利于仔鱼摄食等优点[42]。因此,为保护重要水生生物的繁殖及子代的孵化生长,调查轮虫的资源结构具有重要的意义[43]。本研究为长江“十年禁渔”政策实施前期的研究结果,可用于今后评估不同阶段禁渔实施效果。
