干草对羔羊瘤胃发育影响及其机制的研究进展
2023-01-17李科南杜海东娜仁花
李科南 杜海东 娜仁花
(内蒙古农业大学 动物科学学院,呼和浩特 010018)
众所周知,对于反刍动物来说,瘤胃是降解饲粮中碳水化合物的主要器官,对机体新陈代谢、免疫和健康都很重要,而饲粮中的碳水化合物降解过程依赖于瘤胃微生物的发酵作用。瘤胃中的微生物不但可将哺乳动物无法直接利用的植物纤维分解供能,而且还可为人类提供富含优质蛋白质的肉类和奶制品,但同时也会产生甲烷。因此,研究者对反刍动物瘤胃微生物组成和功能以及其影响因素,如日粮、年龄、地理位置、宿主种类等进行了较为深入的研究[1]。关于瘤胃微生物群落和微生物发酵过程的研究可追溯到19世纪中期[2]。但由于成年反刍动物瘤胃微生物群落具有较强稳定性,一些调控措施对于成熟瘤胃中微生物群落只能产生短暂的影响,一旦停止调控瘤胃微生物群落组成和发酵将会恢复到处理之前的状态[3],许多幼龄反刍动物瘤胃微生物群落组成不完善、瘤胃结构和功能不健全,通过早期干预调控幼龄阶段瘤胃微生物定植及瘤胃结构和功能发育可能会对反刍动物后期产生长期影响[4-7]。
固体饲粮(包括精料和干草)是影响幼畜瘤胃发育的重要原因之一[8],早期研究发现精料在瘤胃分解产生大量的丙酸和丁酸,有利于瘤胃乳头生长[9],而干草能量浓度较低,幼龄反刍动物瘤胃具有纤维降解功能的微生物群落建立不完善,所以不建议反刍动物断奶前补饲干草[10]。但仅饲喂精料会导致瘤胃角化不全、食糜粘附等症状[11],而在精料基础上补饲优质干草,有助于增加瘤胃容积,减少瘤胃食糜粘附,维持瘤胃乳头正常形态[8],甚至提高断奶前、后的生长性能[12]。目前虽然已有关于干草对幼龄反刍动物瘤胃发育影响及其作用机制的研究,但认识并不全面。而明晰幼龄反刍动物瘤胃发育过程是深入了解干草对幼龄反刍动物瘤胃发育影响及作用机制的基础,所以本综述以羔羊作为幼龄反刍动物模型,对羔羊瘤胃发育过程和干草对羔羊瘤胃发育影响及其作用机制的最新研究进展进行了总结,以期为干草在幼龄反刍动物饲粮中的应用提供理论依据。
1 羔羊瘤胃发育过程
羔羊出生后,瘤胃经历了非反刍阶段(从出生到21 d)、过渡阶段(21~56 d)和反刍阶段(56 d以后)的生理变化[13]。这种变化主要与瘤胃发育有关,瘤胃发育主要包括组织形态学发育(瘤胃重量和乳头的生长)、代谢功能(发酵能力和酶活性)和微生物定植(细菌、古菌、真菌和原虫等)[14]。
1.1 瘤胃组织形态学发育
瘤胃组织形态学发育主要包括瘤胃上皮发育和与此相关的瘤胃容积、瘤胃乳头长度、宽度、密度、瘤胃壁厚度、肌肉层厚度以及瘤胃壁颜色和肌肉层的变化[8]。初生绒山羊羔羊瘤胃净重占全胃净重的29.34%,56 d时瘤胃净重占全胃净重为52.53%,初生绒山羊羔羊瘤容积为9.33 mL,到56 d时为1 466.67 mL,增长了157.20倍[15]。初生绒山羊瘤胃背囊乳头高度和宽度以及瘤胃肌层厚度随日龄的增加逐步增加,其中瘤胃背囊乳头高度和肌层厚度从28 d开始采食干草后显著增厚。羔羊的角质层从14 d开始,随着采食饲草的增加,逐步增厚[15]。但不同日龄补饲开食料对羔羊瘤胃形态发育影响不同。相比于42 d补饲,7 d补饲开食料显著提高瘤胃乳头长度和宽度[16],这与早期补饲开食料增加瘤胃内挥发性脂肪酸(Volatlie fatty acids,VFA)的含量有关,尤其是VFA中的丁酸和丙酸,可以通过胰岛素、表皮生长因子(Epidermal growth factor,EGF)和胰岛素样生长因子1(Insulin-like growth factor,IGF-1)的间接反应刺激瘤胃上皮的发育[17]。
1.2 瘤胃代谢功能
1.3 瘤胃微生物群落建立
由于反刍动物生长所需的能量主要来源于瘤胃微生物对底物的发酵,并且瘤胃微生物群落的建立有利于瘤胃结构和生理功能的发育,所以深入了解瘤胃微生物的建立过程对早期调控瘤胃微生物群落组成非常重要[24]。羔羊微生物群落定植过程大致分为3个阶段[1]:最初定植阶段(出生后0~2 d),微生物主要来源于母体(包括初乳、产道和唾液)、外界环境等,如芽孢杆菌(Bacillus)和乳球杆菌属(Lactococcus)[25-26];过渡阶段(3~15 d),由初乳过渡到常乳,可利用乳中营养成分(单糖、乳糖或乳脂等)的菌群比例逐渐升高,如:拟杆菌属、乳酸杆菌属等[27]。另外,此阶段已定植细菌也是成熟瘤胃中常见的菌属,如普雷沃氏菌属和拟杆菌属;成熟阶段(28 d后),固体饲料摄入增加,可降解淀粉和纤维等微生物比例逐渐升高,并成为瘤胃中的稳定菌属,如:普雷沃氏菌属(Prevotella)、理岩菌科RC9(Rikenellaceae RC9)等[26]。此外,产甲烷菌在山羊出生后1周内开始定植,无论哪个日龄,产甲烷菌的优势属主要是甲烷短杆菌(Methanobrevibacter)、甲烷球形属(Methanosphaera)和未知甲烷菌族群C(Rumen cluster C)[28]。然而,相比与细菌和甲烷菌,厌氧真菌和原虫在幼龄反刍动物瘤胃的定植时间较晚。厌氧菌在出生后8~10 d出现,主要由新美鞭菌属(Neocallimastix)组成[29]。但是,也有研究报道,出生当天的羔羊瘤胃中便已有真菌定植[15,28]。原虫建立时间一般较其他微生物群落晚,通常可在产后15 d内发现。由于原虫主要依赖唾液进行传播,假如新生反刍动物不与成年动物接触,瘤胃原生动物就无法建立[30]。值得注意的是,虽然在采食固体饲粮之前,羔羊瘤胃微生物群落已经建立,但固体饲料的类型对瘤胃微生物群的形成非常重要。因此,早期开始采食固体饲粮的阶段可能是调控瘤胃不同生态位微生物建立的一个最佳“窗口期”[30]。
2 干草对羔羊瘤胃发育的影响
2.1 干草对羔羊瘤胃形态的影响
饲粮成分对羔羊瘤胃发育至关重要,优质干草既有助于增加瘤胃容积和重量,还可减少瘤胃上皮食糜粘附,避免高精料饲喂导致的瘤胃酸中毒、采食量降低和乳头分叉[8]。当羔羊仅饲喂精料时,由于精料颗粒度小,物理刺激清除角蛋白和死亡角质层细胞的能力弱,同时精料发酵产生大量VFA引起瘤胃液pH下降,这些结果会共同导致幼畜瘤胃上皮产生坚硬的角蛋白层,形成角化不全。角质层过厚会使瘤胃上皮血流减慢和乳头退化,影响VFA的吸收,而添加优质干草可以改善由精料引起的瘤胃上皮角化不全[8]。但干草的补饲效果受粗饲料来源、添加水平、纤维长度以及补饲时间的影响[31]。杨斌[21]发现与自由采食精料组相比,在精料基础上添加苜蓿能够减少瘤胃上皮食糜粘附及炎症反应,提高瘤胃乳头长度和肌肉层厚度;补饲苜蓿对瘤胃pH无影响,但降低了VFA的浓度,这可能与补饲苜蓿在一定程度上增加了营养物质的吸收有关。Norouzian等[32]在21日龄羔羊饲粮中添加不同比例(0、7.5%和15%)的苜蓿干草,发现添加比例为15%的苜蓿干草可降低角质层厚度,增加瘤胃壁肌肉层厚度,对羔羊生产性能无影响。为了进一步研究不同粒度大小的粗饲料对羔羊瘤胃发育的影响,Norouzian等[33]将添加比例为15%的将苜蓿切割成2 mm和3~4 cm 2个长度,发现长度为3~4 cm的苜蓿干草显著降低了瘤胃上皮角质层,增加瘤胃壁肌肉层厚度。除了干草添加水平、纤维长度对羔羊瘤胃形态发育有影响外,吕明等[34]研究发现,羔羊出生第2周开始补饲苜蓿干草瘤胃乳头和黏膜下层厚度显著高于8周龄开始补饲苜蓿干草的羔羊。此外,在评价羔羊饲粮营养价值时,中性洗涤纤维(Neutral detergent fiber,NDF)、非纤维性碳水化合物(Non fiber carbohydrate,NFC)和NDF/NFC是常用的指标。反刍动物饲粮NDF主要来自干草,所以许多研究评价了干草来源的NDF水平对羔羊的影响。解彪[35]通过在21~90日龄羔羊饲粮中添加苜蓿干草使饲粮NDF含量分别为10%、15%、20%和25%,发现提高NDF水平会提高瘤胃液pH,降低瘤胃乳头宽度和乳头角质化程度,但NDF水平对瘤胃乳头长度无影响,NDF最适添加水平为15%~25%。与解彪的研究结果类似,邓维[36]发现羔羊饲粮NDF水平为20%和25%时,有较好的增重效果。琚思思[37]研究结果表明羔羊饲粮NDF水平为16%~20%最适。
综上所述,羔羊哺乳期间在精料基础上补饲优质干草有助于减少瘤胃上皮食糜粘附及炎症反应,维持瘤胃乳头正常形态和角质层厚度,但干草补饲效果受添加水平、纤维长度以及补饲时间的影响,未来需要更细致的研究致力于探索羔羊最适的干草添加水平、纤维长度和补饲时间,并综合考虑这些因素之间的相互作用。
2.2 干草对羔羊瘤胃微生物群落的影响
干草对瘤胃微生物群落组成尤为重要,但目前有关早期补饲干草对羔羊瘤胃微生物群落的影响报道较少。吕小康[38]研究发现,与仅饲喂精料组相比,饲喂精料加苜蓿显著提高羔羊瘤胃内容物中纤维杆菌属、琥珀弧菌属、琥珀酸菌属和瘤胃球菌属的丰度,这些菌属是主要的纤维降解菌属。马万浩[12]发现在羔羊开食料中添加10%苜蓿干草显著提高瘤胃内容物中具有纤维分解功能细菌的数量以及瘤胃内容物中普雷沃氏菌属的相对丰度,但饲喂不同饲粮对育肥期羔羊瘤胃微生物无持续的影响。新生羔羊与母羊一起放牧14 d后,普雷沃氏菌属_1(Prevotella_1)、拟杆菌目_S24_7(Bacteroidales_S24_7)、瘤胃杆菌属(Ruminobacter)和月形单胞菌属(Selenomonas)的相对丰度增加[39]。以上研究结果提示在羔羊断奶前补饲干草有助于纤维分解菌的定植,但对瘤胃微生物无持续影响。此外,瘤胃上皮的微生物种类与瘤胃发育密切相关,因此受到了广泛关注[13],虽然其占瘤胃微生物总数的比例不到1%,对VFA产量的贡献也很低[40],但它们是与宿主直接相互作用的微生物,在瘤胃上皮发育中发挥作用。杨斌[21]发现,与补饲精料的羔羊相比,在精料基础上添加苜蓿诱导瘤胃上皮琥珀酸菌属(Succiniclasticum)、未分类的瘤胃球菌科(UnclassifiedRuminococcaceae)、分支杆菌属(Mogibacterium)和丁酸弧菌属(Butyrivibrio)的丰度显著增加,使瘤胃上皮在断奶前就已经适应由这些细菌引起并维持的上皮生长和代谢模式。因此,在精料基础上添加干草还可以提高瘤胃上皮微生物区系的稳定性,减少断奶应激。
2.3 干草影响羔羊瘤胃发育的分子机制
2.3.1干草影响羔羊瘤胃上皮发育
最近,利用转录组测序技术探究幼龄反刍动物瘤胃上皮细胞在基因水平变化的研究越来越多,丰富了人们对瘤胃上皮细胞分子变化机制的了解。与仅饲喂乳汁相比,在乳汁基础上添加精料可以促进瘤胃乳头的发育[41],这主要是由关键调控基因的表达驱动,如转化生长因子β1(Transforming growth factor β1,TFGB1)、叉头蛋白O1(Forkhead box protein O1,FOXO1)和过氧化物酶体增殖物激活受体α(Peroxisome proliferator-activated receptor α,PPARA)[42]。林丽梅[43]研究早期补饲开食料对羔羊瘤胃上皮生长相关基因表达和信号通路的影响,发现补饲开食料后,羔羊瘤胃中乙酸和丁酸浓度升高,其可作为信号分子上调瘤胃上皮细胞生长调节因子丝裂原活化蛋白激酶1(Mitogen-activated protein kinase 1,MAPK1)、磷脂酰肌醇-4,5-二磷酸3-激酶催化亚基亚型(Phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit beta isoform,PIK3CB)、肿瘤坏死因子配体超家族成员10(Tumor necrosis factor ligand superfamily member 10,TNFSF10)、整合素alpha 6(Integrin alpha-6,ITGA6)、Snail家族转录因子2(Snail family transcriptional repressor 2,SNAI2)、萨尔瓦多同系物1(Salvador homolog 1,SAV1)和肿瘤抑制基因(Discs large homologue l,DLG1)基因的表达,下调促凋亡家族(Bcl2-associated agonist of cell death,BAD)基因的表达,从而促进了瘤胃上皮的生长发育。干草对羔羊瘤胃发育具有重要意义,但目前关于干草刺激羔羊瘤胃上皮细胞生长、分化相关基因表达影响的研究报道有限。吕小康[38]研究不同饲粮对羔羊瘤胃腹囊上皮差异基因的影响,发现饲喂精料或者精料加苜蓿颗粒显著降低了瘤胃上皮中IGFBP4和IGFBP2的基因表达,IGFBP4和IGFBP2对IGF具有抑制作用,所以IGF的促细胞增殖作用得到充分发挥,但精料组和精料加苜蓿颗粒组细胞增殖与凋亡基因表达量无显著差异。差异表达基因富集通路分析显示,补饲精料组和精料加苜蓿颗粒组的差异基因主要富集到细胞因子-细胞因子受体相互作用、血小板活化、细胞粘附分子、系统性红斑狼疮、麻疹、NF-kB(Nuclear factor kappa-light-chain-enhancer of activated B cells,NF-kB)信号通路、IL-17信号通路等,这些信号通路与机体的免疫和炎症反应有关,提示羔羊饲喂精料加苜蓿颗粒可能影响了羔羊的免疫和炎症反应。Connor等[41]提出,干草通过抑制NF-κB活性,刺激TRIM40 (Tripartite motif containing 40) mRNA表达,保护上皮黏膜免受炎症反应。Yang等[44]发现饲喂开食料加苜蓿的羔羊瘤胃早期发育效果优于仅饲喂开食料的羔羊,苜蓿调控瘤胃发育主要通过两种主要类型的调节网络:细胞内钙信号通路以及氨基酸和脂肪酸的代谢网络。钙信号通路在断奶前瘤胃组织中上调基因涉及膜去极化和肌肉组织发育,包括小窝蛋白(Caveolin 3,CAV3)、钙离子电压门控通道蛋白α亚基1D(L-type voltage-dependent calcium channel alpha 1D subunit,CACNA1D)、SH3和富含半胱氨酸结构域的蛋白(SH3 and cysteine-rich domain-containing protein,STAC)、缝隙连接蛋白1(Gap junction protein alpha 1,GJA1)、钾离子电压门控通道互作蛋白2(Potassium voltage-gated channel-interacting protein 2,KCNIP2)、RAR相关孤儿受体A(RAR-related orphan receptor A,RORA)、PPARG共激活剂1α(PPARG coactivator 1 alpha,PPARGC1A)、一氧化氮合酶2(Nitric oxide synthase,NOS2)和蛋白激酶Cβ(Protein kinase C beta,PRKCB),下调高尔基相关分泌通路激酶FAM20C(Family with sequence similarity 20 member C,FAM20C)转录水平的表达量,加速瘤胃肌肉细胞的钙内流,促进瘤胃组织发育;瘤胃氨基酸代谢上调基因包括支链氨基酸转氨酶2(Branched-chain amino acid transaminase 2,BCAT2)、烯酰辅酶A水合酶(enoyl-CoA hydratase,ECHS1)、酰基辅酶A脱氢酶(acyl-CoA Dehydrogenase,ACADS)、羟基异丁酰辅酶A水解酶(3-hydroxyisobutyryl-CoA hydrolase,HIBCH)、羟基酰基辅酶A脱氢酶(Hydroxyacyl-CoA dehydrogenase,HADH)、醛脱氢酶1(Aldehyde oxidase 1,AOX1)、甲基丙二酰CoA变位酶(methylmalonyl-CoA mutase,MUT)和丙酰辅酶A羧化酶α/β多肽(Propionyl-CoA carboxylase α/β polypeptide,PCCA/PCCB),瘤胃脂肪酸代谢上调基因包括植烷酰辅酶A 2羟化酶(Phytanoyl-CoA 2-hydroxylase,PHYH)、溶质载体家族27(脂肪酸转运蛋白),成员2(Solute carrier family 27 member 2,SLC27A2)、乙酰辅酶A乙酰转移酶1(acetyl-CoA acetyltransferase 1,ACAT1)和3-氧酰基-(酰载体蛋白)合酶(3-oxoacyl-ACP synthase,OXSM)。
2.3.2干草影响羔羊瘤胃代谢功能
瘤胃微生物代谢产生的VFA可为反刍动物提供所需能量的60%~80%[45]。杨斌[21]比较了饲喂开食料和在开食料基础上添加苜蓿对瘤胃营养物质吸收和利用的影响,发现补饲苜蓿降低VFA的浓度可能与瘤胃上皮的吸收作用有关;此外,仅饲喂开食料的羔羊瘤胃上皮中,断奶前后NHE1、NHE2和NHE3的表达量不同程度增加,说明仅饲喂开食料的羔羊瘤胃pH可能需要通过NHE1、NHE2和NHE3来维持,而早期补饲苜蓿的羔羊NHE1、NHE2和NHE3的表达量在断奶前后不变,说明早期补饲苜蓿的羔羊瘤胃pH不需要通过其他的调节来维持。邓维[36]研究不同NDF水平的开食料(通过添加苜蓿草粉调节开食料中NDF水平)在不同日龄补饲对羔羊瘤胃上皮的VFA转运相关基因表达的影响,发现羔羊断奶前补饲不同NDF水平的开食料,影响了与VFA转运相关基因的表达;21日龄补饲,促进MCT1、NHE1和NHE3基因的表达,从而维持瘤胃内环境的稳定。此外,在犊牛上的试验也发现,断奶前补饲干草较仅饲喂精料组显著提高了瘤胃上皮MCT1的表达量[46]。
生酮作用是瘤胃成熟的标志,PPARs是调控生酮作用和瘤胃上皮细胞增殖与分化的重要转录调节因子,包括PPARA、PPARD和PPARG 3个亚型[47]。瘤胃生酮作用主要指的是丁酸生成乙酰辅酶A,然后乙酰辅酶A在HMGCS的作用下转化为3-羟基-3-甲基-戊二酸单酰辅酶A(3-hydroxy-3-methylglutaryl coenzyme A,HMG-CoA),随后再在HMGCL的作用下将HMG-CoA转化为乙酰乙酸。最后再由3-羟基丁酸脱氢酶1(3-hydroxybutyrate dehydrogenase 1,BDH1)催化生成β-羟丁酸,并进入血液循环。其中,HMG-CoA是瘤胃上皮的关键代谢物,而HMGCS是瘤胃生酮过程中的限速酶。HMGCS存在两种形式包括HMGCS1和HMGCS2,其中起主要作用的是HMGCS2,且HMGCS2的表达受到PPARs的调控[47],PPARs调控瘤胃生酮作用机制见图1。吕小康[38]研究不同饲粮对羔羊瘤胃腹囊上皮差异基因的影响,发现与代乳粉组相比,精料加苜蓿颗粒组与仅饲喂精料组显著提高了瘤胃上皮中HMGCL、HMGCS2和BDH1的表达量,从而提高瘤胃生酮和利用酮体的能力,而精料加苜蓿颗粒组与仅饲喂精料组中瘤胃腹囊上皮中HMGCL、HMGCS2和BDH1的表达无显著差异,这说明在精料基础上补饲苜蓿并不会降低瘤胃生酮和利用酮体的能力;吕小康的研究还发现在精料基础上添加苜蓿并不会提高PPARA、PPARD和PPARG的表达量,并且HMGCL和HMGCS2作为PPARs的下游靶基因,表达量显著提高,而PPARs 3个亚型在不同饲粮组之间无显著差异,说明调控瘤胃上皮细胞生酮基因表达除了PPARs之外,还有其他转录因子。Connor等[42]研究表明,在断奶时饲喂干草会激活参与能量产生的基因通路,其中雌激素相关受体α(Estrogen-related receptor α,ESRRA)可能与PPARD共同参与脂肪酸的吸收和代谢,ESRRA被认为是调控能量代谢的转录因子,是调节生长犊牛瘤胃发育和功能的潜在靶点。
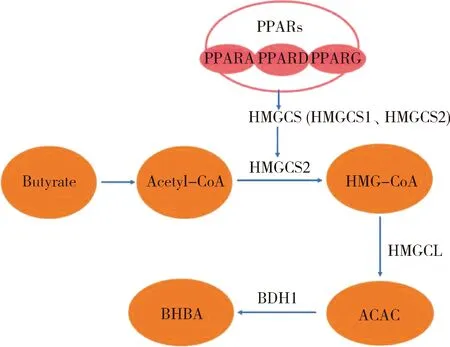
PPARs,过氧化物酶体增殖物激活受体;PPARA,过氧化物酶体增殖物激活受体α;PPARD,过氧化物酶体增殖物激活受体δ;PPARG,过氧化物酶体增殖物激活受体γ;HMGCS,3-羟基-3-甲基戊二酰辅酶A合成酶;HMG-CoA,3-羟基-3-甲基-戊二酸单酰辅酶A;HMGCL,3-羟基-3-甲基-戊二酸单酰辅酶A裂解酶;ACAC,乙酰乙酸;BDH1,3-羟基丁酸脱氢酶1;BHBA,β-羟丁酸。 PPARs, Peroxisome proliferator-activated receptors; PPARA, Peroxisome proliferator-activated receptor α; PPARD, Peroxisome proliferator-activated receptor δ; PPARG, Peroxisome proliferator-activated receptor γ; HMGCS, 3-hydroxy-3-methylglutaryl-CoAsynthase; HMG-CoA, 3-hydroxy-3-methylglutaryl coenzyme A; HMGCL, 3-Hydroxymethyl-3-methylglutaryl-CoA lyase; ACAC, Acetoacetic acid; BDH1, 3-hydroxybutyrate dehydrogenase 1; BHBA, β-hydroxybutyrate. 图1 PPARs调控生酮作用机制Fig.1 PPARs regulates the mechanism of ketogenesis
以上研究表明羔羊断奶前补饲干草对瘤胃发育有影响,这与羔羊断奶前补饲干草增强了瘤胃上皮细胞发育及VFA代谢有关(干草调节瘤胃发育的关键途径和环节见图2)。但仍需进一步评估羔羊断奶前补饲干草调控瘤胃发育的关键途径及相关主效基因,以及其在瘤胃上皮细胞发育和VFA的吸收代谢过程中的表达与参与程度需进一步验证。
3 小 结
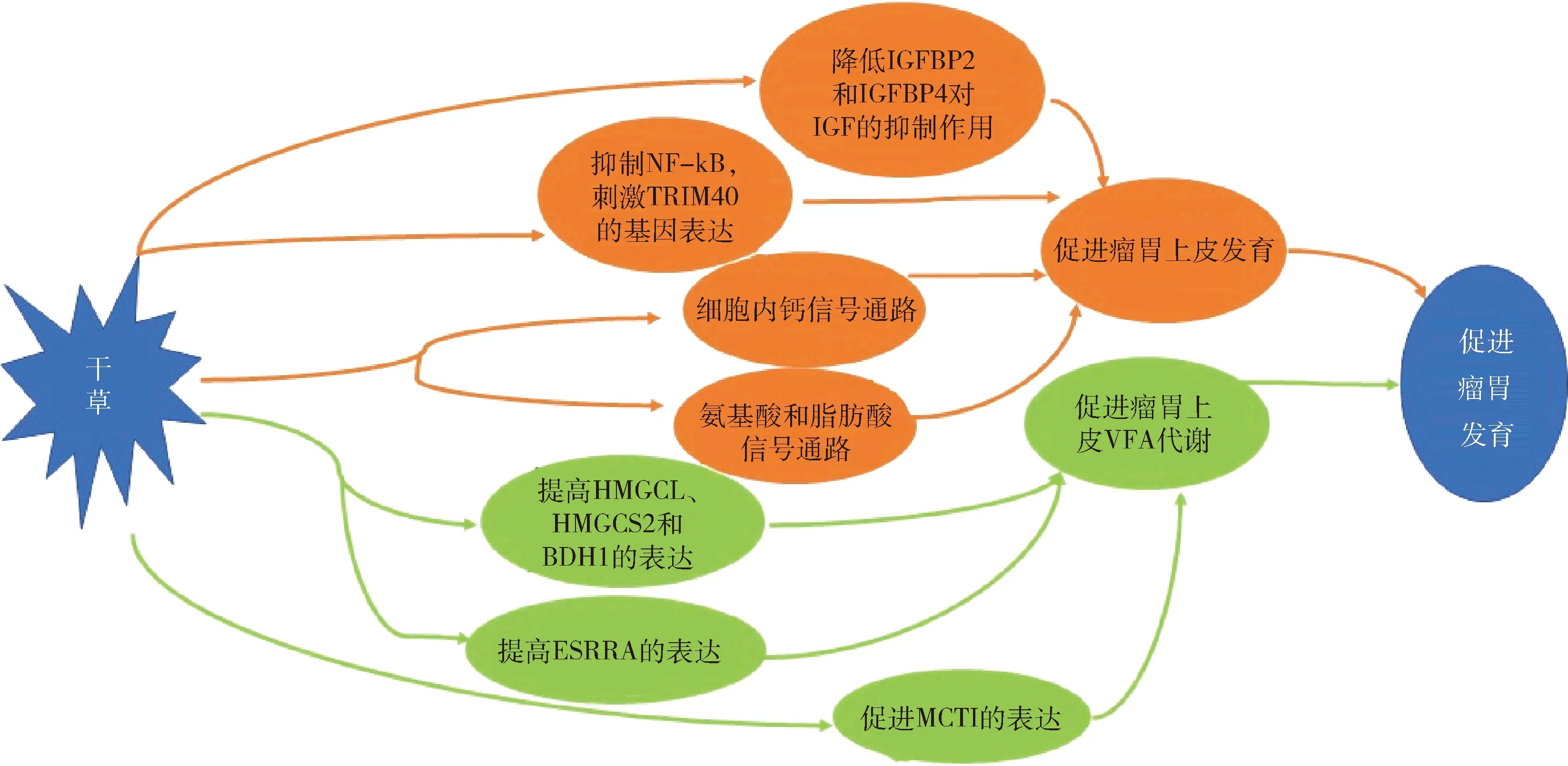
IGFBP2,胰岛素样生长因子结合蛋白2;IGFBP4,胰岛素样生长因子结合蛋白4;IGF,胰岛素样生长因子;NF-kB,活化B细胞的核因子kappa-轻链增强子;TRIM40,tripartite motif containing 40;HMGCL,3-羟基-3-甲基-戊二酸单酰辅酶A裂解酶;HMGCS2,3-羟基-3-甲基戊二酰辅酶A合成酶Ⅱ;BDH1,3-羟基丁酸脱氢酶1;ESRRA,雌激素相关受体α;MCT1,单羧酸转运载体1;VFA,挥发性脂肪酸。 IGFBP2, Insulin-like growth factor binding protein 2; IGFBP4, Insulin-like growth factor binding protein 4; IGF, Insulin-like growth factor; NF-Kb, nuclear factor kappa-light-chain-enhancer of activated B cells; TRIM40, Tripartite motif containing 40; HMGCL, 3-Hydroxymethyl-3-methylglutaryl-CoA lyase; HMGCS2, 3-hydroxy-3-methylglutaryl-CoAsynthase Ⅱ; BDH1, 3-hydroxybutyrate dehydrogenase 1; ESRRA, Estrogen-related receptor α; MCT1, Monocarboxylate transporter 1; VFA, Volatile fatty acid.图2 羔羊断奶前补饲干草调节瘤胃发育的关键途径和环节Fig.2 Key approaches and links to regulating rumen development by supplementing hay before weaning lambs
羔羊断奶前补饲干草不仅可以增加瘤胃容积,减少瘤胃食糜粘附,维持瘤胃乳头正常形态,还有助于纤维分解菌在瘤胃定植,促进瘤胃上皮细胞发育及VFA代谢。干草促进断奶前羔羊瘤胃发育的机制可能与干草调节细胞内钙信号通路及氨基酸和脂肪酸的代谢网络,降低IGFBP2和IGFBP4的基因表达量,抑制NF-kB信号通路,提高HMGCL、HMGCS2、BDH1和MCT1的基因表达量,激活ESRRA参与能量产生的基因通路有关。尽管已经明确干草补饲效果受添加水平、纤维长度以及补饲时间的影响,但未来仍需要更细致的研究致力于探索羔羊最适的干草添加水平、纤维长度和补饲时间,并综合考虑不同因素之间的相互作用。此外,干草调控断奶前羔羊瘤胃发育机理仍需进一步深入研究,针对干草调控羔羊瘤胃发育关键途径及相关主效基因,在瘤胃VFA的吸收代谢和上皮细胞发育过程中的表达与参与程度需进一步验证。
