舟山岛北部海域鱼类群落结构及其生物多样性
2023-01-14陆延牛威震程爱勇俞存根江新琴
陆延,牛威震,程爱勇,俞存根*,江新琴*
(1.国能浙江舟山发电有限公司,浙江 舟山 316012; 2.浙江海洋大学 水产学院,浙江 舟山 316022)
舟山渔场位处东海北部,水文环境受江浙沿岸水、台湾暖流和黄海冷水团的季节性交互影响,水质肥沃,饵料生物丰富,盛产大黄鱼Pseudosciaenacrocea、小黄鱼Pseudosciaenapolyactis、带鱼Trichiurushaumela和曼氏无针乌贼Sepiellamaindroni等,是中国渔业资源最丰富、生产力水平最高的渔场之一[1]。舟山海域为长江、钱塘江和甬江入海径流交汇区,域内岛屿星罗棋布,生态环境适宜,为多种经济鱼、虾和蟹类等水生生物的产卵、索饵肥育场,对东海渔业可持续发展和生态系统健康稳定具有十分重要的作用,曾是舟山渔场的主要捕捞作业区。但是,由于受过度捕捞、环境污染、栖息地破坏及气候变化等多种因素影响,自20 世纪80 年代初开始,该海域的传统渔业资源已相继衰退,生物群落结构发生更替。
为实现渔业资源可持续利用,揭示生物多样性变化趋势及生物群落结构演替规律,多年来,学者对舟山渔场及舟山岛礁附近海域鱼类资源的数量分布[1-10]、鱼卵与仔稚鱼[11-14]、群落结构与生物多样性[15-23]、生态位与种间关系[24-26]等进行了一系列研究,主要包括大小鱼山、普陀山和崎头洋附近海域等。舟山岛北部海域处在舟山本岛与岱山岛、长涂镇之间,地形呈西北-东南走向,涉及灰鳖洋、黄大洋及莲花洋,历史上盛产银鲳Stromateidesargenteus、鳓Ilishaelongata、蓝点马鲛Scomberomorusniphonius、鮸Miichthysmiiuy、曼氏无针乌贼和海蜇Rhopilemaesculenta等。但是,对该海域鱼类资源及其群落结构特征等研究至今仍属空白,且以往研究多是根据春秋两季的调查资料进行。本研究中,根据2020年10月至2021年6月四个季节在舟山岛北部海域开展渔业资源底拖网调查获得的鱼类数据,进行了鱼类群落结构及生物多样性研究,并结合2010年5月、11月在黄大洋附近海域开展的春、秋季渔业资源补充调查数据(调查方法与本次调查一致),对比分析近10 年鱼类群落结构的演替趋势及其影响因素,以期为该海域渔业资源的可持续利用提供基础数据。
1 材料与方法
1.1 研究区域及站位设置
于2020年10月(秋季)、2021年1月(冬季)、4月(春季)、6月(夏季)在舟山岛北部海域开展渔业资源底拖网调查,调查海域范围为29°59′51.46″~30°22′52.94″N,121°54′14.13″~122°33′45.76″E,共设置20个站位(图1,表1)。调查船为“浙普渔32207号”拖网渔船,主机功率为330 kW;调查网具网目规格为0.018 mm×80 mm。按照《海洋渔业资源调查规范》(SC/T 9403—2012)进行渔业资源调查,调查时每个站位拖曳1 h,拖速为3.5 kn,将每个站位拖网所获渔获物全部装入样品袋,并进行编号记录后用冰鲜保存,带回实验室进行渔获物样品的生物学测定,使用电子天平称重(精确度为0.1 g)。

本图基于自然资源部标准地图服务网站GS(2019)3333号标准地图为底图制作,底图边界无修改。The figure is based on the standard map GS(2019)3333 in the standard map service website of Ministry of Natural Resources of the People’s Republic of China, with no modifications of the boundaries in the standard map.图1 舟山岛北部海域渔业资源调查站位图Fig.1 Survey stations of fishery resources in the north sea area of Zhoushan Island

表1 舟山岛北部海域渔业资源调查站位经纬度Tab.1 Longitude and latitude of survey stations of fishery resources in the north sea area of Zhoushan Island
1.2 方法
1.2.1 优势种确定 鱼类优势种采用相对重要性指数(index of relative importance,IRI)确定,计算公式为
IRI=(N+W)×F。
(1)
其中:N为某物种的尾数占总尾数的百分比(%);W为某物种的质量占总质量的百分比(%);F为某物种出现的站位数占调查总站位数的百分比(%)。将IRI≥1 000的物种定为优势种,100≤IRI<1 000的物种定为常见种,10≤IRI<100的物种定为一般种,IRI<10的物种定为少见种[6,19-20]。
1.2.2 生物多样性分析 采用Shannon-Wiener多样性指数(H′)、Pielou均匀度指数(J)和Margalef丰富度指数(D)分析生物多样性,计算公式[27-29]为
(2)
J=H′/lnS,
(3)
D=(S-1)/lnN。
(4)
其中:S为鱼类总种数;ni为某物种的个体数;N为鱼类的总个体数。
1.2.3 群落结构相似性分析 采用聚类分析方法研究调查海域不同季节各站位鱼类群落结构的相似性,采用非度量多维标度(non-metric multi-dimensional scaling,NMDS)进行二维标度排序,用等级聚类(CLUSTER)方法分析调查海域鱼类群落结构的季节变化。以胁迫系数(stress)衡量NMDS二维标度排序结果的优劣程度。通常认为当stress<0.05时,具有较佳的代表性;当0.05≤stress<0.1时,排序结果基本可信,可以认为是一个较好排序;当0.1≤stress<0.2时,其图形具有一定解释意义[30]。Bray-Curtis相似性系数(B)计算公式为
(5)
其中:S为四个季节的鱼类种类数;xij为第i种鱼类在j站位中的渔获质量(kg);xim为第m种鱼类在j站位中的渔获质量(kg)。
鱼类适温性判定主要以相关文献[31-32]为参考;鱼类种名鉴定主要参照《中国海洋生物名录》[33]。Pearson相关性检验采用SPSS 21.0软件完成,生物多样性分析采用Primer 5.0软件完成。
2 结果与分析
2.1 种类组成及渔获量
根据2020年10月(秋季)、2021年1月(冬季)、4月(春季)和6月(夏季)4个航次渔业资源底拖网调查获得的样品,在调查海域共鉴定出鱼类50种,隶属于12目26科45属。其中,春季22种,占鱼类总种数的44.0%,夏季29种,占鱼类总种数的58.0%,秋季25种,占鱼类总种数的50.0%,冬季27种,占鱼类总种数的54.0%;在捕获的鱼类中,以鲈形目Perciformes为最多,共有18种,占鱼类总种数的36.0%,不同季节鲈形目所占比例更高,分别为春季54.5%、夏季37.9%、秋季36.0%、冬季44.4%;其次为鲱形目Clupeapallasi,共有8种,占16.0%,不同季节鲱形目所占比例分别为春季22.7%、夏季20.7%、秋季28.0%、冬季22.2%。
从表2可见:调查海域不同季节鱼类种类组成差别较大,四季共有种较少,仅13种(占鱼类总种数的26%),分别是康氏小公鱼Stolephorascommersoni、黄鲫Setipinnataty、刀鲚Coiliaectenes、凤鲚C.mystus、龙头鱼Harpodonnehereus、棘头梅童鱼Collichthyslucidus、带鱼Trichiurushaumela、银鲳Pampusargenteus、髭缟虾虎鱼Tridentigerbarbatus、六丝钝尾虾虎鱼Amblychaeturichthyshexanema、红鳗虾虎鱼Taenioidesrubicundus、孔虾虎鱼Trypauchenvagina和半滑舌鳎Cynoglossussemilaevis;只在一个季节出现的鱼类有26种,占52.0%。这表明,调查海域鱼类以洄游性种类的季节性分布为主。

表2 调查海域不同季节鱼类种类及生态类型Tab.2 Fish species and ecological type in survey sea area in different seasons
四个季节的渔获量调查数据分析显示,春、夏、秋、冬季鱼类渔获量分别为20.610、55.269、14.323、10.725 kg,总渔获量为100.927 kg,占拖网总渔获量的75.43%。从每小时渔获量(以下简称渔获率)分析,以龙头鱼、银鲳、蓝点马鲛、带鱼和鮸占比为最高,分别占26.8%、17.4%、15.0%、7.0%和6.6%,合计高达72.8%;从不同季节的渔获率分析,春季渔获率较高的为蓝点马鲛和龙头鱼,分别占73.2%和14.0%,夏、秋季渔获率较高的均为银鲳和龙头鱼,其中,夏季各占26.3%和23.7%,秋季各占12.4%和73.5%,冬季渔获率较高的为凤鲚和鮸,分别占45.28%和29.15%。这表明,不同季节鱼类群体数量较多的只有少数几种鱼类,而大多数鱼类群体数量均较小。
2.2 区系特征及生态类群
2.2.1 区系特征 调查海域的鱼类种类由暖水性种(占56.0%)和暖温性种(占44.0%)组成(表2),未出现冷温性和冷水性种。鱼类区系属亚热带性质印度-西太平洋区的中-日亚区,该结果与东海区渔业资源和调查规划[32]结论相一致。
2.2.2 生态类群 根据鱼类的适温性及栖所类型,可以将调查海域捕获的鱼类分为以下3大生态类群。
河口性鱼类:9种,占鱼类总种数的18%。主要包括刀鲚、凤鲚、鲻Mugilcephalus、四指马鲅Eleutheronematetradactylum、髭缟虾虎鱼、斑尾刺虾虎鱼Acanthogobiusommaturus、矛尾虾虎鱼Chaeturichthysstigmatias、红鳗虾虎鱼和暗纹东方鲀Takifugufasciatus等。
沿岸性鱼类:16种,占鱼类总种数的32%。主要包括斑鰶Konosiruspunctatus、康氏小公鱼、赤鼻棱鳀Thrissakammakensis、龙头鱼和长鳍底灯鱼Benthosemapterotum等。
近海性鱼类:25种,占鱼类总种数的50%。主要包括海鳗Muraenesoxcinereus、中华小沙丁鱼Sardinellanymphaea、鳓Ilishaelongata、黄鲫、黑鮟鱇Lophiomussetigerus、日本下鱵鱼Hyporhamphussajori、尖海龙Syngnathusacus、翼红娘鱼Lepidotriglaalata和短鳍红娘鱼Lepidotriglamicropterus等。
2.3 优势种及常见种
从表3可见:调查海域不同季节鱼类,春季优势种有2种,分别为斑尾刺虾虎鱼和龙头鱼,常见种有3种,分别为棘头梅童鱼、蓝点马鲛和银鲳;夏季优势种有6种,分别为康氏小公鱼、棘头梅童鱼、龙头鱼、斑尾刺虾虎鱼、带鱼和银鲳,常见种有1种,为黄鲫;秋季优势种有1种,为龙头鱼,常见种有2种,分别为银鲳和中华小沙丁鱼;冬季优势种有1种,为凤鲚,常见种有4种,分别为龙头鱼、刀鲚、鮸和红鳗虾虎鱼。四季无共有优势种,龙头鱼除冬季外,为3个季节的优势种,斑尾刺虾虎鱼仅在春、夏季出现,是这两个季节的优势种。另外,除斑尾刺虾虎鱼外,康氏小公鱼、凤鲚、龙头鱼、棘头梅童鱼、带鱼和银鲳6个优势种均为四个季节的共有种。
2.4 生物多样性特征
2.4.1 季节变化 从表4可见,H′指数、J指数和D指数的季节变化趋势不同,其中,H′指数为夏季>秋季>春季>冬季,J指数为春季=夏季>秋季>冬季,D指数则为冬季>秋季>夏季>春季。

表3 调查海域不同季节鱼类优势种和常见种Tab.3 Dominant and common species of fish in survey sea area in different seasons
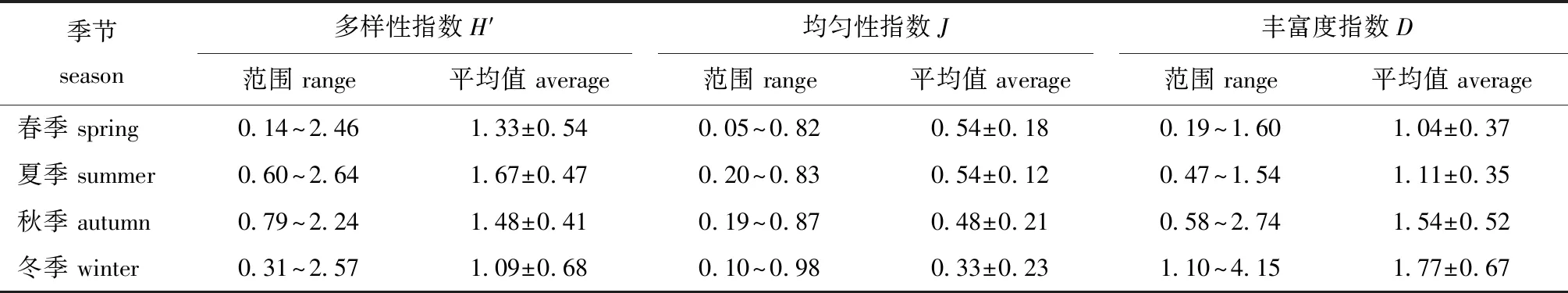
表4 调查海域不同季节鱼类生物多样性指数分布范围Tab.4 Distribution range of fish biodiversity index in the survey area in different seasons
2.4.2 空间分布 从表5可见:春季,H′指数以调查海域的中部和东部为较高,西南部较低,最高值出现在17号站,最低值出现在40号站;J指数则以调查海域的西部为较高,中部和东部为较低,最高值也出现在17号站,最低值出现在21和40号站;D指数以调查海域东部为较高,西南部为较低,最高值出现在48号站,最低值出现在40号站。
夏季,H′指数和J指数均以调查海域的东南部为较高,东北部为最低,其中,H′指数和J指数最高值均出现在48号站,最低值均出现在35号站;D指数以调查海域的中部为较高,东南部为较低,最高值出现在16号站,最低值出现在36号站。
秋季,H′指数以舟山岛与秀山岛之间海域较高且分布较集中,明显高于其他海域,而在海域东部较低,最高值出现在34号站,最低值出现在35号站;J指数也以调查海域中部为较高,主要分布在舟山岛与秀山岛之间海域,而其他海域均较低,最高值出现在24和34号站,最低值出现在35号站;D指数以调查海域中部偏北为较高,西北部较低,最高值出现在8号站,最低值出现在46号站。
冬季,H′指数和J指数相似,均以莲花洋附近海域为较高,除此之外,其他大部分海域均较低,其中,H′指数最高值出现在24号站,最低值出现在18号站;J指数最高值出现在31号站,最低值出现在18和34号站;D指数以调查海域的最东端为较高,而在舟山岛与秀山岛之间海域为较低,最高值出现在46号站,最低值现在18号站。
2.5 聚类及排序结果
调查海域鱼类群落CLUSTER等级聚类图和NMDS非度量多维标度分析如图2、图3所示。根据计算与对比分析,聚类分析和NMDS分析的结果较为一致。从等级聚类分析可以看出,四个季节鱼类群落总体可以划分为4~6个组群。其中,春季在50%相似性水平上可以分为4个组群,分别为第一组群(17和42号站)、第二组群(21和40号站)、第三组群(33、14、24、5、8和13号站)和第四组群(18、26、35、30、31、34、46、48、16和36号站),代表性鱼类为龙头鱼、银鲳和蓝点马鲛;夏季,在55%相似性水平上可以分为4个组群,分别为第一组群(35号站)、第二组群(46和48号站)、第三组群(36、8、26、16、24、5和18号站)和第四组群(34、17、21、33、40、31、13、30、14和42号站),代表性鱼类为龙头鱼、银鲳和带鱼;秋季,在43%相似性水平上可以分为5个组群,分别为第一组群(46号站)、第二组群(33号站)、第三组群(31、21、42、40、14和30号站)、第四组群(36、48、8、16和35号站)和第五组群(13、17、26、5、18、24和34号站),代表性鱼类为龙头鱼、银鲳和黄鲫;冬季,在35%相似性水平上可以分为6个组群,分别为第一组群(8号站)、第二组群(16号站)、第三组群(24号站)、第四组群(18号站)、第五组群(46和48号站)和第六组群(42、13、40、14、21、31、33、26、36、5、30、34、17和35号站),代表性鱼类为鮸、凤鲚和刀鲚(图2)。

表5 调查海域各站位各季节鱼类生物多样性指数Tab.5 Fish biodiversity index at various sampling sites and seasons in the survey area

图2 鱼类群落CLUSTER等级聚类分析图Fig.2 CLUSTER analysis of fish community structure

图3 鱼类群落结构NMDS排序图Fig.3 NMDS ordination chart of fish community structure
鱼类群落相似性程度以夏季最高,冬季最低,四个季节鱼类群落主要以龙头鱼和银鲳等为代表,但不同群落的代表性鱼类各不相同。由NMDS排序结果可知,调查海域春、夏、秋、冬季的胁迫系数分别为0.21、0.12、0.18和0.17(图3)。其中,夏、秋季和冬季stress均为0.1~0.2,表示此结果具有一定的解释意义。
3 讨论
3.1 鱼类种类组成分析
本次调查中,共捕获并鉴定出鱼类50种,从鱼类适温性来看,主要以暖水性种和暖温性种为主,这与成庆泰等[34]1964年的调查结果相一致,也反映了调查海域的水文环境及鱼类区系具有亚热带海区的特点。本次调查显示,不同季节鱼类种类数不同,以夏季的鱼类种类数最多(29种)、春季最少(22种),不同季节的鱼类种类组成也不同,四季共有种有13种,仅占26%,而只在一个季节出现的有26种,占52.0%,表明调查海域的鱼类以洄游性种类的季节性分布为主。从鱼类适温和生态习性上来看,四季暖水性鱼类种数与暖温性鱼类种数比例分别为春季59%∶41%、夏季52%∶48%、秋季64%∶36%和冬季56%∶44%;四季河口性、沿岸性和近海性生态类群鱼类种数比例分别为春季27%∶32%∶41%、夏季17%∶28%∶55%、秋季24%∶32%∶44%和冬季19%∶48%∶33%。可以看出,夏季暖水性鱼类种数与暖温性鱼类种数占比相差最小,而近海性鱼类种数占比最高,可能是因为夏季随着调查海域的水温升高,不同适温类型的鱼类,特别是一些原来分布在相对偏外侧的近海性鱼类洄游或迁徙到此生栖、产卵,而到了秋、冬季,随着水温下降,近海性暖温性鱼类向外侧深水处进行越冬洄游,故其占比在调查海域逐渐降低。与2010年春、秋季在舟山黄大洋海域的调查结果[3]相比,两次调查所获得的鱼类种数比较接近,分别为48种[3]、50种。种类组成中鲈形目相差较小,但2010年捕获的一些软骨鱼类在本次调查中均未有捕获,这可能与调查站位和时间略有差异有关。
3.2 优势种特征分析
优势种是指群落中占优势的种类,它包括群落结构中数量上最多、体积上最大、对生境影响最大的种类。本次调查中,从鱼类生态习性来看,调查海域四季鱼类优势种均以沿岸性与河口性种类为主,但不同季节的鱼类优势种更替明显。龙头鱼是该调查海域的最重要的优势种,四季的IRI值均较高,其中在春、夏、秋季为优势种,特别是秋季,IRI值高达8 196.93,冬季该鱼虽不是优势种,但IRI值也高达852.07。夏季优势种数量最多,有康氏小公鱼、棘头梅童鱼、龙头鱼、斑尾刺虾虎鱼、带鱼和银鲳6种,且棘头梅童鱼、龙头鱼、带鱼和银鲳4种是经济鱼类,夏季还是唯一有近海性鱼类成为调查海域优势种的季节,说明夏季调查海域是众多不同适温类型和生态习性鱼类的生栖场所。2015—2016年郭朋军等[20]于春、秋季在舟山沿岸渔场开展底拖网渔业资源调查时发现,春季鱼类优势种为六丝钝尾虾虎鱼,秋季鱼类优势种为龙头鱼。本次调查中,秋季海域鱼类优势种与郭朋军等[20]的调查结果一致,而春季则不同。2006—2007年俞存根[1]在舟山渔场开展四季拖网渔业资源调查时发现,鱼类优势种为日本红娘鱼Lepidotriglajaponice、绿鳍鱼Chelidonichthyskumu、六丝钝尾虾虎鱼、细条天竺鲷Apogonlineatus、海鳗、黑鮟鱇、星康吉鳗Congermyriaster、小黄鱼、前肛鳗Dysommaanguillaris、短吻舌鳎Cynoglossusabbreviatus、棘头梅童鱼、刺鲳Psenopsisanomala、龙头鱼、多棘腔吻鳕Coelorhynchusmultispinulosus、褐斑三线舌鳎ynoglossustrigrammus和角木叶鲽Pleuronichthyscornutus等。与俞存根[1]的研究相比,本次调查海域的鱼类优势种数量大大减少,且具有较大差异。这可能有两方面的因素,一是调查海域不同,虽然均在舟山渔场内,但2006—2007年的调查范围涉及整个舟山渔场范围,而本次调查范围较小,仅局限在舟山岛与岱山岛、长涂岛中间海域,不同的海域范围必然导致鱼类优势种类组成不同;二是随着时间的推移,因捕捞与环境污染等人类活动干扰及全球气候变化,可能出现调查海域的优势种类演替现象。与对周边海域的研究结果相比较可以发现,本次调查所获得的鱼类优势种与郑献之等[3]在舟山黄大洋海域春、秋季均以龙头鱼为优势种的研究结论基本相吻合;与苗露等[19]在普陀山附近海域的研究结论比较,秋季鱼类优势种一致,均以龙头鱼为优势种,但春季不同。引起上述现象的具体原因有待进一步研究。综合各文献报道[1-3,5-8,10,19-20,22],秋季,龙头鱼是舟山渔场各个海域最重要的优势种之一,其他优势种类则随地点和季节改变而发生变化。
3.3 生物多样性分析
生物多样性是衡量调查海域鱼类生态环境质量的重要标志。本次调查中,四个季节鱼类群落结构中仅有春季stress>0.2,原因可能是在春季调查中,捕获到少数大体型的蓝点马鲛,其质量在整个季节的鱼类质量中占比过大,从而导致胁迫系数过大。夏季作为群落结构相似性最高的季节,胁迫系数接近0.1,说明群落结构最为稳定,聚类分析效果最好,这一方面与夏季渔获量最多、多样性指数最高有关;另一方面,夏季出现6种优势种且优势度水平基本相差不大,对于夏季群落结构的稳定性起到了至关重要的作用。同时生物多样性的变化及群落结构特征的划分等可能还会受到温度或盐度等环境因子的影响,是否还会有更多的影响因素存在,仍值得进一步研究分析。
本次调查中,鱼类群落多样性指数季节差异较大,多样性指数以夏季最高,均匀度指数以春季最高,丰富度指数以冬季最高。其原因可能是夏季随着台湾暖流势力增强,调查海域水温上升,大量鱼类洄游到该海域进行生殖产卵活动,因此,生物多样性较高;春季近海性种类陆续向调查海域移动,出现近海性种类、沿岸性种类和河口性种类的鱼类数和个体数比较均衡的现象;冬季丰富度指数高则可能是群体数量比较大的近海性种类随着水温下降,离开调查海域洄游至外侧深水区越冬,导致调查海域渔获的鱼类总个体数下降形成的现象。不同区域的环境条件及干扰因素直接影响生物多样性指数,从调查海域生物多样性空间分布规律来看,夏、秋季多样性指数和均匀度指数最低值集中分布在黄大洋中部海域的35号站,丰富度指数最低值分布在舟山黄大洋中部和北部海域的46号站;冬、春季多样性指数、均匀度指数及丰富度指数最低值多出现在舟山岛与秀山岛之间的近岸区(21、18、16、8号站及大小鱼山南部的40号站)。原因可能是夏、秋季大量近海性种类分布在该区域产卵和索饵,造成优势种种类数较多且群体数量较大;冬、春季情况可能是因为靠近近岸区,受人类活动干扰大及环境污染因素导致的。是否还有其他影响因素,仍有待进一步调查、监测和分析。
4 结论
1)舟山岛北部近岸海域鱼类资源以夏季多样性水平最高,鱼类种类数最多且略高于附近海域。
2)该海域以洄游性鱼类种类数占比较大,沿岸性的本地种虽占比较少但常作为优势种而出现。
3)四季中以夏季群落结构最为稳定,若出现个体较大或尾数较多的鱼类,则会突出影响群落结构稳定性及生物多样性水平。
