抱茎金花茶(Camellia tienii)的叶绿体基因组特征分析
2023-01-10丁祥青毕远洋陈佳婷练芳松李文芳吴丽君邹双全
丁祥青, 毕远洋, 陈佳婷, 向 双, 练芳松, 李文芳, 吴丽君, 邹双全
(1.福建农林大学园林学院,福建福州 350002; 2.福建农林大学林学院,福建福州 350002;3.福建省洋口国有林场,福建南平 353001; 4.福建省林业科学研究院,福建福州 350012)
金花茶隶属于山茶科(Theaceae)山茶属(Camellia)金花茶组(sect.ChrysanthaChang)植物,是山茶花中唯一的珍稀金黄色花瓣类型[1]。花开时呈杯、壶或碗状,花瓣呈亮丽的金黄色,十分夺目,花瓣蜡质似半透明,晶莹且油润,观赏价值极高,被誉为“植物界大熊猫”“幻想中的黄色山茶”,被列为我国一级或二级保护植物[2]。同时,金花茶的叶、花等器官中含有多种活性成分,如多酚、多糖、黄酮类、皂苷、氨基酸等,在食品或中药中发挥重要作用。金花茶植物主要分布在我国南部亚热带季风气候区和越南北部热带季风区,属分布极为狭窄的植物种群,其狭窄栖息地的气候差异大,导致金花茶的花、叶形态变异丰富,迄今为止已发掘金花茶植物42种5变种[3]。抱茎金花茶也称越南黄抱茎金花茶,是一种重要的金花茶植物,因其叶子和茎相抱而得名,其造型独特,花型优美,近年来在园林和园艺界颇受关注。
叶绿体基因组起源于古老的诱捕事件,原始的真核生物捕获并吞噬了古蓝藻成为内共生体,在漫长的进化过程中,蓝藻基因组由于大量基因转移到真核生物核基因组中导致其在遗传上依赖于真核生物而成为一质体基因组,真核生物也因此从异养转变为自养。植物叶绿体基因组(chloroplast DNA,简称cpDNA)因进化极为保守通常为典型的环状双链四分体结构[4-6],具有1个大单拷贝区(large single copy,简称LSC)、1个小单拷贝区(small single copy,简称SSC)和1对反向重复区(inverted repeat,简称IR)。被子植物叶绿体基因组通常基因结构、基因顺序和含量高度保守,加上结构简单、分子量小,具有易解析、极少发生重组和变异、母系遗传、进化路线独立等特点,在物种鉴定和亲缘关系研究等方面具有独特应用价值和发展前景[7-8]。
前人对金花茶组植物的分类大都基于表型或者简单的分子标记,表型分类易受环境影响,片段序列分子标记不能完全反映DNA相应部位的碱基变异。此外,这些分析所涉及物种不够全面,使得金花茶植物鉴定困难、亲缘关系紊乱,给黄色山茶花种质创新带来巨大困难。现今山茶属系统发育关系尚存争议,虽然此前有2项重要研究[9-10]揭示了部分山茶属植物的叶绿体基因组,为山茶属系统发育关系研究提供了证据,但山茶属系统发育的重建仍缺乏大量信息[11]。本研究首次利用Illumina测序数据对抱茎金花茶叶绿体全基因组进行了表征,抱茎金花茶叶绿体基因组序列的揭示将为山茶属系统发育、金花茶植物分子鉴定等研究提供大量信息位点。
1 材料与方法
1.1 文库构建和测序
本研究试验材料2020年采集于福建省洋口国有林场(117°48′54.18″E,26°47′30.16″N)。采用改良的十六烷基三甲溴化铵法(cetylrimethylammonium bromide,简称CTAB)从抱茎金花茶嫩叶中提取DNA,用琼脂糖凝胶电泳和分光光度计检测DNA的浓度和质量,将高质量的DNA使用Covaris仪超声波打断,通过末端修复、DNA片段3′加A、连接接头处理后,选取270 bp的DNA片段进行PCR扩增,对扩增产物进行引物二聚体去除后,用于构建文库。最后将质量检测合格的文库在Illumina平台进行测序。
1.2 序列拼接及注释
SOAPnuke 1.6.5 (https://github.com/BGI-flexlab/SOAPnuke) 用于过滤原始数据中的低质量序列,过滤参数为-n 0.05-l 20-q 0.4-i-Q 2-G-M 2-A 0.5-d。本研究共获得了19.03 Gb clean data。用 GetOrganelle 软件[12]对clean data进行自动化组装。组装完成后用Bandage 软件可视化,去除多余contig并编辑成环状,环状序列即为抱茎金花茶叶绿体全基因组序列。以凹脉金花茶(Camelliaimpressinervis)(NC022461)的全叶绿体基因组为参考,用 Plastid Genome Annotator软件[13]对抱茎金花茶叶绿体全基因组序列进行预测基因注释。用 Geneious 软件可视化注释结果并人工检查编码基因的起始密码子和终止密码子,得到抱茎金花茶全叶绿体基因组的最终注释结果GenBank文件。组装注释好的抱茎金花茶叶绿体全基因组fasta文件和GenBank文件提交至 NCBI GenBank数据库(访问号为OL435568),用Organellar Genome DRAW (OGDRAW) 软件可视化抱茎金花茶叶绿体全基因组。
1.3 金花茶植物叶绿体基因组比较分析
从NCBI GenBank数据库下载山茶(Camelliajaponica)等5个山茶属植物的叶绿体基因组数据,用在线网站IRscope(https://irscope.shinyapps.io/irapp/)进行IR边界可视化分析。 从NCBI GenBank数据库下载贵州金花茶(Camelliahuana)等9个金花茶植物的叶绿体基因组数据(其中显脉金花茶Camelliaeuphlebia叶绿体基因组数据为笔者所在课题组测序组装注释并上传),用在线网站mVISTA(http://genome.lbl.gov/vista/mvista/submit.shtml)进行叶绿体基因组比对可视化分析。使用本地mafft进行基因序列比对。
1.4 重复序列及SSR鉴定
用在线软件REPuter鉴定抱茎金花茶cpDNA中的重复序列,包括正向重复(F)、反向重复(R)、互补重复(C)和回文重复(P),最小重复设置为 30 bp,最小重复序列长度距离设置为3。用在线网站MISA(https://webblast.ipk-gatersleben.de/misa/)进行简单重复序列(simple sequence repeat,简称SSR)识别,将单核苷酸、二核苷酸、三核苷酸、四核苷酸、五核苷酸、六核苷酸的最小重复数分别设置为8、5、4、3、3、3,2个SSR之间的最小距离设置为100 bp。
1.5 系统发育树构建
从NCBI GenBank数据库下载大头茶(Polysporaaxillaris)、长果大头茶(Polysporalongicarpa)和18个金花茶植物的叶绿体基因组序列。用本地MAFFT进行多序列比对,以大头茶(Polysporaaxillaris)和长果大头茶(Polysporalongicarpa)为外类群用在线网站CIPRES(http://www.phylo.org/)的RAxML-HPC2 on XSEDE构建系统发育树,用 Bootstrap method 进行自展值检验,设置1 000次重复。
2 结果与分析
2.1 抱茎金花茶叶绿体基因组基本特征
抱茎金花茶叶绿体基因组为共价闭合的双链环状分子,全长156 591 bp,LSC为86 172 bp、 SSC为18 275 bp、IR为26 072 bp(图1)。抱茎金花茶叶绿体基因组中共预测注释到134个基因(表 1),包括88个蛋白编码基因,38个 tRNA 基因和8 个 rRNA 基因。88个蛋白编码基因中包含29个与自我复制有关的基因,45个与光合作用有关的基因,6个其他编码蛋白质的基因和8个功能未知的基因。134个基因中有95个基因是独特的,其余39 个基因位于反向重复区,位于反向重复区的基因包含17 个蛋白编码基因(1 个rps19、1个ycf1,1个ycf68,2个rpl2、2 个rpl23、2个ycf2、2 个ycf15、2个ndhB、2 个rps7、2个rps12),14 个 tRNA 基因(2 个trnI-CAU、2 个trnL-CAA、2 个trnV-GAC、2 个trnL-GAU、2 个trnA-UGC、2 个trnR-ACG、2 个trnN-GUU)和 8 个rRNA 基因(2 个rrn16、2 个rrn23、2个rrn4.5、2个rrn5)。抱茎金花茶叶绿体基因组中有16个基因含有1个内含子,2个(clpP、ycf3)基因含有2个内含子。抱茎金花茶全叶绿体基因组GC 含量为37.3%,AT 含量为62.7%。

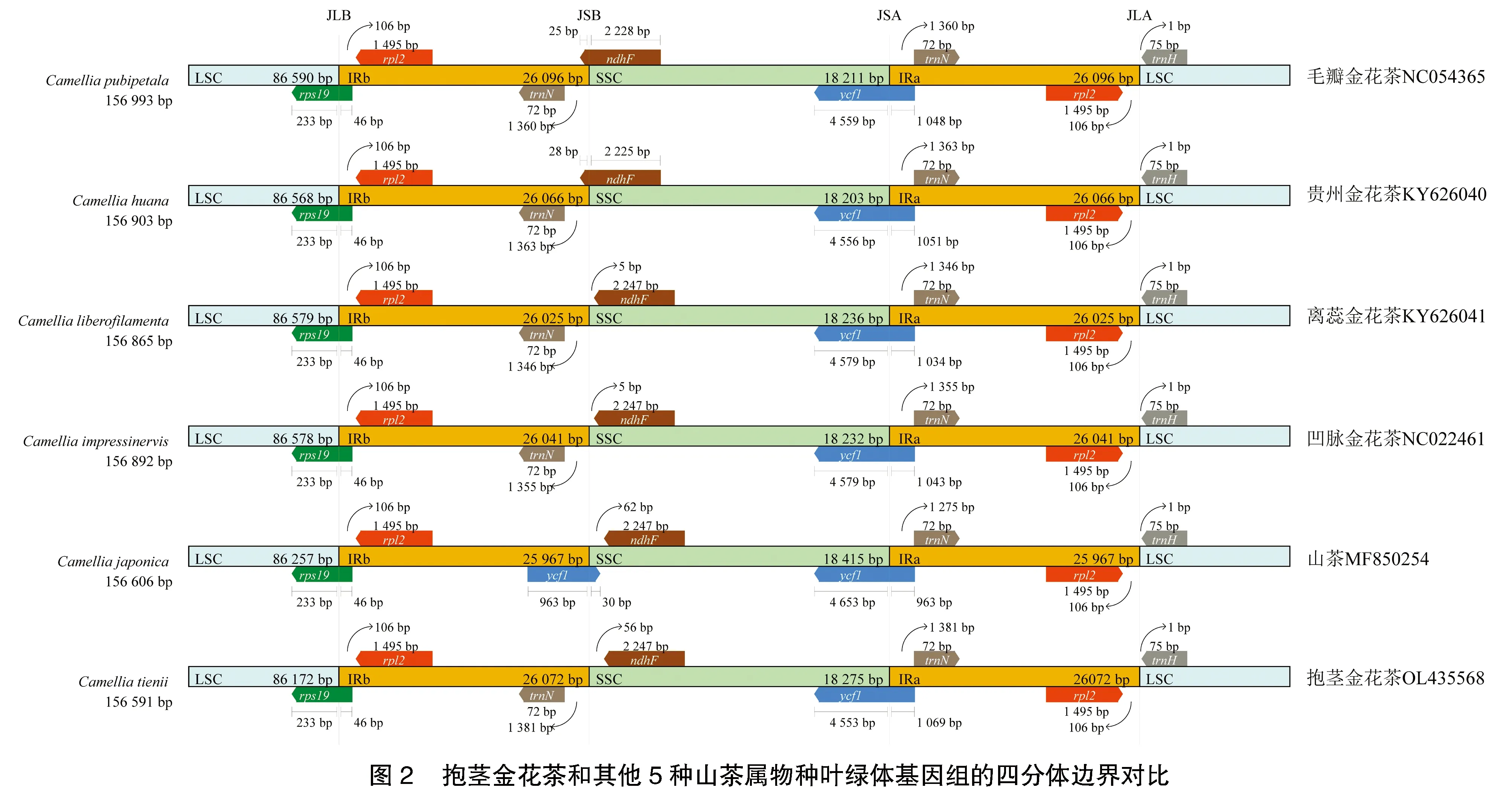
2.2 IR 边界的扩张与收缩
通过比较6种山茶属植物(山茶、毛瓣金花茶、贵州金花茶、离蕊金花茶、凹脉金花茶、抱茎金花茶)叶绿体基因组 IR 区与 LSC 和 SSC 区的边界,发现山茶属植物的叶绿体基因组较为保守,但总体上金花茶植物与山茶的叶绿体基因组边界存在一定差异(图2)。叶绿体基因组最长的为毛瓣金花茶,为156 993 bp;最短的为抱茎金花茶,为156 591 bp。其基因组长度的差异主要发生在 LSC区,相比于山茶,5种金花茶植物的IR区更长而SSC区更短,表明金花茶植物在进化过程中叶绿体基因组IR区可能有一定扩张。6个山茶属植物的rps19基因均跨越 LSC 和 IRb 边界;毛瓣金花茶和贵州金花茶的ndhF基因跨越 IRb 和 SSC,而其他4种植物的ndhF基因完全位于SSC;ycf1基因均跨越 SSC 和 IRa;trnH基因均位于位于LSC。
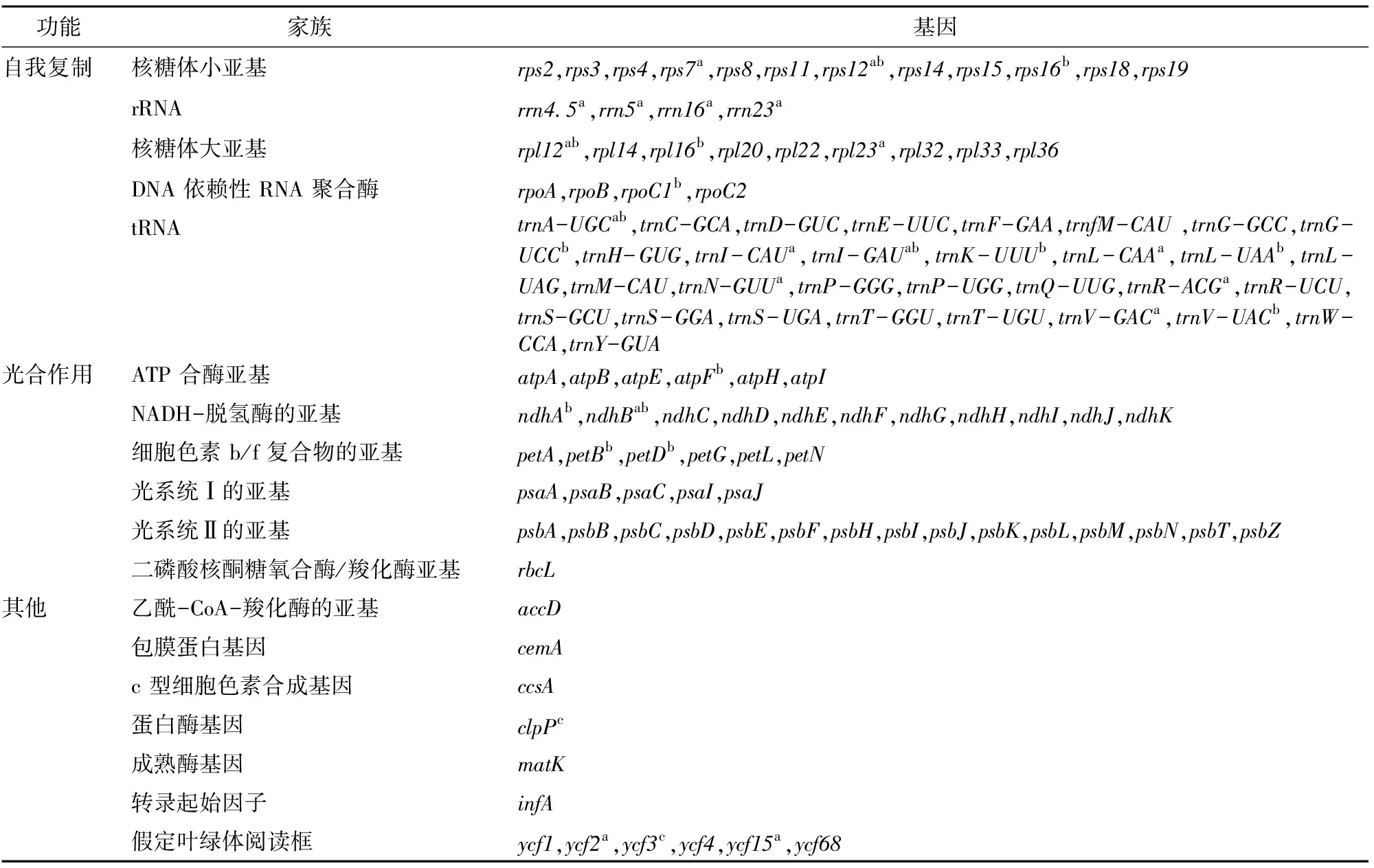
表1 抱茎金花茶叶绿体基因组注释基因
2.3 重复序列和SSR分析
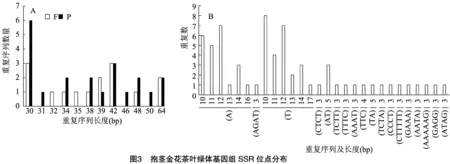
重复序列分析结果如图 3所示,抱茎金花茶叶绿体基因组含36条重复序列,其中有21条回文重复(P)和15条正向重复(F),重复序列长度范围为30~64 bp。在抱茎金花茶叶绿体全基因组中检测到67个SSRs位点,包含6种碱基重复类型,其中单碱基重复48个、二碱基重复4个、三碱基重复1个、四碱基重复11个、六碱基重复2个,混合型SSR 1个。67个SSR中有40个位于基因间区,10个位于内含子,17个位于外显子。
2.4 金花茶植物叶绿体基因组比较
以毛瓣金花茶(Camelliapubipetala)叶绿体基因组序列为参考,对9种金花茶植物的叶绿体基因组序列进行了可视化比对分析(图4)。金花茶植物叶绿体基因组序列高度相似,表明其进化十分保守。但金花茶植物叶绿体基因组序列依然存在一些差异位点,且差异性表现为非编码区>编码区、LSC>SSC>IR。编码区存在较高差异的序列为rps16和ycf3基因,非编码区存在较高差异的序列为ycf4-cemA、ycf15-trnL-CAA和rrn5-trnR-ACG,这些高变异序列经后续深入研究有望成为金花茶植物可靠的分子标记。随后选取毛瓣金花茶和抱茎金花茶的ycf15-trnL-CAA序列进行比对(图5),这段序列长度为370 bp,在215 bp和306~309 bp的位置存在差异,可以有效区别毛瓣金花茶和抱茎金花茶,碱基差异可能是由于不同的生存环境造成。
2.5 系统进化分析
为了解析金花茶植物的系统发育关系,以大头茶和长果大头茶为外类群,利用19个金花茶物种的全叶绿体基因组构建系统发育树。结果(图 6)表明,金花茶(NC024661)、小果金花茶、龙州金花茶、金花茶(NC039645)、显脉金花茶组成的分支和抱茎金花茶的分支互为姊妹分支,具有较近的亲缘关系,且自展支持率为68%。小黄花茶在金花茶组植物中形成一个基部分支,然后是抱茎金花茶。根据聚类结果,可以将所选金花茶植物归为6个大类。第1类为小黄花茶;第2类为抱茎金花茶、显脉金花茶这一分支;第3类为簇蕊金花茶;第4类为薄叶金花茶、小花进化茶、富宁金花茶、离蕊金花茶;第5类为凹脉金花茶、东兴金花茶;第6类为顶生金花茶、毛瓣金花茶、贵州金花茶、崇左金花茶、四季金花茶。每一类的金花茶植物彼此间有较近的亲缘关系。
3 讨论与结论
本研究采用双末端测序策略对抱茎金花茶进行了基因组重测序,组装并注释了抱茎金花茶的全叶绿体基因组,这为金花茶植物的物种鉴定和亲缘关系等研究提供了数据支持。抱茎金花茶全叶绿体基因组大小、结构、基因组成和顺序、CG含量等均与山茶属其他已报道的植物(山茶、浙江红花油茶、凹脉金花茶等)相似。分析发现,抱茎金花茶叶绿体基因组ndhD具有非标准起始密码子ACG。一些山茶属植物atpB和atpE有3 bp的重叠区域,psbC和psbD有52 bp的重叠区域[10],而在抱茎金花茶中atpB和atpE的重叠区域为4 bp,psbC和psbD的重叠区域为52 bp。此外,在抱茎金花茶中trnP-GGG和trnP-UGG重叠,这与油茶结论[14]一致。值得注意的是,大多数山茶属植物叶绿体全基因组GC含量均为37.3%,基因组的 GC 含量是判断物种间亲缘关系的重要指标[15];在被子植物的很多类群中infA和rps16常从叶绿体基因组中丢失[16],而在山茶属叶绿体基因组中这些基因依然保留;相比于禾本科(132 bp)和豆科(287 bp)[10],抱茎金花茶的重复序列长度较短(30~64 bp)且数量相对较少,种种迹象均表明,山茶属植物叶绿体基因组的进化可能是缓慢而保守的。
简单重复序列(SSR)是指分布在整个基因组中 1~6 bp 的重复序列,是一种高效分子标记[7,17]。在抱茎金花茶 cpDNA 中发现 67 个 SSR位点,出现最多的 SSR为单碱基重复,全部由 A/T 组成,而在二至六碱基重复中也有明显的A/T碱基偏好性。在抱茎金花茶中检测到的SSR包含6种碱基重复类型,而在红花油茶中仅鉴定出了A/T 单碱基重复类型[4],表明同为山茶属的抱茎金花茶和红花油茶的叶绿体基因组序列存在较大差异。近年来,基于叶绿体基因组筛选DNA 条形码进行物种鉴定的技术日趋成熟[18-19],陈莹等在进行金花茶组种间鉴别研究中发现rpl16系列和psbA-tmH序列能准确鉴别毛瓣金花茶[20],本研究鉴定了rps16、ycf3、ycf4-cemA、ycf15-trnL-CAA和rrn5-trnR-ACG等高变异序列,这些序列的揭示能够为金花茶植物DNA 条形码的开发提供大量信息位点。

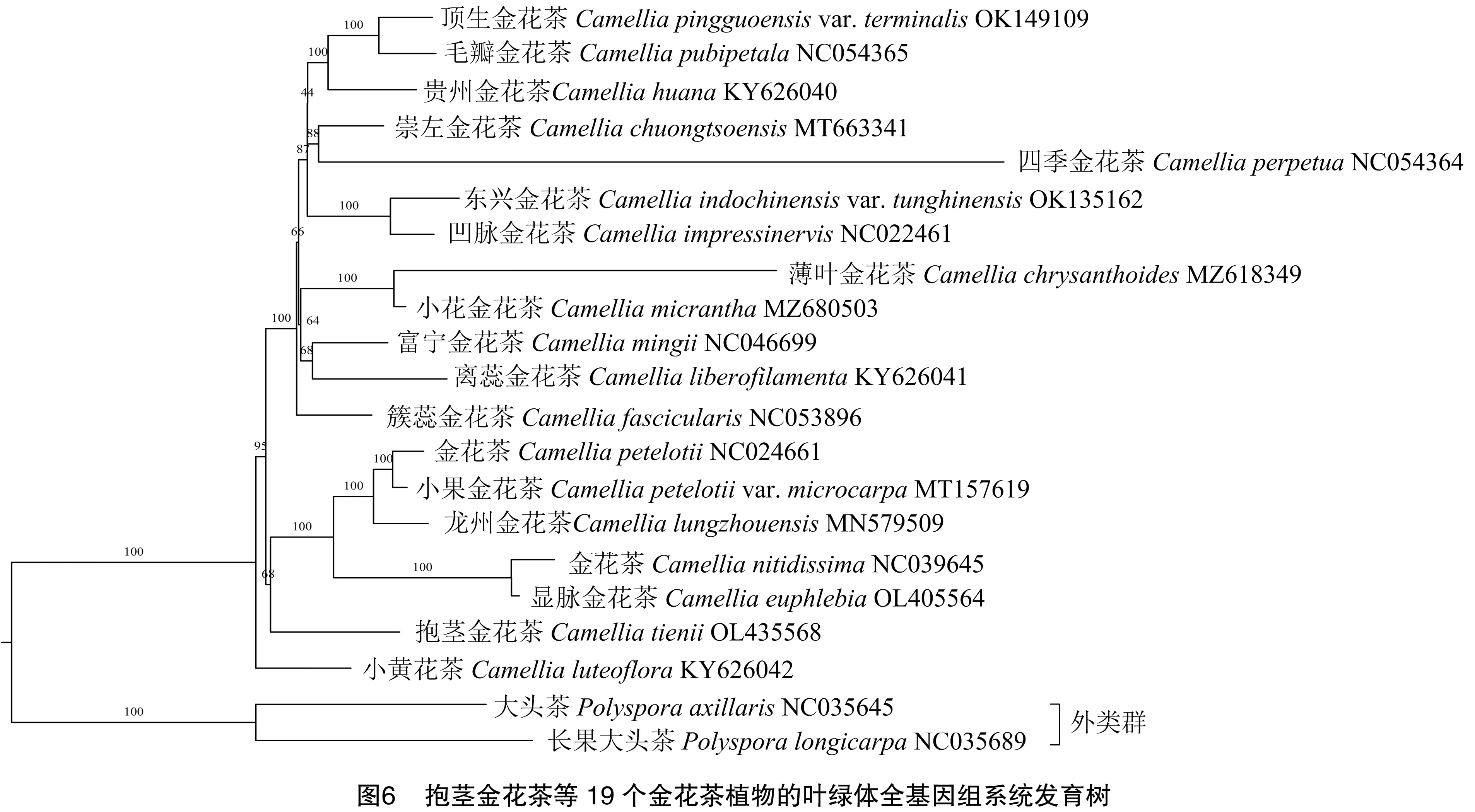
金花茶首次在我国广西发现,其金黄色、蜡质光泽的花瓣填补了山茶家族没有金黄色花的空白。研究者经过30多年的努力,明确了最适作为培育黄色系山茶花品种的杂交父母本分别为金花茶植物和山茶植物[2]。由于金花茶植物表型、生境相似,难以分辨,大量杂交种存在鉴定困难、亲本不明、亲缘关系紊乱等问题,严重滞缓了黄色山茶花新品种的培育进程。因此,明确金花茶植物间的亲缘关系对于黄色山茶花的培育具有重要价值。肖丽梅通过对金花茶植物花粉形态聚类分析认为,毛籽金花茶与东兴金花茶的亲缘关系较近[21];覃冬梅通过对金花茶叶片形态进行聚类分析认为,柠檬黄金花茶和四季金花茶亲缘关系较近[22];刘付永清利用叶绿体小单拷贝序列对金花茶植物的系统发育关系进行研究,认为地理分布相近的金花茶植物亲缘关系也较近[23]。本研究结果表明,抱茎金花茶和金花茶(NC024661)、小果金花茶、龙州金花茶、金花茶(NC039645)、显脉金花茶等亲缘关系较近,为杂交育种培育黄色山茶花奠定了相关理论基础。
