黄瓜果实冷害的发生机制与控制技术研究进展
2023-01-10卢晓婷殷菲胧刘纯友刘云芬
卢晓婷, 殷菲胧, 刘纯友, 刘云芬, 帅 良
(1.贺州学院食品与生物工程学院/广西康养食品科学与技术重点实验室,广西贺州 542899;2.广西科技大学生物与化学工程学院,广西柳州 545000)
黄瓜(CucumissativusL.)别称青瓜、胡瓜,是葫芦科(Cucurbitaceae)一年生的蔓生或攀缘草本植物,雌雄同株[1]。黄瓜果实口感清脆、味甜多汁,有独特的芳香气味,富含水分和维生素C,食用价值高,深受消费者喜爱。在低温条件下,植物组织因无法进行正常的代谢过程,会发生各种生理生化变化。黄瓜贮藏在低温条件下冷害症状不明显,在转移至室温下放置一段时间后,才会出现各种冷害症状:果实表皮组织呈水渍状、变软、褪绿黄化,并从皮孔开始出现明显的点状腐蚀,易被病原菌感染引起腐烂变质[2]。因此,采取有效的措施防止、减轻黄瓜冷害的发生,对延长黄瓜的贮藏期和货架期十分必要。
1 黄瓜冷害发生的生理生化机制
果蔬对低温胁迫的反应存在多样性,最初反应包括:膜脂相变、蛋白质或酶功能障碍等现象;继而带来一系列生理生化反应变化,如原生质流动停止、溶质渗漏增加,呼吸速率增加、乙烯生物合成的改变,及氧化磷酸化解偶联、次级代谢产物含量变化等[3]。这些改变最终会引起一系列的冷害症状。现有多种不同的生理生化机制可阐释冷害症状的发生,包括膜脂相变学说、活性氧代谢失调学说、能量代谢亏损学说、细胞壁结构物质代谢异常学说[4-6]。通过讨论黄瓜冷害发生的根本原因,可以探究有效的冷害控制方法。
1.1 低温对膜脂代谢的影响
一般来说,植物受到低温胁迫时,细胞膜会最先感知寒冷继而引起相变,从液晶态变成凝胶态,使细胞内原生质变为固态、难以流动。植物细胞在短期低温胁迫范围内通过调节其生理代谢来适应低温,否则由于膜脂代谢失衡而发生冷害,这是目前普遍接受的产生冷害的根本原因[7]。主要表现为细胞膜结构损伤、膜结合酶能力下降,膜选择透过性增加和破坏、胞内电解质大量渗漏[4]。
低温胁迫直接导致植物细胞内电解质大量外渗,故通常用电解质外渗率来表征果蔬细胞膜透性的破坏情况。此外,细胞膜损伤主要是由膜脂过氧化引起的,可通过多种反应了解其氧化的程度。丙二醛(malonaldehyde,MDA)水平的上升和膜脂肪酸成分变化就是膜结构和功能改变带来的现象[8]。MDA是膜多不饱和脂肪酸快速过氧化作用产物,低温环境下果蔬中MDA含量的增加与冷害症状的出现基本一致[4],故其含量可以评价黄瓜果实细胞膜系统氧化损伤的程度。果蔬中不饱和脂肪酸有利于维持细胞膜在低温下的稳定性,降低膜的相变温度,保护细胞结构,减轻冷害损伤。故膜脂肪酸含量和不饱和度的高低可以作为衡量膜脂过氧化程度的重要指标,若氧化严重会出现脂肪酸饱和度升高、磷脂和半乳糖值降低等情况,而且不饱和度越高,膜的流动性越强。
磷脂是生物膜的主要成分,而低温贮藏引发冷害会造成磷脂降解。磷脂酶D(phospholipase D,PLD)可将磷脂水解成磷脂酸(phosphatidic acid,PA),引起膜负弯曲和功能障碍,磷脂酶C(phospholipase C,PLC)和磷脂酶A(phospholipase A,PLA)在低温下也能催化甘油磷脂的降解,造成不可逆的膜损伤,脂氧合酶(lipoxygenase,LOX)可增强植物细胞膜的流动性和脂质的不饱和性[9-11],所以通过磷脂酶和脂氧合酶的活性变化可以判断膜脂代谢情况。
1.2 低温对活性氧代谢的影响
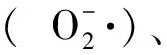
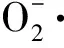
1.3 低温对能量代谢的影响
先前研究已表明,细胞膜损伤与细胞能量损失有关[17]。在低温胁迫下,植物为维持自身生命活动需要,能量损耗增加导致线粒体活性增强、ROS积累,从而影响线粒体上的酶和电子传递链,使膜透孔道被打开,导致线粒体功能异常、能量利用效率下降。此外,氧化磷酸化的解偶联会降低细胞合成的三磷酸腺苷(ATP)水平[3]。因此,认为减轻低温胁迫影响的机制可能与高水平ATP和能荷有关。
能荷水平的变化直接取决于能量代谢。线粒体H+-ATPase、Ca2+-ATPase、琥铂酸脱氢酶(succinate dehydrogenase,SDH)和线粒体细胞色素C氧化酶(cytochrome C oxidase,CCO)是能量代谢过程中的关键酶,这些酶的变化能反映线粒体的功能及合成能量的变化,其活性可被作为细胞能量代谢的指标[18]。研究表明,果蔬中这4种酶的活性会随着低温贮藏时间的延长逐渐降低,果蔬的冷害程度也随之加深。姜玉等研究发现,外源水杨酸处理能通过提高H+-ATPase、Ca2+-ATPase、SDH和CCO的活性,维持细胞内的能量代谢平衡,从而使黄瓜果实内的ATP含量和能荷保持在较高水平,延缓冷害的发生[19]。结果表明,通过增强能量代谢相关酶活性、提高能量状态和不饱和/饱和脂肪酸比值,促进能量代谢正常进行,进而可减轻黄瓜果实采后发生冷害。
1.4 低温对细胞壁结构物质代谢的影响
现有研究结果普遍认为,果实的质地主要取决于果实中果胶、纤维素、半纤维素等细胞壁物质的含量。低温胁迫同样影响果实中与果胶降解有关的酶活性,一方面抑制了β-半乳糖苷酶和纤维素酶活性上升,造成其果胶质的降解受阻[20];另一方面,果胶甲酯酶活性增强、多聚半乳糖醛酸酶活性减弱,两者变化不协同,导致细胞壁中的果胶代谢异常、果实不能正常软化。通过显微观察发现,果蔬在采后低温贮藏过程中发生冷害,诱导可溶性果胶增溶、使果实过分绵软[6],果实表面凹陷症状的发生实际上是由细胞死亡和细胞塌陷引起的。李佳乐等用梯度热水处理有效减轻了贮后黄瓜冷害,并证明抑制黄瓜果胶酶、纤维素酶及β-葡萄糖苷酶等细胞壁水解酶活性的上升,可延缓原果胶的水解及纤维素的分解,保持果实较高的硬度[21]。
1.5 低温对次级代谢产物的影响
腐胺(putrescine,Put)、亚精胺(spermidine,Spd)和精胺(spermine,Spm)等多胺类(Polyamines,PAs)是一类与植物激素类似的胺类化合物,具有脂肪族氮结构,在高等植物的分生组织和生长旺盛的组织中的浓度较衰老组织中高,在植物受到非生物胁迫时也会积累[22]。研究表明,低温诱导植物组织中多胺含量增加,而高水平的多胺可有效缓解果蔬的冷害症状并延长贮藏期[23]。在低温条件下,黄瓜果实在冷害症状出现之前,呈现内源腐胺和亚精胺开始大量增加、精胺含量较低且无明显变化规律[24]。研究证明,多胺处理会提高植物对低温的耐受性,施用亚精胺提高了黄瓜果实抗氧化酶活性和耐冷性,腐胺则通过抑制1-氨基环丙烷-1-羧酸(ACC)合酶的合成和活性减缓了乙烯的生物合成[25-26]。因此,多胺可作为激发应激保护反应的激发子,补偿低温应激的负面影响。
γ-氨基丁酸(gamma-aminobutyric acid,GABA)是一种非蛋白氨基酸,是植物体内重要的内源信号分子,对植物的胁迫响应、生长发育具有重要调节作用[27]。在植物中,细胞内GABA水平通常较低,但它们在干旱、盐分和低温胁迫下可迅速大量积累,并参与抵御这些胁迫,在黄瓜采后冷害中也有针对性研究[28,11]。其代谢途径由3种酶组成:谷氨酸脱羧酶(glutamate decarboxylase,GAD)、γ-氨基丁酸转氨酶(γ-aminobutyrate aminotransferase,GABA-t)和琥珀酸半醛脱氢酶,其中,GAD是形成GABA的关键酶。在冷胁迫下,胞内Ca2+形式的复合物与钙调素(CaM)的水平升高,促进GAD催化谷氨酸形成GABA[28]。通过多胺分解途径,GABA可以进行代谢转化。Spd、Spm经多胺氧化酶(polyamine oxidase,PAO)催化成Put、Put再经二胺氧化酶(diamine oxidase,DAO)催化生成代谢产物GABA;GABA还可以通过GABA-t转化为琥珀酸半醛,然后转化为琥珀酸参与三羧酸循环反应[29]。
脯氨酸是一种具有特殊刚性构象的氨基酸,常常参与调节细胞渗透平衡,维持细胞膜、蛋白质结构的稳定,还能作为活性氧清除剂和脂质过氧化抑制剂[30]。有证据表明,脯氨酸含量与果蔬抗冷性呈正相关,说明脯氨酸积累可能是高等植物抵抗低温胁迫的另一适应机制[31]。在高等植物中,脯氨酸一般通过Δ1-吡咯啉-5-羧酸合成酶(Δ1-pyrroline-5-carboxylate synthetase,P5CS)由谷氨酸合成,也可通过鸟氨酸-δ-转氨酶(ornithineδ-aminotransferase,OAT)由鸟氨酸替代合成,增强P5CS和OAT活性可以提高脯氨酸的生物合成能力[30]。脯氨酸脱氢酶(proline dehydrogenase,PDH)是催化脯氨酸发生降解的酶,抑制PDH活性有利于降低脯氨酸的分解。这3种酶对植物体内脯氨酸合成与降解形成动态的调节与控制,它们活性的改变是引起果实中脯氨酸含量变化的原因。
综上所述,研究采后处理对低温胁迫下植物体内多胺、γ-氨基丁酸和脯氨酸代谢及相关酶活性的影响,可为3种物质积累的生理意义提供更有价值的信息。
2 黄瓜冷害的分子机制
分析和验证冷胁迫相关基因,探究黄瓜抗低温冷害机制,可为诱导黄瓜耐冷性和预防贮藏冷害奠定基础。低温可影响调控酶的RNA和DNA转录、翻译和细胞代谢[32]。目前研究中,主要通过转录组筛选鉴定低温胁迫下差异表达的冷害相关酶、基因和转录因子,探究减轻采后冷害的潜在机制,确定介导植物适应低温的特异性分子,为植物抗冷性的产生提供了新的线索。
2.1 黄瓜耐低温相关基因及功能研究
Kong等发现,低温胁迫下出现冷害症状,通常伴随着细胞膜结构损伤、膜脂质过氧化及编码PLD、LOX的基因CaPLD和CaLOX活性和表达的增加[7];同时,低温诱导了CaPLDα4转录本的强积累,且受CaNAC1转录因子正向调控,而CaPLDα4在植物的非生物胁迫中起着重要作用[33]。更进一步来讲,褪黑素能通过上调CaGAD、CaP5CS、CaOAT和下调CaPDH基因的转录水平,触发黄瓜的脯氨酸代谢酶系统调节膜渗透,增强抗冷性[31]。类似地,随着贮藏时间的延长,ATPase、NAD和VPP的酶活性和基因表达均呈下降趋势,但PLD活性和基因表达明显提高[34],这与上述细胞膜损伤和能量缺乏诱导冷害发生的机制相呼应,说明这些与膜脂代谢、活性氧代谢、能量代谢相关酶的活性和基因表达水平的变化可导致细胞膜脂过氧化和能量损失,从而使果实发生冷害。
黄瓜中有多个与低温相关的转录因子家族,这些成员通过调节相关防御基因的表达,参与响应冷害的耐受性。如,热激转录因子CsHSFs基因的表达与热处理和冷锻炼诱导的黄瓜果实耐冷性有关,其中,CsHSF7、CsHSF11响应低温胁迫、表达量显著升高[35]。乙烯响应因子(ethylene-responsive factors,ERFs)是一类植物特有的转录因子,在响应生物及非生物逆境中发挥重要作用。通过分析ERF转录因子在黄瓜冷害及减轻冷害的进程中的表达模式,发现29个ERF基因可能与黄瓜果实冷害相关,且认为CsERF基因和黄瓜组蛋白去乙酰化酶基因CsHDAs的表达水平与黄瓜果实冷害有一定关系[36];更进一步的,CsERF17能够识别并结合膜脂合成代谢相关基因的启动子并激活它们的活性,说明ERF转录因子可能通过调控膜脂合成代谢相关基因的转录,进而调控低温胁迫下黄瓜的膜脂合成代谢,减少冷害发生[37]。WRKY转录因子参与植物免疫反应、发育和衰老的调控,已在黄瓜中发现61个WRKY家族基因[38]。进一步研究发现,过表达黄瓜细胞核中的CsWRKY46基因使转基因植株具有抗寒性,并以脱落酸(abscisic acid,ABA)依赖途径正向调控冷信号通路[39]。CBF转录激活因子CBF1、CBF2和CBF3(DREB1b、DREB1c和DREB1a)与许多低温诱导基因调控区域中的CRT/DRE元件结合,诱导其转录激活,从而控制植物对低温的反应,如过表达G蛋白Gγ亚基同源基因CsGG3.2使黄瓜对低温胁迫表现出耐受性,缓解低温损伤,同时增加CBF转录因子的表达及对其的调控[40-41]。
miRNA(microRNA)是一类来自真核生物、长度为18~24 nt的内源性非编码小分子RNA,在转录后的基因调控网络中起着关键作用。miRNA介导的基因反应是植物响应非生物胁迫的又一潜在机制。在逆境环境下,miRNA通过与目标基因mRNA碱基互补配对,对目标mRNA进行剪切或翻译抑制,或是调节RNA诱导沉默复合物(RNA-induced silencing complex,RISC)来降解目标基因的mRNA,使果蔬对各种环境胁迫做出反应[42]。近年来,miRNA在植物逆境适应过程中的调控作用已经有越来越多的报道。低温会影响甘薯抗逆应答miRNA的表达,这些miRNA是通过靶向一个蛋白质编码基因来调节甘薯对温度胁迫的应激反应[43],虽然大部分miRNA与靶基因的表达呈负相关水平,但也有可能存在正向调控机制。不同表达水平的miRNA关键基因参与不同途径的调节,miR319通过活性氧代谢参与番茄对低温胁迫的耐受调节[44]、miR528通过靶向多酚氧化酶(polyphenol oxidase,PPO)在低温下引起香蕉果皮褐变[45]。
2.2 黄瓜耐低温相关蛋白及功能研究
目前,研究中发现多个蛋白与黄瓜耐冷性密切相关。冷驯化(pre-storage cold accuration,PsCA)可以减轻黄瓜果实的冷害,上调DNA损伤诱导蛋白1(CsDDl1),提示冷驯化诱导的黄瓜耐冷性涉及CsDDl1的转录[46]。植物富含甘氨酸RNA结合蛋白(GR-RBP)在响应非生物胁迫方面的作用在黄瓜果实中也有体现,如PsCA显著上调了低温下黄瓜果实中CsGR-RBP3的转录水平,而过表达CsGR-RBP3显著上调了9个参与抗低温等防御反应的拟南芥基因,说明CsGR-RBP3在诱导黄瓜抗冷性中发挥了积极的作用[47]。通过蛋白组分析发现,贮前冷驯化使低温贮藏黄瓜中12个蛋白出现差异积累,而且大多数上调的蛋白质的积累与转录本水平的上升趋势高度一致[48]。
目前,关于黄瓜耐低温蛋白的研究鲜有报道,对于相关蛋白如何受低温影响、在转录水平的变化以及它们参与低温响应的分子机制仍需进一步研究。因此,探究黄瓜耐低温响应途径及调控因子的研究对于黄瓜耐低温胁迫反应的分子机制研究具有非常重要的意义。
2.3 黄瓜响应低温胁迫的相关信号转导机制
植物响应低温是一个非常复杂的调控网络。ROS可以作为关键的信号分子参与下游信号传导,诱导逆境胁迫相关基因表达,使植物能更好地在逆境胁迫条件下生存。仅有的一些研究表明,植物体内存在一些可能的ROS信号受体,如拟南芥中的乙烯受体ETR1、乙烯响应因子ERF6及热激转录因子HSF可能参与对ROS信号的响应,同时,ROS还可提高多种转录因子的表达,包括WRKY、bZIP、MYB基因家族等[49-52]。此外,ROS的下游信号可能还涉及Ca2+和Ca2+相关蛋白、一氧化氮(NO)、ABA等,如ABA可通过诱导H2O2的快速产生调控NO的产生进而激活丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)级联反应,上调ABA信号途径中抗氧化酶基因表达与活性[53-54]。
脱落酸(abscisic acid,ABA)是一种广泛存在于高等植物中的内源激素,是植物响应生物与非生物胁迫的重要信号物质[55]。在低温条件下ABA积累与其生物合成的增加相关,同时表现为ABA在低温下调控许多下游基因表达,如激活黄瓜叶片中冷诱导基因CBF1和黄瓜果实中CsGR-RBP3的表达来提高耐冷性[47,55-56]。ABA合成途径中的关键酶 9-顺式-环氧类胡萝卜素双加氧酶(9-cis-epoxycarotenoid dioxygenase,NCED)、玉米黄酯环氧化酶(eaxanthin epoxidase,ZEP)、乙酸醛氧化酶(ascorbate oxidase,AAO)活性及其基因表达上调,促使ABA含量大幅上升,植物抗逆性增加[52]。ABA信号传导途径中的核心反应是由ABA受体 PYR/PYL/RCARs(pyracbactin resistance/pyracbactin resistance-like/regulatory component of abscisic acid receptor)、正调控因子蔗糖非酵解相关蛋白激酶2(sucrose non-fermented related protein kinase 2,SnRK2)、负调控因子蛋白磷酸酶2C(protein phosphatase 2C,PP2C)组成的信号调控系统在低温刺激下迅速诱导ABA积累,ABA与受体PYR/PYL/RCAR结合形成复合物,导致PP2C活性被抑制,从而激活靶向膜蛋白、离子通道和转录因子的SnRK2,促进ABA响应基因转录,从而提高植物对低温的耐受性[55]。施用外源ABA发现,ABA处理促进了抗氧化酶活性,推迟了冷害症状且抑制了衰老劣变[57],同时提高了脯氨酸含量,降低了膜脂过氧化,有效缓解了细胞膜损伤,增加了植物耐冷性[58-59]。此外,ABA还可通过调节气孔运动,以增强植物应对环境胁迫的能力[52]。
3 冷害的控制措施
运用适当的处理方法诱导黄瓜的耐冷性,以提高黄瓜抗冷性和延缓采后低温贮藏冷害发生的重要方式,包括温度调节(如热处理[60]和冷处理[61-62])、控制采后贮藏条件(如提高环境湿度[63]、改良气调贮藏[64])、紫外线照射[65]等物理方法,及施用水杨酸、茉莉酸甲酯、硝普钠、多胺等化学物质[11,66]。通过这些处理改善冷害的发生,包括维持细胞膜完整性和高能状态,改善抗氧化系统,促进细胞壁正常修饰,及诱导渗透调节物质积累[67-70]。以下讨论了冷害控制方法的有效性和可能的控制机制。
3.1 温度调节处理
阶段降温可与预冷处理相结合,将阶段降温处理的前段视为去除田间热的采后预冷环节,使果蔬既达到预冷的效果,同时诱导耐冷性。王斌等使用冷库[62]、代慧等利用泡沫箱的隔热性[71],均使贮藏黄瓜实现逐渐降温的效果,增强了黄瓜果实的抗冷性,减少了瞬间低温对细胞膜的伤害,维持了更高的抗氧化活性,减轻了冷害的发生。
冷激处理(cold shock treatment,CST)是指对采后果实做不致发生冷害和冻害的短时低温处理,以延长贮藏时间、提高果蔬贮藏品质的物理保鲜方法。冷激处理下,果蔬呼吸强度降低,有效延缓了果蔬后熟进程,提高了膜脂过氧化保护酶的活性,减轻了黄瓜贮藏中的冷害[72]。因此,黄瓜在处于冷害温度范围内运输时,可通过预先冷激的方法减轻冷害以减少损失。
热激处理通过调节抗氧化酶活性、抑制活性氧产生,增加多胺和脯氨酸含量以及诱导热激蛋白的积累,提高果蔬对冷害的耐受性。黄瓜经过合适温度热激处理,硬度增大,失质量率、LOX活性、氧自由基含量减小,这说明适宜的热激处理能够延缓膜脂过氧化发生,达到减轻冷害、延长贮藏期的作用[73-74]。采前对黄瓜果实进行高温处理,采后 10 ℃ 的贮藏条件下仍能保持较好的耐冷性[75]。乔勇进通过热处理显著提高黄瓜内源多胺的含量来减轻冷害发生[24]。
间歇升温是指将果蔬产品在低温生长或贮藏期间1次或多次移至常温一段时间来中断低温伤害。这种方法使受损的冷冻组织在暴露于温暖的温度时得以修复,同时分解冷害条件下积累的有毒物质并补充冷害中消耗的物质,提高细胞代谢活性,降低冷害发生率[23]。但需要注意温度变化速度和平均温度梯度,避免因热损伤造成软化、黄化等不良现象。
3.2 气调贮藏
气调贮藏是目前较为先进的果蔬贮藏手段,实质是在冷藏的基础上对气体成分进行调节,通过抑制果蔬采后呼吸作用来实现采后保鲜,主要分为人工气调贮藏(CAS)和自发气调包装(MAP)[23]。冷害温度下,气体成分对冷害的影响有显著性差异,其中,5% O2和1% CO2处理组效果最佳[76]。用袋密封包装的黄瓜比未包装的黄瓜遭遇的冷害要轻,可能是因为密封袋内CO2浓度上升、O2浓度下降,延缓了抗氧化酶活性降低,同时低温胁迫诱导产生的高水平多胺也有助于提高果实的耐冷性[77]。因此,可结合气调贮藏进行黄瓜果实的贮运,以减轻低温贮藏冷害的影响,提高采后黄瓜的货架期。
3.3 紫外线辐射处理
紫外线照射已在多种果蔬的采后低温贮藏中有所应用,紫外线短期处理能有效减轻冷害,能够延缓微生物的侵袭和衰老。短时间紫外线照射减缓果实软化速度,不仅能刺激生成乙烯,还使多胺的积累有增加的趋势、提高黄瓜果实的总抗氧化能力,这些反应可能均有利于提高果实组织对抗衰老和冷害能力[78-79]。
3.4 高湿环境处理
高湿贮藏能够显著抑制黄瓜果实冷害指数和失重率的升高,减少水分、叶绿素、抗坏血酸、氨基酸等物质的损失,显著增加抗氧化酶活性和抗氧化能力,减轻ROS对细胞膜的损伤作用,较好地维持了黄瓜果实在低温冷藏期间的品质,展示出良好的发展应用前景[63]。
3.5 涂膜处理
涂膜处理主要是减慢蒸腾的速度、降低呼吸强度,抑制采后果实中的水分从组织流失,防止表皮下的细胞塌陷形成凹陷,如使用食用蜡[80]提高商品外观,降低冷害程度;它的作用方式还可能是涂层具有抗氧化效果,减缓膜脂过氧化的发生、抑制细胞膜透性上升,如低温下采用1.00%壳聚糖涂膜有效地延迟了黄瓜冷害出现的时间,改善了黄瓜在低温下的贮藏品质[81]。
3.6 化学处理
水杨酸(salicylic acid,SA)是一种广泛存在于植物防御反应中重要的信号分子,参与调节植物生长发育中的许多过程[82]。前人研究发现采用水杨酸处理可减轻黄瓜冷害的发生,其诱导抗冷性的机制有多种猜测,主要认为外源SA处理能够降低果实ACC的积累,调节H2O2相关的氧化胁迫反应,诱导抗氧化酶活性,有效抑制细胞膜过氧化作用[83];也有的使黄瓜保持较高的能荷水平、提高脯氨酸和多胺含量使冷害症状减轻[19,23]。
茉莉酸甲酯(methyl jasmonate,MeJA)是植物体内的信号分子,对植物生长发育起调控作用,在植物遭受逆境胁迫时发挥着重要作用。外源MeJA溶液处理抑制了黄瓜果实冷害的发生,发现在贮藏期间膜透率和脯氨酸含量降低,CAT、P5CS、OAT和GAD活性升高,说明MeJA可能通过影响活性氧代谢、脯氨酸含量和GABA代谢来缓解冷害作用[84]。
一氧化氮(nitric oxide,NO)是植物体内参与多种生理过程的重要信号分子,参与植物叶片和根的生长发育、果实的成熟和衰老、抗逆等过程[85]。许多研究表明,内源NO能通过抑制ROS的积累来保护植物细胞、对抗氧化应激,并阻碍细胞膜的结构性损伤。Yang等对黄瓜果实进行适宜浓度的一氧化氮处理,发现NO通过改善抗氧化系统减缓了膜脂过氧化作用,进而提高了黄瓜的抗冷性[68]。外源NO处理提高耐冷性,还可能通过调节脯氨酸代谢、抑制乙烯生成来降低果蔬冷害和品质劣变[86-87]。
6-苄基氨基嘌呤(6-benzylaminopurine,6-BA)是人工合成的细胞分裂素类化合物,对人体无毒害,但应用实例较少。目前,研究表明,6-BA处理可提高SOD、CAT等抗氧化酶的活性,并维持能量供给平衡,使黄瓜的冷害症状减轻[16]。
4 总结与展望
黄瓜作为深受人们喜爱的蔬菜作物,在我国种植广泛、在市场上的需求量高,发展前景广阔。黄瓜果实采后常用低温贮藏进行保鲜,以保证其货架期寿命及商品价值。然而,冷链贮运过程中长时间的低温不良影响,会引发黄瓜果实一系列生理生化变化,使其产生冷害症状,造成重大的经济损失。因此,减缓和防止低温贮藏造成的冷害,对黄瓜采后贮藏保鲜具有十分重要的意义。
阐明果蔬冷害机制是寻求有效控制果蔬冷害新方法的理论基础。综合现有的研究成果,对黄瓜冷害产生的机制有了更深刻的认识,目前主要有膜脂相变理论、活性氧自由基伤害理论、能量亏损理论、细胞壁降解理论,以及冷害同植物激素、次级代谢产物、冷害相关信号途径之间的联系,不同的响应机制对应延缓和降低冷害症状的有效措施也各不相同。随着高新技术的发展,全转录组、代谢组学技术应运而生,这为从更深层次解析黄瓜冷害的发生机制提供了便利,如筛选出黄瓜果实对低温应激的miRNA/靶标组合,验证miRNA及其靶基因在低温下的差异表达,更深入地探究了解黄瓜果实在冷害温度胁迫下的分子调控机制。
在防止或减轻采后果蔬低温贮藏冷害措施方面,目前主要有热处理、冷激处理、气调贮藏、紫外辐射和化学处理等,但其如何减轻冷害的确切机制仍然不清楚。不同控制技术之间还可以相互结合优化,以达到最优的抗冷害效果。考虑到影响果蔬采后低温贮藏冷害的因素,除了贮藏环境等外界因素外,还要注意黄瓜本身所具有的特性与冷害发生的联系;在今后的研究中,可以选择利用基因工程的手段培育出耐低温、抗冷害的黄瓜新品种,从根本上保证各种质资源的品质。
