基于代谢组学分析补喂发酵豆粕对伊犁马驹粪便代谢物的影响
2023-01-10钟浩杰姚新奎罗鹏辉姚岳扬王川坤任万路
钟浩杰,姚新奎,2,罗鹏辉,孟 军,2,姚岳扬,王川坤,任万路
(1.新疆农业大学动物科学学院,乌鲁木齐 830052;2.新疆马繁育与运动生理重点实验室/新疆农业大学马产业研究院,乌鲁木齐 830052;3.新疆维吾尔自治区畜牧总站,乌鲁木齐 830052)
0 引 言
【研究意义】代谢组学可以系统、高效、全面的反映动物机体对复杂营养物质的代谢过程,是一种非常适合研究营养与代谢复杂关系的方法[1]。代谢组学利用先进的分析化学技术更系统、更全面的检测血液、尿液或粪便中的小分子代谢物。生物医学科学样本包括组织或组织提取物以及生物液体[2]。粪便中大量的化合物直观的展现了消化道以及不同空间的菌群对营养物质的消化吸收的过程。识别饮食改变引起的细微代谢变化,使用更综合的方法全面的分析粪便代谢物,对获得更多马匹消化、吸收和代谢的相关信息具有重大意义。【前人研究进展】发酵工艺已被广泛用于提高豆粕的营养物质的生物利用度[3]和降低抗营养因子的水平[4]。Frias等[5]也证实了发酵过程能够降解SBM的抗营养和致敏物质。Wang等[6]、Espinosa等[7]研究发现,仔猪饲粮中添加发酵豆粕对氨基酸消化率、代谢能、表观回肠消化率均无显著影响,但可以显著改善断奶后的生长性能。吴丹丹等[8]研究表明,奶牛饲粮中添加100g/d小肽能显著提高微生物蛋白产量,有效提高氮的消化利用率和沉积效率,减少氮排放,提高奶牛生长性能。Feng等[9]研究发现,相比于普通豆粕,发酵豆粕可以显著提高肉鸡肠道中胰蛋白酶、脂肪酶和蛋白酶活性,提高空肠黏膜绒毛高度,降低隐窝深度。【本研究切入点】发酵豆粕在牛、羊、猪、鸡以及水产等动物营养研究中获得了相应的研究成果,但是在马匹上的研究鲜见报道。需研究补喂发酵豆粕对伊犁马驹粪便代谢物的影响。【拟解决的关键问题】基于UPLC-MS/MS检测方法以及多元统计分析,研究补喂发酵豆粕对伊犁马驹粪便代谢物的影响,从分子水平分析发酵豆粕对伊犁马驹代谢的影响机制,为伊犁马驹专用饲料的研究与开发提供依据。
1 材料与方法
1.1 材 料
1.1.1 马 匹
试验于2021年5月至2021年10月在昌吉良源农牧业发展有限公司马场进行,选取10匹体况相近、无消化系统疾病病史的12月龄伊犁马驹,随机分为两组,每组5匹马。对照组和试验组分别在干草和普通精饲料的基础上添加5%的普通豆粕和5%的发酵豆粕。分别饲于不同圈舍,预饲7 d,试验期为56 d,整个试验过程中,所有马均为舍饲,垫料选取清洁干燥的木屑,每天分3次给料,自由饮水和舔食舔砖。
1.1.2 基础饲料
试验所用发酵豆粕由新疆酵博仕生物科技有限公司生产,菌种为植物乳酸杆菌、地衣芽孢杆菌、酿酒酵母等。试验过程中,所有试验马匹每天饲喂干草(1.5 kg/100 kg体重),每天按照每匹马驹体重的1%饲喂精饲料。表1
1.1.3 样品采集
试验56 d无菌采集试验马匹直肠粪便,所有用于非靶向代谢组学分析的样品均收集在5 mL无菌冻存管中,样品立即放入液氮中储存,直到粪便代谢产物的提取。
1.1.4 仪器与试剂
(1)质谱:四级杆-静电场轨道阱高分辨质谱仪(Thermo Fisher Scientific);
(2)色谱:超高效液相色谱系统(Dionex);
(3)离心机(Thermo Fisher Scientific);
(4)色谱柱:Waters, ACQUITY UPLC BEH Amide 1.7 μm,2.1 mm×100 mm column;
(5)试剂:乙腈、乙酸铵、甲醇等。
1.2 方 法
1.2.1 代谢物提取
称取100 mg粪便样本,将甲醇、乙腈和水按4∶4∶2的比例混匀,预冷后取400 μL加入粪便样本中,充分混匀,-20℃静置60 min,离心机调至4℃离心20 min,将上清液进行干燥处理,乙腈和水按1∶1的比例混匀,取100 μL加入上述经过干燥处理的容器中再次溶解提取到的代谢物,混匀后 4℃离心15 min,取2 μL上清液质谱分析。
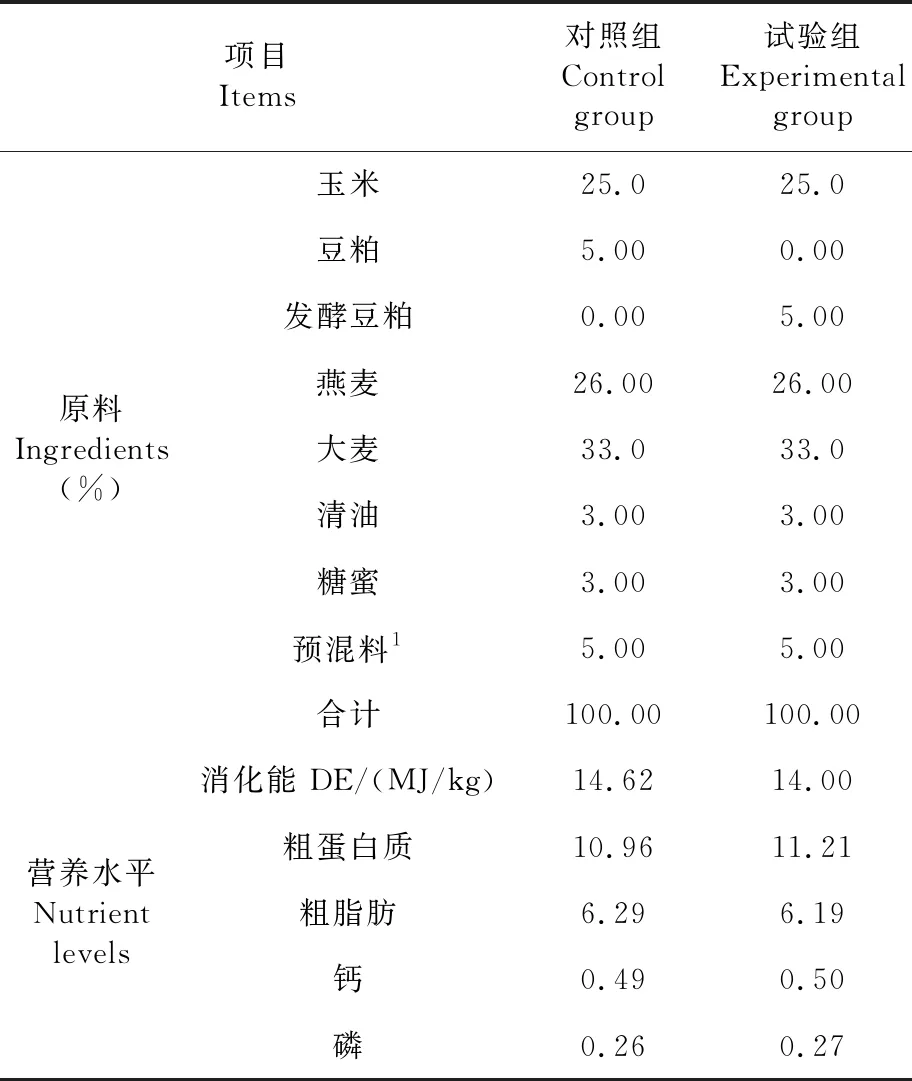
表1 试验日粮组成及营养水平
1.2.2 LC-MS/MS
(1)色谱条件
采用 ACQUITY UPLC BEH C18柱进行色谱分离,色谱柱柱温为40℃,流速为 0.3 mL/min。对代谢物进行不同梯度脱洗。个样品检测量为5 μL。分析过程中样品始终处于4℃环境中。采用随机分析。将设置好的QC样品置于各样品间依次检测。
(2) 质谱条件
各组样品将在正、负电场两种环境下检测,经UHPLC分离后用四级杆-静电场轨道阱高分辨质谱仪进行质谱分析。
1.3 数据处理
用Compound Discoverer 3.0软件对质谱条件下获得的原始数据进行峰提取、峰对齐、峰校正、标准化等处理。输出由样本名称、谱峰信息、及峰面积组成的三维数据矩阵。采用二级谱图匹配和精确质量数匹配的方式对化合物分子结构进行鉴定,与数据库比对。数据经Pareto-scaling预处理后,进行PCA和OPLS-DA分析。
2 结果与分析
2.1 QC样本总离子流图谱比较
研究表明,各色谱峰保留时间和峰响应强度均波动小,在实验过程中仪器误差引起的变异较小。图1
2.2 正、负离子模式PCA模式下数据分离趋势
说明2组样品在代谢物组成上有明显差异。图2
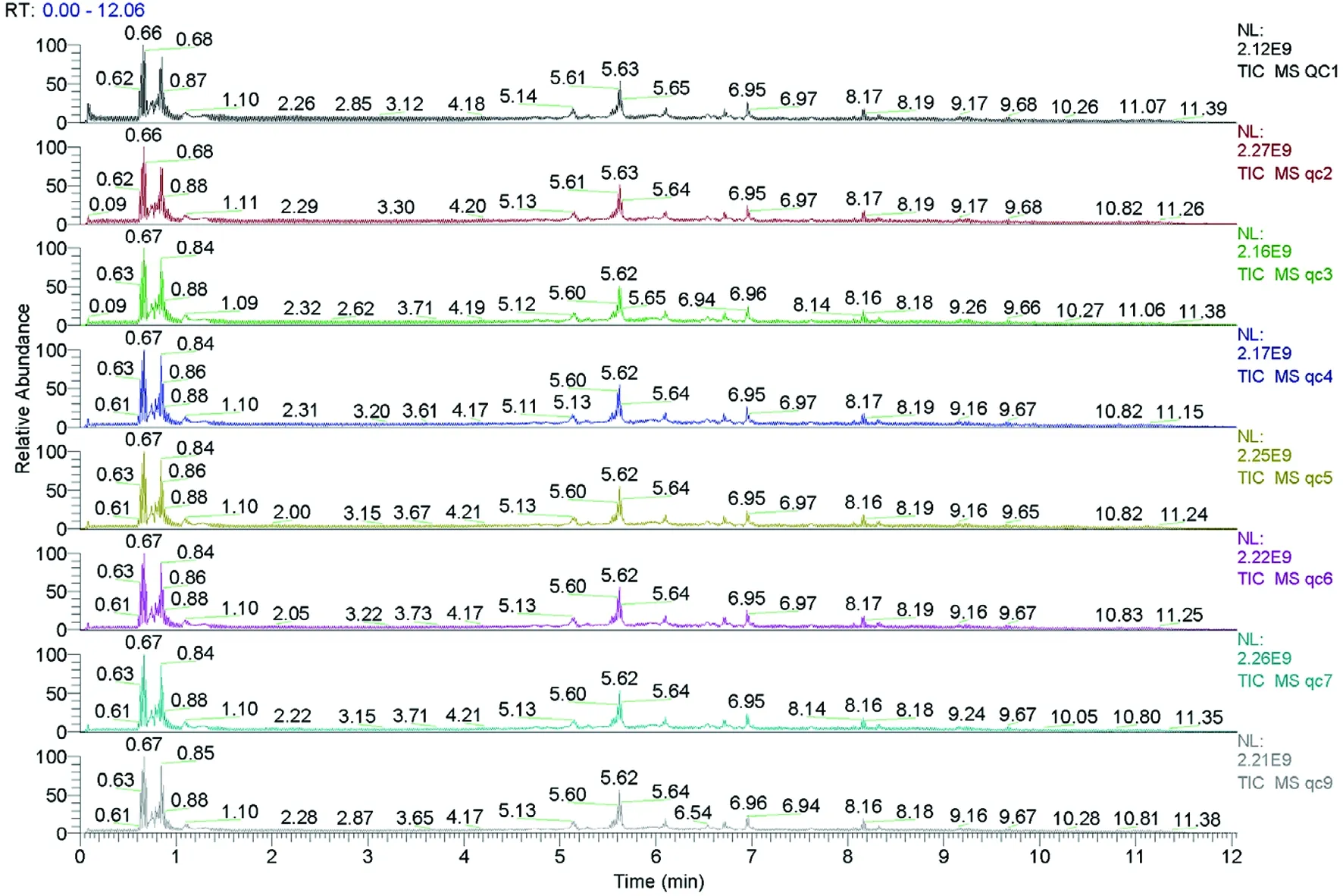
图1 QC样本TIC图谱
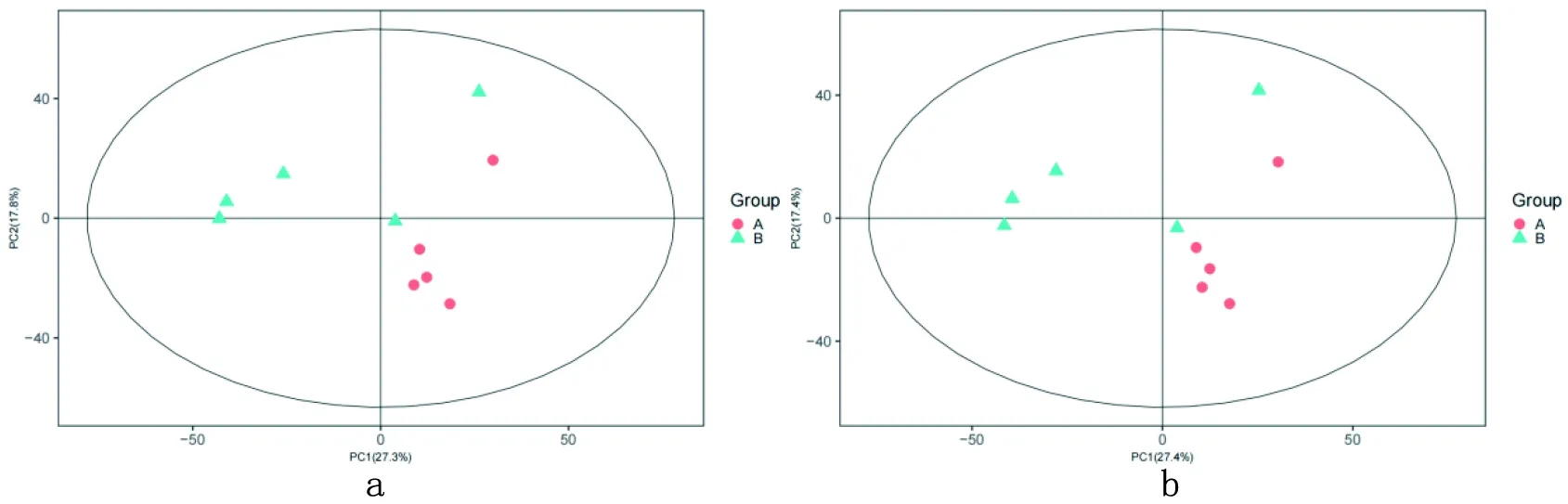
a:对照组Control group b:试验组Experimental group
2.3 POLS-DA分析
研究表明,在正、负离子模式下的POLS-DA置换检验,各组Q2均小于0.05,没有过度拟合。表2,图4
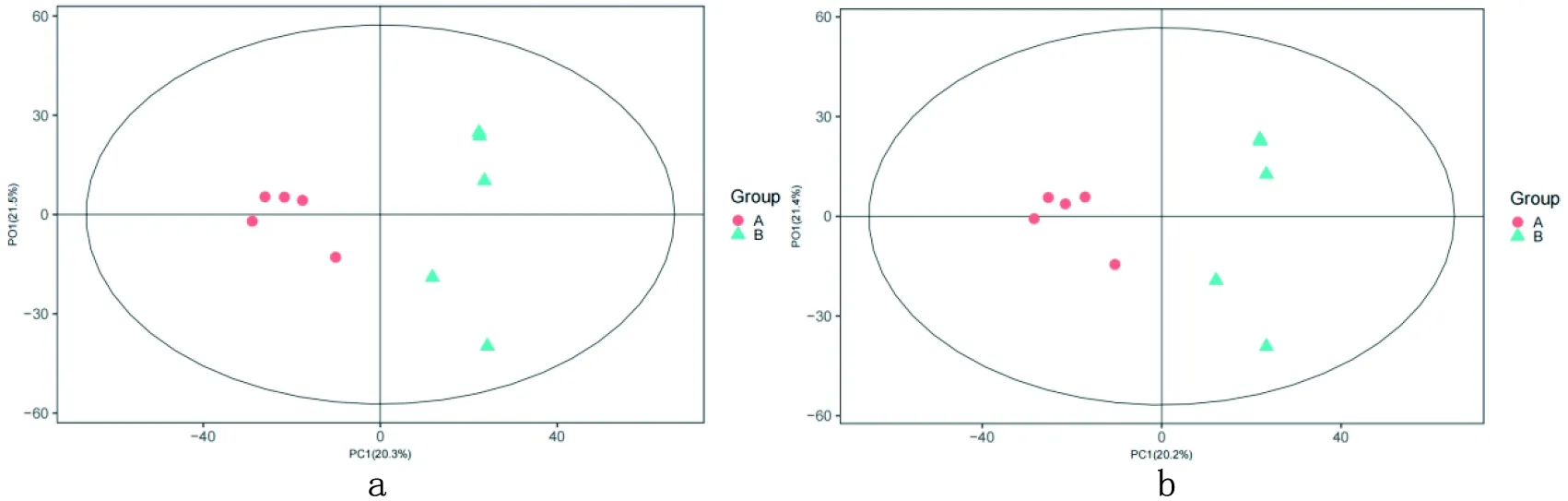
a:对照组Control group b:试验组Experimental group
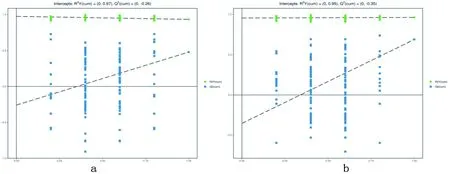
图4 正(a)、负(b)离子模式OPLS-DA得分比较

表2 OPLS-DA模型评价参数
2.4 差异代谢物筛选对比
研究表明,在试验组与对照组的对比中,共筛选出211种差异代谢物,其中130种代谢物上调,181种代谢物下调。图5
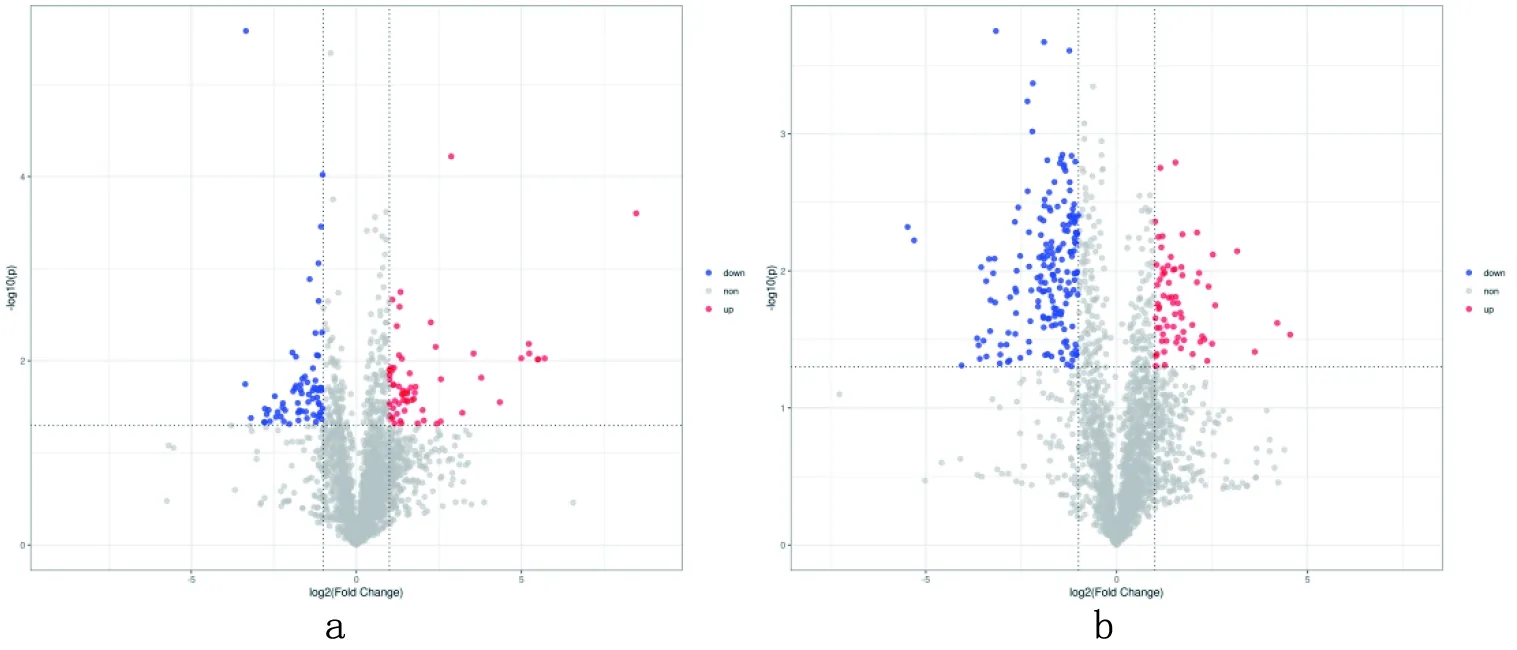
图5 正(a)、负(b)离子模式火山对比
试验组和对照组的对比中,共筛选出显著性差异代谢物9种,添加5%的发酵豆粕显著提高了马驹粪便中果糖基甘氨酸、甜菜碱、甘露醇等物质的含量(P<0.05),显著降低了高香草酸、丙酮酸、苯乙醇、烟酸和对羟苯基乙醇的含量(P<0.05)。 表3
2.5 差异代谢物聚类对比
研究表明,正、负离子模式下分别有4个和5个簇,正离子热图中的簇1、簇2在对照组中分布最多,在试验组的部分个体中少量分布;簇3和簇4在试验组中的分布明显高于对照组。负离子热图中的簇1、簇2在对照组中分布最多,而簇3、簇4、簇5集中分布在试验组。图6
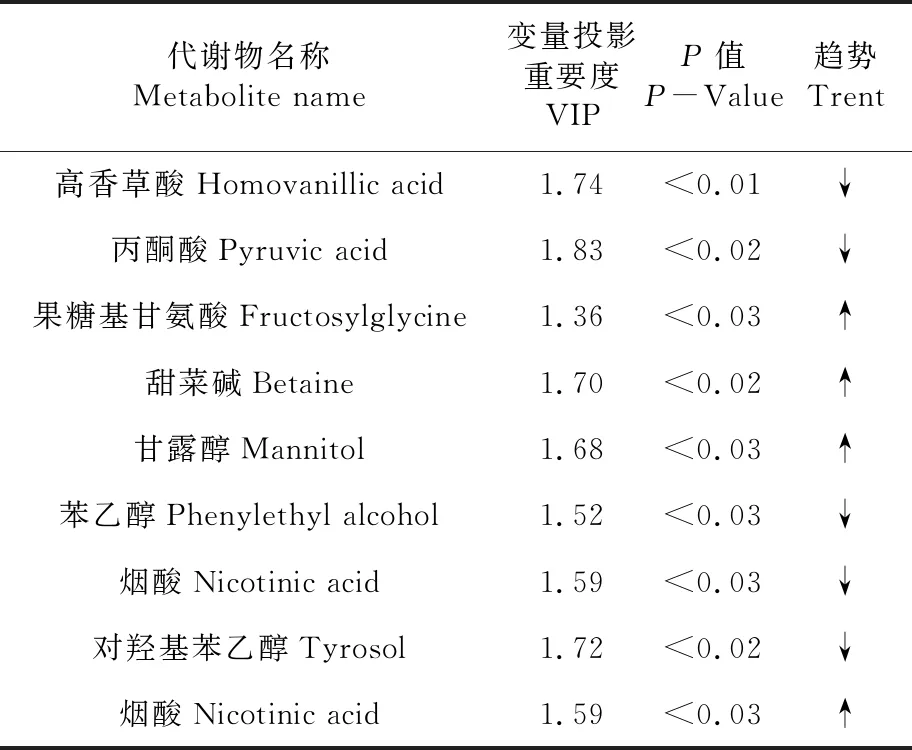
表3 试验组与对照组之间显著性差异代谢物
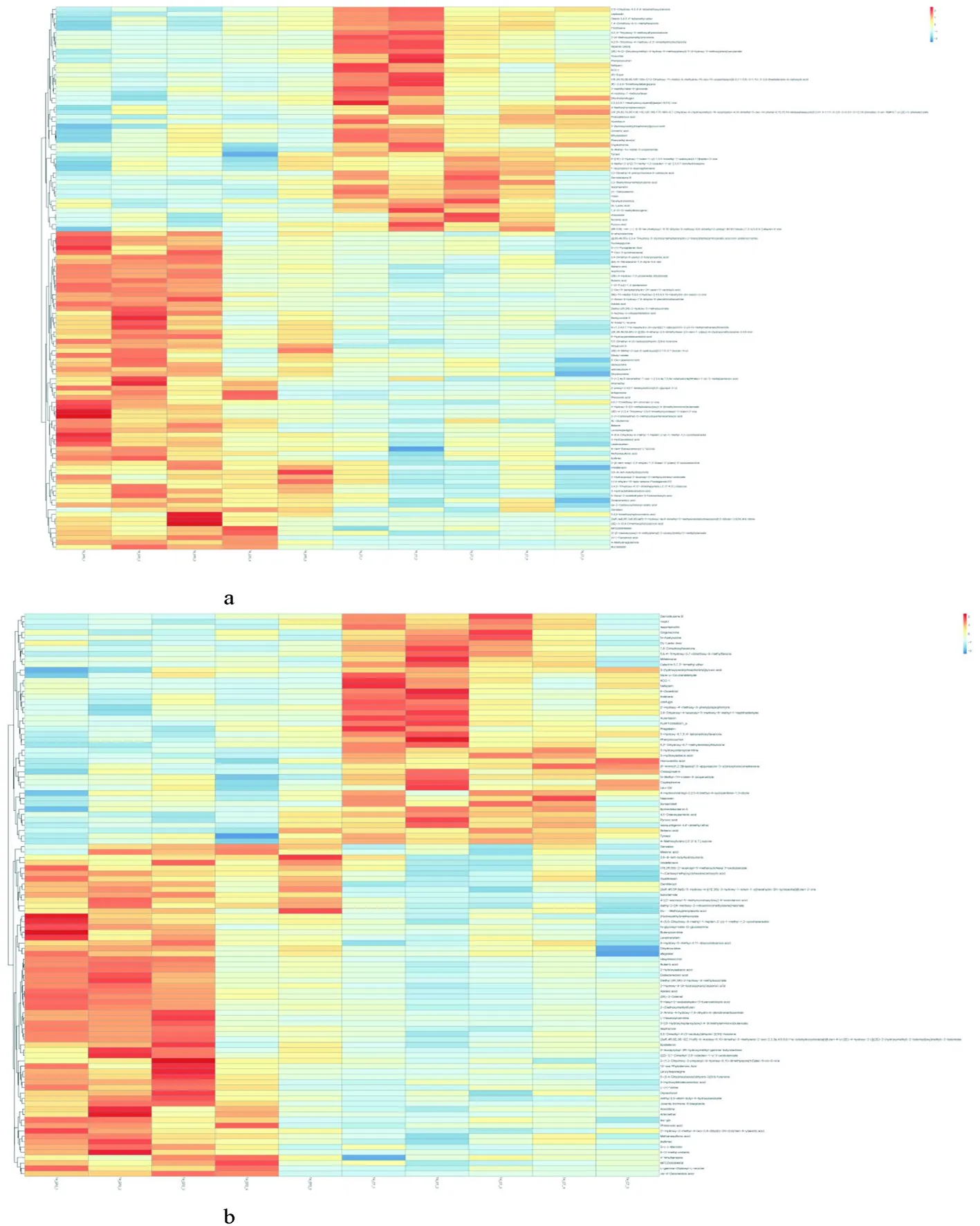
图6 正(a)、负(b)离子模式聚类热图
2.6 粪便差异代谢物KEGG通路对比
研究表明,试验组与对照组两组间差异显著的代谢通路,主要包括在甘氨酸、丝氨酸和苏氨酸代谢(Glycine, serine and threonine metabolism)、酪氨酸代谢(Tyrosine metabolism)、ABC转运蛋白(ABC transporters)、苯丙氨酸代谢(Phenylalanine metabolism)、百日咳(Pertussis)、多巴胺能突触(Dopaminergic synapse)、胰岛素分泌(Insulin secretion)、HIF-1信号通路(HIF-1 signaling pathway)烟酸和烟酰胺代谢(Nicotinate and nicotinamide metabolism)等通路。图7
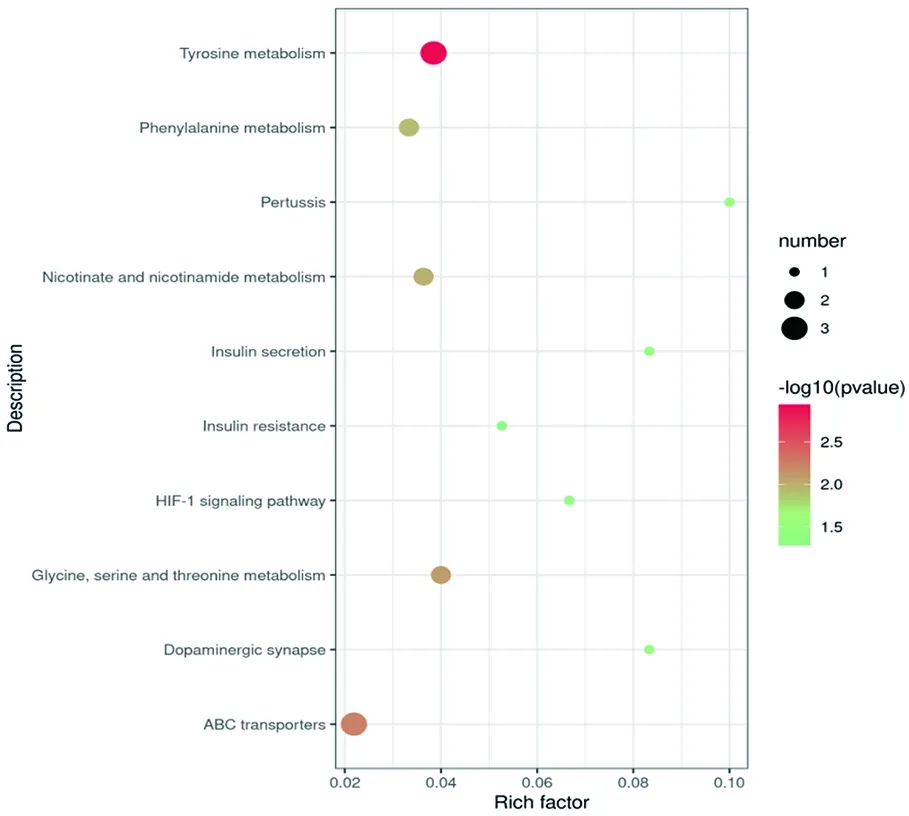
图7 试验组与对照组粪便代谢通路
3 讨 论
Escalona等[10]研究,将核磁共振代谢组学分析应用于不同种类的马生物样本,并且生成了一份“代谢物图谱”,其中包含了在普通马体液中识别出的所有代谢物。粪便和尿液代谢谱有助于解释宿主和微生物的相互作用,而血浆代谢组可以推断宿主的生理机能。自Escalona等[10]将核磁共振代谢组学分析应用于不同种类的体液样本以来,NMR代谢组学被进一步应用于马的生物体液,包括滑液[11]、尿囊液[12]和气管冲洗液[13]。
甜菜碱又称三甲基甘氨酸,甜菜碱最初在甜菜中被分离出来,该物质具有多种生理活性,目前已作为一种饲料添加剂,应用于各种畜禽养殖中。甜菜碱的渗透作用能够调节动物体内渗透压,提高血清中生长激素水平,促进动物生长;具有改善胴体成分、提高肉品质等功效[14,15]。除了作为渗透性物质的作用外,其代谢作用已被证明在保护肝脏和其他组织以及减轻心血管危险因素方面具有重要作用。甜菜碱被认为是预防慢性疾病的重要营养物质[16]。甜菜碱是动物机体内重要的甲基供体,促进脂肪分解,抑制脂肪合成[17]。贺绍君等[18]研究发现热应激状态下,在基础饲粮中添加甜菜碱可改善AA肉鸡生长性能,提高十二指肠消化酶的活性,维持盲肠内微生物区系平衡,从而减轻热应激对肉鸡的危害。研究通过添加5%的发酵豆粕显著提高了伊犁马驹粪便中的甜菜碱、甘露醇、果糖基甘氨酸等物质的含量,并参与ABC转运蛋白和甘氨酸、丝氨酸和苏氨酸代谢通路,说明饲料中添加5%的发酵豆粕能够调节伊犁马驹渗透压以及脂肪、蛋白质和氨基酸代谢,并具有改善饲料适口性、抗应激和转甲基等作用。
烟酸又称尼克酸,是动物必须维生素之一,广泛分布于动植物种[19]。烟酸作为辅酶Ⅰ(NAD)和辅酶Ⅱ(NADP)的前体,既在机体碳水化合物、蛋白质和脂肪代谢中扮演着重要的角色,又能参与体内能量代谢的调节[20,21]。具有抗炎、抗氧化、调节脂质代谢[22]、缓解应激等多种生物学功能。试验中,试验组烟酸含量显著高于对照组,通过烟酸代谢通路调节维生素代谢,饲粮中添加5%的发酵豆粕能够调节伊犁马驹碳水化合物、蛋白质和脂肪代谢,并且具有抗炎、抗氧化、缓解应激等功能。
4 结 论
饲料中添加5%的发酵豆粕显著提高了伊犁马驹粪便中甜菜碱、果糖基甘氨酸、甘露醇和烟酸等物质的含量,根据不同代谢产物在机体内发挥的生理功能可知补喂发酵豆粕能够调节伊犁马驹碳水化合物、蛋白质、脂肪和氨基酸代谢,并且具有抗氧化、抗炎、缓解应激、调节渗透压等多种生物学功能。
参考文献(References)
[1] Gibney M J, Walsh M, Brennan L, et al. Metabolomics in human nutrition: opportunities and challenges [J].AmericanJournalofClinicalNutrition, 2005, 82(3): 497-503.
[2] Patti G J, Yanes O, Siuzdak G. Innovation: Metabolomics: the apogee of the omics trilogy [J].NatureReviewsMolecularCellBiology, 2012, 13(6): 263-269.
[3] Hotz C, Gibson R S. Traditional food-processing and preparation practices to enhancing the bioavailability of micronutrients in plant-based diets [J].TheJournalofNutrition, 2007, 137:1097-1100.
[4] Egounlety M, Aworh O C. Effect of soaking, dehulling,cooking and fermentation with Rhizopus oligosporus on the oligosaccharides, trypsin inhibitor, phytic acid and tannins of soybean (Glycine max Merr.), cowpea (VignaunguiculataL.Walp) and ground bean (Macrotyloma geocarpa Harms)[J].JournalofFoodEngineering, 2003, 56:249-254.
[5] Frias J Y, Song S, Martínez-Villaluenga C, et al. Immunoreactivity and amino acid content of fermented soybean products [J].JournalofAgriculturalandFoodChemistry, 2008, 56:99-105.
[6] Wang Y, Luwq, Lidf, et al. Energy and ilea digestible amino acid concentrations for growing pigs and performance of weanling pigs fed fermented or conventional soybean meal [J].Asian-AustralasianJournalofAnimalSciences, 2014, 27(5): 706-716.
[7] Espinosa C D, Oliveira M S F, Lagos L V, et al. Nutritional value of a new source of fermented soybean meal fed to growing pigs [J].JournalofAnimalScience, 2020, 98(12): 357.
[8] 吴丹丹, 滕乐邦, 栾正庆, 等. 小肽对奶牛瘤胃微生物蛋白产量、产奶性能和氮排泄的影响[J]. 动物营养学报, 2016, 28(4): 1019-1098.
WU Dandan, TENG Lebang, LUAN Zhengqing, et al. Effects of small peptides on rumen microbial protein yield, milk performance and nitrogen excretion of dairy cows [J].JournalofAnimalNutrition, 2016, 28(4): 1019-1098.
[9] Feng J, Liu X, Xu Z R, et al. Effects of fermented soybean meal on digestive enzyme activities and intestinal morphology in broilers [J].PoultryScience, 2007, 86(6): 1149-1154.
[10] Escalona EE, Leng J, Dona A C, et al. Dominant components of the thoroughbred metabolome characterised by 1H-nuclear magnetic resonance spectroscopy: a metabolite atlas of common biofluids [J].EquineVeterinaryJournal, 2015, 47:721-730.
[11] Anderson JR, Phelan MM, Clegg PD, et al. Synovial fluid metabolites differentiate between septic and nonseptic joint pathologies[J].ProteomeResearch, 2018, 17:2735-2743.
[12] Beachler T, Gracz H, Long NM, et al. Allantoic metabolites, progesterone, and estradiol-17β remain unchanged after infection in an experimental model of equine ascending placentitis [J].JournalofEquineVeterinaryScience, 2019, 73:95-105.
[13] Bazzano M, Laghi L, Zhu C, et al. Metabolomics of tracheal wash samples and exhaled breath condensates in healthy horses and horses affected by equine asthma[J].JournalofBreathResearch, 2018, 12(3):046015.
[14] 郭雪峰, 边连全, 付亮亮. 甜菜碱在畜牧业上的应用研究[J]. 今日畜牧兽医, 2007, 2(3): 49-50.
GUO Xuefeng, BIAN Lianquan, FU Liangliang, et al. Application of betaine in animal husbandry [J].Today'sAnimalHusbandryandVeterinary, 2007, 2(3): 49-50.
[15] 岳萌萌, 卜也, 何丽华, 等. 甜菜碱的生物学功能及在肉牛生产中的应用研究[J]. 黑龙江畜牧兽医, 2021, 6(11): 44-47.
YUE Mengmeng, BU Ye, HE Lihua, et al. Biological function of betaine and its application in beef cattle production [J].HeilongjiangAnimalHusbandryandVeterinary, 2021, 6(11): 44-47.
[16] Craig S A. Betaine in human nutrition [J].TheAmericanJournalofClinicalNutrition, 2004, 3(80): 539-549.
[17] Gao X, Zhang H, Guo X F, et al. Effect of Betaine on Reducing Body Fat-A Systematic Review and Meta-Analysis of Randomized Controlled Trials[J].Nutrients, 2019, 11(10): 2480.
[18] 贺绍君, 赵书景, 李静, 等. 甜菜碱对热应激肉鸡生长性能、十二指肠消化酶活性及盲肠微生物区系的影响[J]. 动物营养学报, 2014, 26(12): 3731-3739.
HE Shaojun, ZHAO Shujuan, LI Jing, et al. Effects of betaine on growth performance, duodenal digestive enzyme activity and cecal microflora in Heat Stressed Broilers [J].JournalofAnimalNutrition, 2014, 26(12): 3731-3739.
[19] 龙凡, 梅文亮, 许兰娇, 等. 烟酸的生物学功能及其在畜禽生产中的应用[J]. 动物营养学报, 2014, 26(12): 3731-3739.
LONG Fan, MEI Wenliang, XU Lanqiao, et al. Biological function of niacin and its application in livestock and poultry production[J].JournalofAnimalNutrition, 2014, 26(12): 3731-3739.
[20] Kim H S, Xiao C Y, Wang R H, et al. Hepatic-specific disruption of SIRT6 in mice results in fatty liver formation due to enhanced glycolysis and triglyceride synthesis[J].CellMetabolism, 2010, 3(12): 224-236.
[21] Ding H Y, Li Y, Liu L H, et al. Sirtuin 1 is involved in oleic acid-induced calf hepatocyte steatosis via alterations in lipid metabolim-related proteins [J].JournalofAnimalScience, 2021, 10(99): 250.
[22] Kamanna V S, Ganji S H, Kashyap M L. Recent advances in niacin and lipid metabolism [J].CurrentOpinioninLipidology, 2013, 3(24):239-245.
