原位冷冻光电关联技术的研究进展
2023-01-07李尉兴徐晓君
卢 婧,李尉兴,徐晓君,纪 伟
(中国科学院生物物理研究所, 北京 100101)
1 引 言
细胞是所有生物的基本功能单位,而细胞功能很少由单一种类的分子完成,大多数细胞功能起源于细胞内不同种类分子之间的相互作用,很多重要的分子复合体根植于它们所在的细胞环境中,无法在不破坏其结构完整性的同时将它们剥离出来。因此,在细胞原位对生物大分子进行观察和结构解析,研究生命活动和疾病入侵的分子机理,既有助于揭示生命现象本质,也关系到国家生物安全、医药卫生、农业和绿色产业发展等重大战略需求。
冷冻电子断层扫描(cryo-ET)是在细胞原位解析生物大分子结构的核心技术[1]。在该过程中,细胞样品被迅速冷冻固定(通常是液氮温度-196°),其中的水跳过结晶态直接进入玻璃态,因此,细胞中的细胞器和大分子复合体得以维持其原始位置和形态;样品制备完成后,送入冷冻透射电镜,通过将样品倾斜不同角度,采集一系列的二维图像,重建出样品的三维图像,可以得到0.1纳米量级的分辨率。然而,cryo-ET要应用于细胞原位检测,还面临3个技术难点:第一,受限于电子穿透深度,样品厚度需要小于300nm[2],大多数细胞样品无法直接成像;第二,电镜图像的衬度来源于样品质量密度分布,而细胞内环境非常嘈杂,很难精确定位所需的生物大分子[3]。第三,很多生物过程在细胞中呈稀疏分布,无法单纯根据电镜图像定位。因此,要在细胞原位进行cryo-ET成像,需要精确定位目标区域,并将样品减薄到300 nm以下。
聚焦离子束(FIB)减薄是目前最有前景的冷冻样品减薄方式[4],FIB通常集成在扫描电镜(SEM)腔室内。聚焦至纳米量级的离子束(常用Ga+离子)接触样品表面后,会溅射出部分样品,并气化为二次离子或是中性原子,从而实现对样品的切割减薄。通过将离子枪和样品设置成一定角度,并对细胞中感兴趣区域上下表面进行切割减薄,可以得到透射电镜所需要的300 nm以下的平整薄片样品[5-6],再将制备好的薄片样品通过冷冻传输系统送入冷冻透射电镜进行断层扫描成像。由于生物样品中大多数都是碳、氢、氧等轻原子,为了避免样品切割范围外的表面损伤,离子束减薄时的电流很小,相应也会花费更长的时间;样品从扫描电镜腔室转移到透射电镜腔室,需要全程处于低温低湿状态,同样费时良久,且样品有一定的污染概率。综上所述,FIB虽然可以将冷冻细胞样品减薄到300 nm以下,但为了提高透射电镜样品制备成功率,需要精确高效地定位目标分子所在区域。
荧光显微成像可以通过用染料或荧光分子标记细胞中特定的蛋白或者亚细胞结构,实现对生物大分子和分子复合物的定位及对其生物过程的观测。将荧光显微镜和电镜结合起来的方式被称为冷冻光电关联显微成像(cryo-CLEM),将荧光显微镜对目标分子的定位优势和电镜的分辨率优势相结合,通过光镜图像和电镜图像的配准,在复杂的细胞环境中定位感兴趣的细胞器或分子复合物,并在原位对其进行观察和解析。为了保证光电关联图像精确配准,细胞冷冻需要在光镜成像之前进行,因为在冷冻过程中可能出现样品载网变形,导致细胞形态变化,而且对于活细胞来说,在光镜成像到原位冷冻期间,内部结构功能也会发生改变。
2 原位冷冻光电关联成像概述
cryo-CLEM成像流程通常如图1(彩图见期刊电子版)所示,首先将生物细胞培养在电镜载网上(图1(a)),并对细胞中感兴趣的结构或过程进行荧光标记(图1(b)),再将载网迅速降低到液氮温度以下(图1(c)),然后将样品转移到冷冻光镜中进行荧光成像,并选择需要减薄的目标位置(图1(d)),接下来,将样品传输到FIB-SEM双束电镜下,将光镜图像和电镜图像进行关联,并将目标位置FIB减薄到300 nm以下(图1(e)),最后将样品转移到透射电镜下进行cryo-ET成像(图1(f))。

图1 原位冷冻细胞光电关联成像示意图。(a)细胞培养;(b)荧光标记;(c)原位冷冻;(d)光镜成像;(e)FIB减薄;(f)cryo-ET成像Fig. 1 Schematic diagram of cryo-CLEM. (a) Cell culturing; (b) fluorescent labeling; (c) fast freezing; (d) fluorescent imaging; (e) FIB milling; (f) cryo-ET imaging
冷冻荧光成像可以精确定位标记生物分子的位置,可是仅对荧光标记的分子种类敏感,缺乏详细的细胞背景,而冷冻电镜则对生物样品中所有分子的电子密度敏感,能够提供详细的细胞背景,但它缺乏广泛适用的、特异性强的标记方法。Cryo-CLEM结合了荧光图像的特异性标记,以及电镜图像的电子密度敏感特性,是一项发展迅速的新兴技术。已经成功用于观测植物细胞中的自噬体[7]、动物细胞中的微管等细胞骨架[8-9]、多层膜结构[10-11]、酵母菌中的质膜隔室[12]和蛋白自噬机制[13]、果蝇脑神经[14]以及神经细胞突触[15]等。
3 原位冷冻技术
为了在cryo-ET下实现生物大分子的结构解析,细胞的原位冷冻固定是样品制备的第一个技术难点。如果将生物样品缓慢降温,其中的水会形成结晶态的冰,结晶态的冰会导致电子束发生散射,不仅会降低透射电镜图像对比度,还会破坏细胞中分子复合物的结构。因此,为了避免细胞中的水在降温过程中出现结晶态,需要使细胞中的温度迅速降低,形成一种玻璃态的冰,这种冰对电子束来说是透明的,不会影响电镜图像质量,而且能完整保存细胞中分子复合物的结构,这就是原位冷冻技术[16]。
目前,根据样品的厚度和种类,原位冷冻技术主要分为快速冷冻和高压冷冻两种。快速冷冻技术主要应用于厚度小于15 μm的样品,其原理是将铺设了样品的电镜载网固定在栓塞上,通过重力驱动将载网迅速浸入冷却腔室,此时腔室中注满了由液氮冷却的乙烷或者乙烷和甲烷的混合物,通过热传导对生物样品进行快速冷冻,这些液体的高热容量、较高的沸点和导热性可以保证冷却速度。很多商用的快速冷冻仪器可以实现自动化冷冻控制和环境参数调节,极大地提高了生物样品的制备效率。例如:FEI公司推出的VitrobotTM系列(图2(a)),Gatan公司的CP3系列(图2(b))等。
而对于超过15 μm厚的细胞样品或者组织样品,由于水本身的导热性较差,无法通过快速冷冻技术实现整个样品的快速降温,故需要采用高压冷冻固定(HPF)的方式[17]。当环境压力增加时,水的融化温度降低,而高压冷冻固定技术可以在几毫秒的时间内对样品施加2×108Pa压力,将水的融化温度降低至-40 °C。高压冷冻固定技术通过降低水的冰点避免形成冰晶,其可以使~250 μm厚度的样品均匀玻璃化[18]。常用的商用高压冷冻仪包括Leica公司的EMPact系列(图2(c)),Bal-Tec公司的HPM系列等(图2(d))。

图2 商用原位冷冻设备。(a)FEI公司VitrobotTM;(b)Gatan公司CP3;(c)Leica公司EMPact;(d)Bal-Tec公司HPMFig. 2 Commercial plunge freezers and high pressure freezers. (a) VitrobotTM from FEI; (b) CP3 from Gatan; (c) EMPact from Leica; (d) HPM from Bal-Tec
4 原位冷冻光电关联系统
4.1 冷冻样品台技术
冷冻光镜和冷冻电镜的关联,依赖于高稳定性的冷冻样品台。此外,对冷冻光镜的物镜也有特殊需求:首先,冷冻光镜的物镜大多使用空气物镜,因为绝大多数商用油浸显微物镜都不适用于低温;其次,物镜的工作距离应该尽量长,因为物镜相对于样品的温度更高,如果距离样品太近,会导致样品结晶或是物镜发生损伤;最后,为了达到更高的分辨率,物镜的数值孔径需要尽量大。世界上第一台用于原位冷冻细胞荧光成像的冷冻样品台(简称“冷台”)由Sartori等人开发,并放置于一个倒置的光学显微镜上[19]。随后,他们和FEI公司合作开发了第二代冷台,并于2010年发布。这个冷台可以使用工作距小于2 mm的显微物镜,冷台中的自动液氮泵可以保证EM载网上的样品处于玻璃态,其一次最多可以加载4个样品载网。同时,Schwartz等人开发了一台可放置于正置光学显微镜上的冷冻样品台[20],随后和Instec公司合作开发了CLM77K系列冷台(图3(a)),为了确保样品不升温,该冷台只能使用工作距为5 mm以上的显微物镜。Linkam公司也在同期开发了适用于多数正置光学显微镜的冷台CMS196(图3(b)),在该冷台中,内置液氮杜瓦瓶的设计使得冷台不需要连接液氮泵,同时可对显微物镜进行降温,使得CMS196可以使用NA为0.9,工作距离仅有300 μm的显微物镜。Leica公司基于Schorb和Brigg的工作[21],开发了商用冷台。该冷台通过断开液氮罐和冷台的直接连接,减少了氮气沸腾和泵送过程中产生的震动。针对冷冻荧光成像,Leica公司还开发了适用于低温工作的空气物镜,在物镜前端使用热导率较低的陶瓷材料,使得物镜的NA可达0.9,工作距离仅有280 μm。2016年,FEI公司开发了适用于倒置显微镜的冷台CorrSight(图3(c)),可以使用NA为0.9的物镜,由于物镜和样品被一个玻璃窗口片隔开,该冷台可以使用任意工作距离大于410 μm,且经过标准盖玻片校正的商用空气物镜。

图3 几种商用冷台和冷台样机实物图。(a)Instec公司的CLM77K;(b)Linkam公司的CMS196;(c)FEI公司的CorrSight;(d)Li等人提出的高稳定性冷台[19];(e)徐涛组提出的高稳定冷台[20]Fig. 3 Commercial cryo stages and prototypes. (a) CLM77K from Instec; (b) CMS196 from Linkam; (c) CorrSight from FEI;(d) Cryo stage proposed by Li[19]; (e) Cryo stage proposed by Xu[20]
上述冷台存在的一个共同问题:较低的温度稳定性和机械稳定性。为了解决这个问题,研究者提出将换样机构和冷台的温度同机械稳定性耦合,以提高冷台的稳定性,进而提高光镜成像分辨率和敏感度。但是这种设计对物镜工作距离有较高要求[22-24]。Hessel等人设计的冷台采用了长工作距物镜,可以在液氦温度(1.6 K)下使用,光路相对简单,但是如果采用高NA物镜,会引起很大的球差和色差[24]。Li等人提出的冷台结构如图3(d)所示,将物镜放置在冷台外的室温中,物镜和冷台之间用光学窗隔开,因此,需要采用更长工作距离的物镜,其能使用的最大NA物镜是0.7[22]。国内方面,中国科学院生物物理研究所的徐涛课题组在2018年报道了一台具备极高的温度稳定性(<0.06 K/10 h)和机械稳定性(<200 nm/5 h)的冷台(图3(e)),使用的商用物镜的NA为0.8,工作距离为3 mm,其采用该冷台实现了单分子定位成像[23]。
4.2 原位冷冻光电关联成像系统
根据原位冷冻光镜相对于电镜的位置,可以将cryo-CLEM成像系统分成两类,一类是分体式cryo-CLEM系统(图4(a)),其冷冻光镜独立于FIB-SEM电镜,样品在冷冻光镜下成像后,通过冷冻传输系统传输到电镜真空腔室内,再进行电镜成像和FIB切割减薄,减薄后的样品薄片再经由冷冻传输到cryo-ET中;另外一类是嵌入式冷冻光电关联系统,其光镜部分嵌入电镜腔室内,冷冻样品在光镜下成像后直接在真空腔室切换到电镜下成像,而且在FIB切割减薄过程中,可以随时将样品移动到光镜成像位置,样品薄片制备完成后,再通过冷冻传输到cryo-ET中。
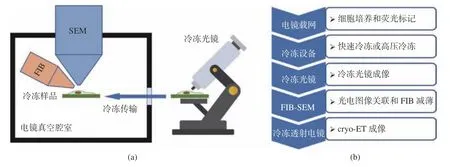
图4 (a)分体式cryo-CLEM的成像系统示意图;(b)分体式cryo-CLEM的成像流程Fig. 4 (a) Schematic diagram and (b) flow chart of imaging process of independent cryo-CLEM system
4.2.1 分体式冷冻光电关联成像系统
如图4(b)所示,分体式冷冻光电关联成像流程如下:首先,在电镜载网上培养细胞和荧光标记;然后,将载网放入冷冻设备中进行快速冷冻或高压冷冻;接下来,通过冷冻传输将样品传到冷冻光镜中进行光镜成像,再通过冷冻传输将样品传入FIB-SEM中,关联光镜和电镜图像,并用FIB切割减薄;最后,将样品冷冻传输到冷冻透射电镜中,进行cryo-ET成像[22]。由于冷冻光镜独立于FIB-SEB双束电镜,可以采用更复杂的光路结构,引入各种超分辨成像模态[23]。
冷冻样品的光学超分辨成像仍然是一个较新的领域:一方面,荧光分子的光漂白(photo-bleaching)和光分解(photo-decomposition)效应在低温下会大大降低,甚至消失[19,24];另一方面,有些荧光蛋白的光转换(photo-switching)能力消失,有些虽然存在,也和室温下的条件差异很大;另外,很多超分辨方法需要使用较高的激光强度,可能会使样品局部升温,生成冰晶,导致生物样品损坏。下面介绍一些已经应用于冷冻样品成像的超分辨成像技术,如图5(彩图见期刊电子版)所示。
冷冻受激发射耗尽显微术(cryo-STED)采用两路激光同时进行扫描,一路是激发光,用于成像,另一路是STED光通过相位调制产生甜甜圈形状的光斑,用于抑制焦点边缘信号,从而获得更小的点扩散函数(图5(a)),其分辨率可达30~80 nm,且可以实现三维成像。有研究者发现低温下,cryo-STED的分辨率可以提升1.6倍[25],然而文献中的样品是缓慢降温,并未进入玻璃态,而且低温下所需的功率密度可达MW/cm2~GW/cm2。此外,还有研究证明300 W/cm2强度激光照射样品1~2分钟,就会破坏玻璃态,导致冷冻样品中产生冰晶。最近一些研究人员将STED成像应用于树脂包埋后的生物样品中,并与透射电镜图像进行关联[8,26-27],而在原位冷冻生物样品中的应用还有待进一步研究。

图5 几种超分辨冷冻荧光显微镜示意图。(a)冷冻STED成像;(b)冷冻单分子定位成像;(c)冷冻结构光照明成像;(d)冷冻Airyscan成像Fig. 5 Schematic diagrams of cryo supper resolution fluorescent microscopy. (a) cryo-STED; (b) cryo-SMLM; (c) cryo-SIM;(d) cryo-Airyscan
基于荧光分子的光转换能力,研究人员提出了冷冻单分子定位成像(cryo-SMLM)。该技术使每次采集图像仅有少量的单个荧光分子发光,并准确定位每个荧光分子的点扩展函数质心,再将多张图像叠加形成一幅超高分辨率的图像(图5(b))。冷冻单分子定位成像包含冷冻光激活定位(cryo-PALM)、冷冻随机光学重建(cryo-STORM)、冷冻荧光活化定位(cryo-fPALM),冷冻光学波动成像(cryo-SOFI)等,分辨率取决于探测到的单个荧光分子的光子数。目前已有多种单分子荧光定位成像应用于冷冻样品中[28-32]。通过采用长工作距,较高数值孔径(0.75~0.8)的空气物镜,提供了出色的分辨率(75~125 nm)。2021年,Andrian等人采用DNA-PAINT单分子成像技术和透射电镜成像关联,实现了对纳米颗粒的高精度光电关联[33]。然而,要达到常温单分子定位成像的纳米级别分辨率[34-35],对于原位冷冻生物样品来说,还面临许多挑战。首先,cryo-SMLM高度依赖于荧光分子在低温下的转换特性[36],前期样品制备较为复杂。针对这一问题,Robichaux等人在2019年将老鼠视网膜做成超薄切片,并进行cryo-STORM和cryo-ET成像,在感光细胞中发现了以前未知的亚结构,并揭示了关键蛋白的分布[37];Moser等人于2019年采用cryo-SOFI成像对哺乳动物细胞进行成像,并与cryo-ET图像进行关联,由于SOFI的成像不需要光漂白或者光开关,所需的激发功率更低,能够有效降低样品结晶的风险[38];Hoffman等人在2020年发表了有关原位冷冻多色单分子光电关联成像的研究成果,他们采用液氦作为冷源,使用蓝宝石盖玻片支撑样品,获得了可以与透射电镜精准关联的高分辨光镜图像[29]。其次,长时间的高强度激光照射,导致样品温度升高,存在玻璃态被破坏的风险,针对这一问题,Liu等人采用方华膜载网代替炭膜载网,以降低载网从激光中吸收的热量,这样可以将用于单分子成像的光功率增加到1.75 kW/cm2,并获得了75 nm的分辨率[32]。可是,由于方华膜对电子的吸收率较高,导致电镜成像对比度和分辨率降低。此外,单分子成像的定位精度与收集到的光子数开方成正比,要实现超高分辨率,需要大量图片叠加,耗时良久,单分子成像视场通常小于50 μm×50 μm,在光电关联应用上,通常需要进行视场拼接,这进一步增加了成像时间。这也是限制冷冻单分子定位成像应用的主要因素。
冷冻结构光照明显微成像(cryo-SIM)采用空间光调制器生成周期性条纹的激发光,并将3个不同方向的条纹光照射到样品上,再利用算法重建出含有高频信息的图像,其横向和轴向分辨率相比宽场成像可以提高2倍(图5(c))。由于多数荧光蛋白低温下的荧光强度更高,更不易漂白,基于此,cryo-SIM已经成功应用到冷冻样品成像中[29]。然而,传统的SIM成像对调制照明光路像质要求非常高,容易在图像重建中出现伪像。对原位冷冻光镜成像来说,显微物镜仅能使用长工作距的空气物镜,加上物镜到样品焦面真空和玻璃态水的折射率偏差,SIM系统的像质难以保证。Arnold等人将在光镜和电镜中均能成像的铁磁小球铺在生物样品上,利用小球的相对位置对光镜和电镜图像进行配准,将图像关联精度提高到200~300 nm[6]。Hoffman等人则首先在液氦温度下,通过cryo-SIM得到光镜图像,再用冷冻树脂替代样品,送入FIB-SEM双束电镜中进行切片成像,最后利用样品中的荧光小球进行光电关联配准[29]。Phillips等人开发了一套3D cryo-SIM成像系统,采用NA为0.9,工作距离为2 mm的物镜。对于488 nm激发光的荧光小球,成像横向分辨率和轴向分辨率分别为210 nm和640 nm[39]。
冷冻Airyscan(cryo-Airyscan)显微成像是一种基于像素重构原理(Pixel Reassignment)的超分辨成像技术,它源自生物光学成像的金标准——共聚焦显微成像技术。cryo-Airyscan将原先的针孔替换为阵列探测器。阵列探测器的每个单元独立成像。通过对每个单元图像解卷积和移位线性运算,可以重建出最终的超分辨图像,其分辨率相对于传统共聚焦成像(未做解卷积)提高了1.7倍(图5(d))。如果将每个探测单元的图像不经过移位直接相加,就得到了共聚焦图像。cryo-Airyscan成像与cryo-SIM成像类似,均可使冷冻样品的荧光强度提高,光漂白效应减弱,并且已经成功应用到冷冻生物样品中[40]。此外,由于Airyscan采用点扫描和有限尺度探测的成像方式,在图像重建之前,就已经有效过滤了焦面外的背景荧光,更适用于荧光分子分布更密集的厚样品。Zeiss公司开发了一套基于Airyscan的cryo-CLEM成像系统,并成功应用于FIB-SEM和Cryo-ET[10,41-42]。
国内方面,中国科学院生物物理研究所成像平台开发了一套基于宽场成像的cryo-CLEM系统HOPE[43],其硬件主体是一台高真空冷台。其可以搭载在宽场荧光显微镜上,实现冷冻荧光成像,并可与透射电镜冷冻样品杆连接。由于机械结构的限制,该冷台只使用了10倍和40倍的空气物镜。此外,徐涛课题组还开发了一套基于单分子成像的冷冻超分辨sCLEM成像系统[20,44]。该系统采用开放式冷冻样品台,其收集端使用柱透镜以提高轴向分辨率,并引入实时漂移校正(校正精度约5 nm)模块。该系统可以使用NA为0.8,工作距离为3 mm的空气镜,横向分辨率可达13 nm,轴向分辨率约为40 nm。
4.2.2 嵌入式冷冻光电关联成像系统
嵌入式冷冻光电关联成像流程如图6(b)所示。首先在电镜载网上培养细胞和荧光标记,再将载网放入冷冻设备中进行快速冷冻或高压冷冻,然后通过冷冻传输将样品传到嵌入式光电关联设备中进行光镜成像、光电图像关联和FIB切割减薄,最后将样品冷冻传输到冷冻透射电镜中,进行Cryo-ET成像。对比分体式系统,嵌入式系统的流程中少了将样品冷冻传输送入光镜和FIB-SEM的过程,而是直接在电镜舱室内完成冷冻光镜成像、光电图像关联和FIB减薄。由于在冷冻传输过程中部分样品存在升温的可能,减少冷冻传输的步骤不仅可以简化成像流程,提高效率,还能减少样品污染的风险。另外,由于冷冻光镜和电镜成像位于同一腔室,可以方便快捷地在两种成像模式中进行切换。FIB的减薄过程,也能够用冷冻光镜进行监控,从而提高减薄的准确率。同时,嵌入式冷冻光镜由于部分嵌入双束电镜中,受限于电镜腔室大小和外形尺寸,很难采用复杂的光路结构,现有的嵌入式冷冻光镜大多采用比较简单的成像模式。

图6 嵌入式cryo-CLEM成像系统的(a)示意图和(b)成像流程Fig. 6 (a) Schematic diagram and (b) block diagram of imaging process of integrated cryo-CLEM system
Faas等人最早将一个宽场显微镜集成到透射电镜中,并应用于观测冷冻样品,命名为iLEM[45]。其可以通过荧光位置判断样品薄片上的感兴趣区域。iLEM的主要限制因素是透射电镜腔室内的空间有限,冷冻光镜的分辨率很低,而且采用光电关联成像的重要原因是这种结构无法实现荧光导航样品减薄。后来,Gorelick等人提出将一个宽场光学显微镜集成到一台商用双束电镜中。其可以使用若干不同的商用显微物镜,最高NA为0.95,工作距为0.33 mm,可实现对原位冷冻样品的高分辨成像[5],同时通过采用铁磁小球进行光电图像配准,指导FIB减薄。随后,Delmic公司开发了一套嵌入式光电关联成像系统——METEOR,该系统可以外挂在商用FIB-SEM电镜舱室上,并通过真空法兰将光路导入电镜腔室内[46],从而实现多色宽场荧光成像,并指导FIB减薄[47-48]。
5 总结与展望
自从Schwartz等人首次将冷冻荧光成像和冷冻电镜成像相结合[49],cryo-CLEM技术已经发展了15年,它结合了冷冻电镜成像的高分辨能力和荧光成像的特异性标记能力,随着细胞原位蛋白解析在生物学上的应用越来越广泛,cryo-CLEM及其派生的荧光导航减薄技术有着广阔的应用前景。然而,作为一项新兴技术,cryo-CLEM还远不够成熟,有很多挑战有待克服。
cryo-CLEM目前有两条技术实现路线:分体式和嵌入式cryo-CLEM成像。分体式cryo-CLEM成像中的冷冻光镜独立于电镜存在,其可以在现有的荧光显微镜上嵌入冷冻样品台,目前已经开发出多种成像模态。目前面临的技术瓶颈主要是冷冻样品台的机械稳定性和温度稳定性,尤其对于超分辨冷冻荧光成像,例如对于cryo-SMLM,冷冻样品台需要达到纳米级别的稳定性,才能获得纳米量级的分辨率,才能更好地与cryo-ET图像匹配[50]。此外,绝大多数冷冻荧光成像使用的都是空气物镜,数值孔径有限,这影响了冷冻荧光成像的分辨率和信噪比。因此,研制能够在冷冻条件下使用的浸没式物镜,进一步提高数值孔径和光子接收效率,也是未来的发展方向。
除了硬件技术本身的限制,冷冻样品本身也是限制冷冻超分辨荧光成像的因素。首先,cryo-SMLM和cryo-STED都需要很高的局部功率密度才能产生光漂白、光转换或者光损耗效应,而过高的激光功率密度产生的热效应会使得冷冻样品发生局部结晶,从而破坏冷冻透射电镜的成像效果。因此,cryo-SMLM和cryo-STED的激发功率密度远低于室温下的SMLM和STED成像,对于cryo-STED来说,较低的STED光功率限制了成像的分辨率,而对于cryo-SMLM来说,如果没有光漂白或是光转换效应,则无法进行单分子成像。在较高激光功率密度下,如何及时给冷冻样品降温,避免样品局部结晶是一个可行的思路。其次,在低温冷冻条件下,很多荧光探针的特性发生了改变,大多数探针的荧光效率更高,同时更难被漂白或发生光转换效应,这对cryo-SIM和cryo-Airyscan这类成像来说是有利因素,而对cryo-SMLM来说则是不利因素,因此,探索单分子荧光探针的低温特性,开发能够在冷冻条件下使用的单分子探针也是未来的发展方向之一。
对于cryo-CLEM来说,将冷冻光镜和电镜模式下所成图像进行精确配准,是成功实现原位光电关联和荧光导航离子束减薄的必要步骤。光电关联图像配准通常分为两步。第一步是粗略配准,通过使用带有坐标标记的电镜载网实现;第二步是精确配准,目前常用的方法是在生物样品制备过程中,铺设在荧光成像和电镜下都能看到的铁磁珠、金颗粒或聚苯乙烯小球,再选取一些小球作为基准,将两幅图像进行配准。由于FIB图像相对于SEM和荧光图像有一个夹角,而且最终的样品薄片小于200 nm,故要实现精准的FIB减薄,需要将荧光图像和FIB图像进行精确的三维配准。理论上来说,只要选取的配准点足够多,就能够实现高精度的三维配准,可是在实际使用中还存在很多问题。例如在分体式cryo-CLEM中,从冷冻荧光成像到冷冻电镜成像,需要经过冷冻样品转移。转移过程可能会改变样品的局部形态,导致配准失败。此外,在配准点中心的选择、目标中心的选择等人工过程中也存在误差,这些都会对配准精度产生影响。针对上述问题,提高原位冷冻光电关联图像配准精度是未来的重要发展方向。
另一种cryo-CLEM成像技术路线—嵌入式cryo-CLEM成像,是将冷冻光镜集成在FIBSEM电镜腔室内,少了一个冷冻样品传输步骤,故可以降低样品结晶和污染的风险,且在FIB减薄过程中,可以方便地检查荧光状态,较分体式cryo-CLEM更有优势。嵌入式cryo-CLEM还处于发展的初级阶段,光镜模态目前仅限于宽场成像,分辨率较低,且缺失三维信息,主要限制因素是电镜腔室内空间有限,而光镜的其他模块如果外挂在电镜主体上,可能影响电镜的机械稳定性。因此,下一步需要做的工作包括:第一,扩展嵌入式cryo-CLEM中光镜的成像模态,引入三维成像和超分辨成像,提高光镜的成像分辨率;第二,使嵌入式cryo-CLEM为荧光导航FIB减薄提供更多信息,便于检查样品减薄过程中的荧光,为了提高荧光导航减薄的精度和准确性,需要发展新的光电关联配准流程和方法。
