格陵兰努普·康格鲁阿峡湾叶绿素a浓度时空变化分析
2023-01-07田佳慧程晓张彦南
田佳慧 程晓,2,3,4 张彦南
研究论文
格陵兰努普·康格鲁阿峡湾叶绿素浓度时空变化分析
田佳慧1程晓1,2,3,4张彦南2,4
(1北京师范大学全球变化与地球系统科学研究院遥感科学国家重点实验室, 北京 100875;2中山大学测绘科学与技术学院, 广东 珠海 519082;3南方海洋科学与工程广东省实验室(珠海), 广东 珠海 519080;4中国高校极地联合研究中心, 北京 100875)
努普·康格鲁阿(Nuup Kangerlua)峡湾的叶绿素(Chl)浓度的时空分布对于气候变化有一定的指示作用。基于此, 结合通过C2RCC算法对Sentinel-3 OLCI反演得到的Chl数据、OSTIA海表面温度(SST)数据以及Nuuk气象站数据, 对2016—2020年该区域的Chl浓度分布进行时空分析。结果表明, 在峡湾入口端, Chl的分布主要受风力和海温的影响, 在9月风力最强盛时达到最大; 而在冰川出水口端, 除风力和水温外, 在冰川融水的作用下, 浮游植物在春季爆发后, 还伴随有秋季二次爆发。
峡湾 叶绿素Sentinel-3 OLCI C2RCC
0 引言
随着全球气候变化的不断加剧, 格陵兰及其周围海域的水文特征(温度、盐度、混合层等)以及整个生态系统正在发生着根本性的变化[1]。格陵兰的峡湾系统是连接海洋和冰盖的纽带[2], 近几十年来, 温度升高[3-4]和格陵兰周围夏季云量的减少[5]导致表面融化的增加以及海洋终结冰川的加速退缩[6-7]正在对峡湾系统产生复杂的影响。
叶绿素(Chl)是海洋初级生产力和水体富营养化的指标[8-9], 大气条件、水文特征和一系列其他过程的相互作用可能对Chl浓度的季节和年际变化产生复杂的影响[10-11]。受冰川融水影响的峡湾, 其养分供应与悬浮物供应是分开的, 养分供应主要来自峡湾口的海水, 悬浮物及淡水主要来自冰川融水, 但融水的稀释作用会导致较低的营养水平[12-13]。海水与冰川融水的混合会改变峡湾的营养盐分布, 融水径流的季节或幅度变化可能会进一步影响浮游植物优势群落[14-15]及Chl浓度的时空变化[16]。Chl浓度不仅与格陵兰峡湾生态系统初级生产力密切相关, 而且对于了解格陵兰冰盖变化对气候和海洋的影响具有重要意义。
目前, 已经有一些学者对格陵兰峡湾的叶绿素浓度展开研究, 但大多数都集中在利用实测数据研究峡湾中Chl的垂直浓度梯度, 且只针对努普·康格鲁(Nuup Kangerlua)峡湾北支, 缺乏对峡湾整体的研究[16-18]。单纯依靠传统的船舶采样方法或依靠浮标进行Chl浓度监测, 无法满足大尺度、时间连续的数据观测需求。光学遥感技术以其时间连续、成本低、便于推导水质参数等优点, 越来越多地用于水质监测[19]。
本文基于Sentinel-3 OLCI数据, 分析2016—2020年Nuup Kangerlua峡湾Chl浓度的时空变化规律以及气象和水文因素对Chl浓度产生的影响。为了解格陵兰峡湾内Chl的时空变化特征及规律提供参考。
1 研究区域
Nuup Kangerlua位于格陵兰岛西南部, 长190 km, 宽7 km, 峡湾总面积为2013 km2, 是格陵兰岛拉布拉多海沿岸最长的峡湾[20]。格陵兰的首府努克位于峡湾口附近。研究区域属于受海洋影响的苔原气候, 冬季寒冷漫长, 夏季凉爽短暂, 最高温通常出现在7月, 月均值为11℃。最低温通常出现在2月, 月均值为–10.9℃。年均降水量超过800 mm。一年中日照时间变化大, 日照时长范围为4~21 h。
Nuup Kangerlua峡湾系统具有复杂的环流模式, 峡湾口存在强烈的潮汐混合现象[21], 在冬季和春季会有近岸海水流入峡湾内部。峡湾被3个入海冰川包围, 从北到南分别为: Narsap Sermia(NS)、Akullersuup Sermia(AS)和Kangiata Nunaata Sermia(KNS)(图1)。由于3个入海冰川的接地深度不同(KNS, AS 和 NS分别约为<250 m, <140 m和<160 m), 位于峡湾内的两条冰川(KNS和AS)与峡湾外部盆地的水体交换水平较低[22]。夏季, 峡湾会受到来自这些冰川融水的影响[23]。

图1 研究区域。2017年7月21日格陵兰努普·康格鲁阿的Sentinel-3 OLCI RGB(波段 8-6-4)合成
Fig.1. Area of study. RGB (bands 8-6-4) composite of Sentinel-3 OLCI image on 21 July 2017 of Nuup Kangerlua on the southwest coast of Greenland
2 数据与方法
2.1 Sentinel-3A OLCI数据
Sentinel-3是欧洲航空局和欧洲气象卫星组织联合运营的卫星项目, 可以提供海洋和陆地观测服务, 共有两颗卫星, 分别为Sentinel-3A和Sentinel-3B。其中, Sentinel-3A卫星于2016年2月发射, 配备了海洋和陆地彩色仪器(OLCI)传感器。OLCI的空间分辨率为300 m, 在高纬度地区的重访时间小于2天。OLCI在MERIS的基础上增加了6个波段, 共设置了21个光谱波段(0.4~ 1.02 µm), 提高了荧光光谱反演的准确性。目前, 大量学者使用Sentinel-3数据进行海洋监测。由于只能获取近一年的OLCI Level-2全分辨率水产品(OL_2_WFR), 为了尽可能延长研究期限,确保数据的一致性, 本研究采用OLCI Level-1B全分辨率产品(OL_1_EFR)。
由于峡湾的冬季(11月—次年3月)生产力水平较低[16], 以及出于对数据质量的考虑, 我们选择了2016—2020年的4—10月作为本研究的时间范围。由于Chl浓度在峡湾变化迅速, 我们选择尽可能多的Sentinel-3 OLCI图像进行时空变化研究。通过目测, 选取2016年至2020年4—10月Sentinel-3 OLCI Level-1B全分辨率 (OL_1_EFR)的207个无云场景和少云场景。数据来自哥白尼开放获取中心(https://scihub.copernicus. eu/)。
2.2 补充数据
2.2.1 水文数据
1. 盐度、浊度和Chl浓度实测数据
使用Zielinski等[24]在MSM65(Greenhab II)航行期间(2017年6月29日—7月3日)收集的盐度、浊度和Chl浓度数据, 筛选了峡湾内7个位置(图1)的测量数据用于剖面绘制。实测时间及点位对应如表1所示。
2. 海表面温度数据(SST)
本研究使用了Operational Sea Surface Temperature and Ice Analysis(OSTIA)系统的海表面温度数据。OSTIA海表面温度数据由英国气象局(https://www.metoffice.gov.uk/)运营, 是欧盟全球环境和安全监测(GMES)MyOcean项目的一部分[25]。该数据具有较高的时空分辨率, 可估算出0.05°的每日全球海表温度[26]。与原位测量结果相比, OSTIA SST的精度约为0.57 K[27]。OSTIA海温数据可从哥白尼海事处(https://marine.copernicus. eu/)获得。

表1 实测时间及点位
2.2.2 气象数据
气象站数据来自格陵兰生态系统监测(GEM)数据集(https://data.g-e-m.dk)。GEM是对北极生态系统和气候变化的影响和反馈进行全面监测和长期研究的项目。其中, 努克的Asiaq- Greenland测量项目为我们提供了气象数据。该数据集提供了从2007年10月至今每10分钟的风向和风速记录。我们采用每10分钟平均的风向和风速进行辅助分析。
2.3 C2RCC算法
由于峡湾内部会受冰川融水和潮汐的影响, 水质较为复杂, 这对遥感影像的大气校正和Chl浓度反演研究提出了更高的要求[28]。
Case 2 Regional Coast Color(C2RCC)算法最初由Doerffer和Schiller[29]提出。用于推导中分辨率成像光谱仪(MERIS)在二类水体中的固有光学特性(IOPs)。除IOPs外, C2RCC还可用于推导不同组分的吸收和表征, 三种主要的光学相关的浓度(浮游植物色素、总悬浮物TSM和有色可溶性有机物)以及与之相关的不确定性。
C2RCC采用了神经网络技术。美国国家航空航天局(NASA)在全球典型海域收集了大量海洋生物光学算法数据集(约500万例)参与神经网络训练, 涵盖沿海和内陆地区的大部分情况。C2RCC的优点之一是它的应用速度远高于其他神经网络算法, 因为它不需要任何迭代。目前已成为欧洲航天局为Sentinel系列卫星开发的处理软件SNAP[30]专题水处理工具包中的重要模块。该算法已应用于Sentinel-3 OLCI、Sentinel-2 MSI、Landsat 8、MODIS等卫星数据的处理, 关于C2RCC的更多信息, 可从SNAP的官方网站(http://step.esa.int/main/toolboxes/sentinel-3-toolbox/)获得。
图2为实测与C2RCC处理获得的Chl浓度值回归。为避免异常值干扰, 以实测点为中心, 将直径为300 m的像元均值与实测值进行匹配。由C2RCC获得的Chl浓度普遍高于测量值, 但仍有中度相关关系(2=0.684), 可以用于判断空间分布趋势。这与根据Vanhellemont和Ruddick[31]的观点一致。

图2 实测与C2RCC算法反演的Chl a回归
Fig.2. Regression of Chlmeasured in situ against Chlderived from C2RCC algorithm
3 结果分析
3.1 叶绿素a浓度的空间变化
3.1.1 海表面叶绿素浓度分布
通过反演, 我们得到了2016—2020年的Chl浓度空间分布, 图3为2017年4—10月每月平均Chl浓度, 在每年的Chl浓度基础上进一步将当月所有数据进行5年平均得到2016—2010年的4—10月Chl浓度空间分布序列(图4)。总体上, 2016—2020年Chl浓度对应月份的空间格局相似, 在远离冰川融水出口的水域, Chl的浓度主要在0.5~2 mg·m–3之间。4—10月, 峡湾整体Chl浓度呈现低—高—低特征。4月, 峡湾整体Chl浓度还不到1 mg·m–3。从5月开始, 最先与NS、KS、KNS冰川融水接触的峡湾北部支的Chl浓度首先增加。6—8月, Chl浓度从峡湾内部到峡湾口逐渐升高, 9月达到最大值, 总浓度在1.5 mg·m–3以上。10月, Chl的浓度迅速下降到1.25 mg·m–3以下。

图3 2017年4—10月每月平均Chl a浓度
Fig.3. The average Chlconcentration of each month from April to October, 2017

图4 2016—2020年月平均Chl a浓度
Fig.4. The average Chlconcentration of each month from 2016 to 2020
3.1.2 叶绿素浓度垂直分布
根据Zielinski等[24]2017年在Nuup Kangerlua的测量数据, 我们绘制了峡湾北部主要分支7个位置的剖面, 如图5所示(北部分支7个采样点的位置见图1)。在10 m以上深度, Chl浓度一般在1 mg·m–3以上。而6和7号位置10 m左右深度的Chl浓度显著高于同深度其他位置, 这与海冰及冰川冰的融化所产生的淡水径流有关。由于2017年7月, 冰川融水往峡湾内注入大量低营养盐度的淡水融水与峡湾内水体的密度差异导致密度分层, 产生了Chl在水下的高值区。而靠近峡湾口的1、2、3号点, 由于潮汐混合, 使得整个水柱均质化, 产生了较为均匀的浊度、盐度, Chl的分布进一步证实遥感数据与现场实测数据的空间趋势可以很好地吻合。
3.2 叶绿素a浓度的时间变化
由于C2RCC会高估高浑浊度水域的Chl浓度[31], 为避免悬浮物浓度对Chl浓度反演产生的影响, 我们选取峡湾北部分支中悬浮物浓度低于5 mg·L–1的区域进行分析。图6为2016—2020年的4—10月北部分支悬浮物浓度低于5 mg·L–1的Chl浓度与海表面温度的关系。其中, 灰色线为使用C2RCC从Sentinel-3 OLCI图像(峡湾北支TSM<5中的区域)中得到的每日Chl浓度的区域平均值。绿色线为对Chl每日浓度进行平滑后的结果, 更能代表Chl浓度变化的趋势。橙色线表示该地区的平均每日海表温度。
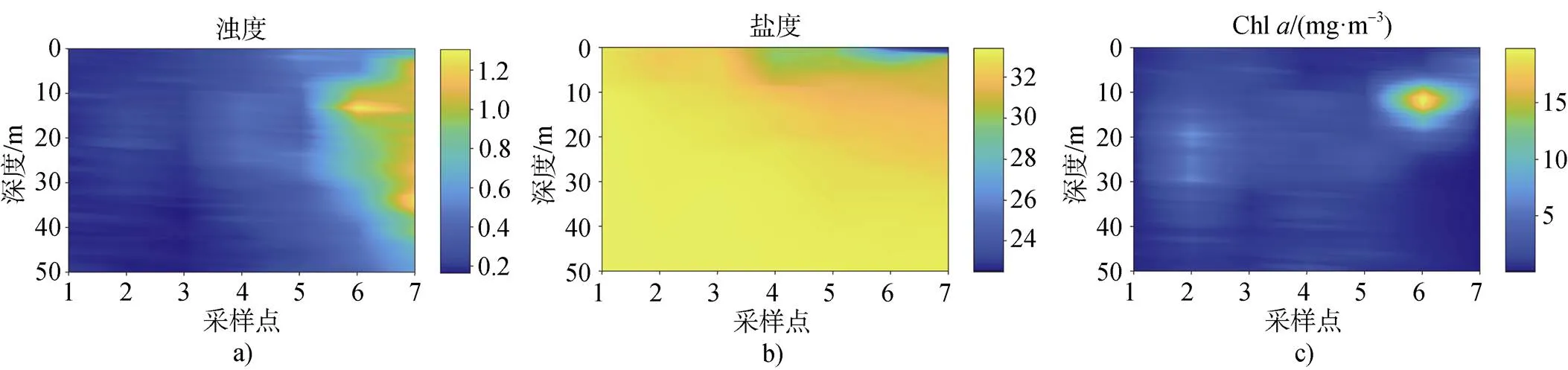
图5 基于实测数据的剖面图。a)浊度; b)盐度; c)Chl a浓度
Fig.5. Distribution profile based on the measured data. a)turbidity; b)salinity; c)Chlconcentration
Nuup Kangerlua的Chl浓度变化具有很强的周期性。对于峡湾北部分支整体而言, 2017年、2018年和2020年Chl浓度与海温的关系明显。2017年, 海温在8月中旬达到最大值(6.4℃), Chl浓度在9月初达到最大值(1.87 mg·m–3)。2018年, 海温在8月中旬达到峰值(6.2℃), Chl浓度在9月中旬达到峰值(1.75 mg·m–3)。2020年全年海温最大值出现在8月中旬, 达到7.5℃, 2020年Chl浓度也在9月中旬达到最大值, 为1.58 mg·m–3。因此, Nuup Kangerlua峡湾内Chl浓度与海温有关, 且Chl浓度峰值出现时间比海温峰值的出现时间晚一个月左右。

图6 2016—2020年逐日海温与Chl a浓度时间序列
Fig.6. Time series of daily SST and Chlconcentration from 2016 to 2020
为进一步研究冰川融水对Chl浓度的影响, 我们综合考虑了Mortensen等[22,32]对Nuup Kangerlua环流模式研究以及与KNS冰川的距离,将峡湾北部分支分为了3个区域(图1)进行统计区域内Chl与SST均值(图7), 分别为海槛区(区域1)与主要峡湾分支(区域2和区域3)。区域1接近峡湾口, 主要会受到由沿海海水与峡湾水体之间的密度差异导致的密集沿海流入影响。区域2产生了外槛区与峡湾盆地之间的密度梯度, 并形成斜压环流模式。区域3靠近入海冰川, 会受到入海冰川产生的淡水驱动, 淡水羽流上涌并与周围峡湾水体混合, 形成了冰下环流模式。

图7 2016—2020年分区统计逐日海温与Chl a浓度时间序列
Fig.7. Time series of daily SST and Chlconcentration in regional statistics from 2016 to 2020
如表2所示, 从峡湾口至接近KNS冰川融水的3个区域中, 接近入海冰川的区域3的海表面温度最大值范围为4.708~5.302℃, 是3个区域中SST最高值相对最低的区域。此外, 该区域为3个区域中Chl浓度高值最大的区域, 为1.33~2.01 mg·m–3。除2018年之外, Chl浓度高值均出现在5月中下旬, 这与冰川融水的汇入有关。随着区域3的Chl浓度逐渐降低, 区域2与区域1的Chl浓度逐渐升高, 其中, 区域2由于受到冰川融水与沿海径流流入的影响, 总体上Chl浓度在3个区域中变化幅度最小, 在6—9月均保持在1.12~1.65 mg·m–3范围内。

表2 2016—2020年每年海表面温度与Chl a浓度最大值及出现日期
3.3 叶绿素a浓度因素分析
Chl浓度会受到温度、风速、营养盐、冰川融水、浮游植物等气象和水文要素的影响。由于Chl浓度变化呈周期性以及受到实测数据的限制, 我们选择2017年进行影响因素分析。
4月, 靠近冰川融水出口的峡湾内部通常被冰覆盖(图8a), 峡湾整体平均海温接近0℃(图8b)。5月, 海表温度逐渐升高, 并于5月底达到3℃, Narsap Sermia冰川融水出口处海冰逐渐融化破裂, 与此同时, 破碎的冰水混合物在峡湾内东风(图8c)的推动下向西移动。6—7月, 海表温度持续上升, 峡湾内部主要受西风控制。海冰逐渐融化, 为峡湾内注入大量淡水, 此时峡湾内靠近融水出口的位置盐度明显低于同深度水平其他实测点(图5b)。在盛行西风的影响下(图8d), 高营养的海水从峡湾口流入峡湾内部, 与冰川融水产生的淡水混合, 为Chl浓度逐渐向峡湾口增加提供了有利条件。8—9月, 峡湾海表温度达到最大值(6℃), 东风逐渐增强。9月, 东风逐渐占上风。根据目视判断, 此时峡湾的海冰覆盖面积达到最小。10月, 海面温度持续下降, 峡湾内的海冰再次冻结, Chl浓度也相应下降。
总之, Nuup Kangerlua峡湾Chl浓度的空间分布会受到风和海表温度以及营养盐度的影响。峡湾外部的海水通过海风的作用涌入峡湾内部, 并通过潮汐混合为峡湾内提供了高营养水, 这为叶绿素浓度的增加提供了有利条件。
4 结论与展望
本文基于Sentinel-3 OLCI数据, 结合海温、盐度、风等水文气象因子, 分析2016—2020年格陵兰Nuup Kangerlua峡湾的Chl浓度时空变化, 得出如下结论。
1. 峡湾的Chl浓度时间变化具有明显的季节性。4—10月, 峡湾整体Chl浓度呈现低—高—低特征。从5月开始, 随着海表面温度升高, 海冰减退, 峡湾内靠近冰川融水出口处Chl浓度最先增加。6—8月, 峡湾整体Chl浓度逐渐升高, 9月达到最大值。10月, Chl浓度迅速下降。

Fig.8. Hydrology and meteorology of Nuup Kangerlua. a) evolution of sea ice conditions from April to October in 2017; b) sea surface temperature (°C) as recorded from OSTIA system (The red line is obtained by Gaussian smoothing, and the window width is 20); c) wind direction and speed as recorded at the Kobberfjord Climate Station from April to October in 2017; d) schematic of the hydrodynamic cycle of fjord in 2017 (The grey rectangle shows the distribution of ice in the fjord)
2. 从空间上看, 从冰川融水注水口到峡湾口, Chl浓度逐渐增加。在盛行风的影响下, 峡湾外部的海水涌入峡湾内部, 与冰川融水产生的淡水混合, 为峡湾内提供了高营养水, 这为叶绿素浓度向峡湾口增加提供了有利条件。
由于受限于峡湾实测数据的可获得性, 我们无法进一步测试由C2RCC反演得到的Chl浓度的准确性。通过对C2RCC反演值与已有实测数据的回归分析与其他学者对C2RCC算法在不同海域的 Chl反演精度研究, 虽然C2RCC与实测值存在差异, 但反演结果与实测值在时空变化上具有一致趋势。因此, 本研究只能初步分析Chl浓度在时空上的变化。希望在未来能有更多公开发表的数据可用来研究格陵兰峡湾。
1 秦大河. 气候变化科学与人类可持续发展[J]. 地理科学进展, 2014, 33(7): 874-883.
2 STRANEO F, CENEDESE C. The dynamics of Greenland's Glacial Fjords and their role in climate[J], Annual Review of Marine Science, 2015, 7: 89-112.
3 FETTWEIS X, BOX J E, AGOSTA C, et al. Reconstructions of the 1900–2015 Greenland ice sheet surface mass balance using the regional climate MAR model[J]. The Cryosphere, 2017, 11(2): 1015-1033.
4 STRANEO F, HEIMBACH P. North Atlantic warming and the retreat of Greenland's outlet glaciers[J]. Nature, 2013, 504(7478): 36-43.
5 HOFER S, TEDSTONE A J, FETTWEIS X, et al. Decreasing cloud cover drives the recent mass loss on the Greenland Ice Sheet[J]. Science Advances, 2017, 3(6): e1700584.
6 VAN DEN BROEKE M, BAMBER J, ETTEMA J, et al. Partitioning recent Greenland mass loss[J]. Science, 2009, 326(5955): 984-986.
7 ENDERLIN E M, HAMILTON G S. Estimates of iceberg submarine melting from high-resolution digital elevation models: Application to Sermilik Fjord, East Greenland[J]. Journal of Glaciology, 2014, 60(224): 1084-1092.
8 PAHLEVAN N, SMITH B, SCHALLES J, et al. Seamless retrievals of chlorophyll-from Sentinel-2 (MSI) and Sentinel-3 (OLCI) in inland and coastal waters: A machine-learning approach[J]. Remote Sensing of Environment, 2020, 240: 111604.
9 MOSES W J, GITELSON A A, BERDNIKOV S, et al. Estimation of chlorophyll-concentration in case II waters using MODIS and MERIS data-successes and challenges[J]. Environmental Research Letters, 2009, 4(4): 045005.
10 GAO S, WANG H, LIU G M, et al. Spatio-temporal variability of chlorophylland its responses to sea surface temperature, winds and height anomaly in the western South China Sea[J]. Acta Oceanologica Sinica, 2013, 32(1): 48-58.
11 王肖颖, 张芳, 李娟英, 等. 2012年夏季挪威海和格陵兰海浮游植物群落结构的色素表征[J]. 生态学报, 2014, 34(8): 2076-2084.
12 MURRAY C, MARKAGER S, STEDMON C A, et al. The influence of glacial melt water on bio-optical properties in two contrasting Greenlandic fjords[J]. Estuarine, Coastal and Shelf Science, 2015, 163: 72-83.
13 庄燕培, 金海燕, 陈建芳, 等. 北冰洋中心区表层海水营养盐及浮游植物群落对快速融冰的响应[J]. 极地研究, 2012, 24(2): 151-158.
14 辜帆, 金海燕, 庄燕培, 等. 2012年夏季水团输送对挪威-格陵兰海营养盐及浮游植物群落结构的影响[J]. 极地研究, 2018, 30(1): 22-31.
15 魏皓, 赵伟, 罗晓凡, 等. 北冰洋浮游生物空间分布及其季节变化的模拟[J]. 海洋学报, 2019, 41(9): 65-79.
16 JUUL-PEDERSEN T, ARENDT K E, MORTENSEN J, et al. Seasonal and interannual phytoplankton production in a sub-Arctic tidewater outlet glacier fjord, SW Greenland[J]. Marine Ecology Progress Series, 2015, 524: 27-38.
17 MEIRE L, MORTENSEN J, RYSGAARD S, et al. Spring bloom dynamics in a subarctic fjord influenced by tidewater outlet glaciers (Godthåbsfjord, SW Greenland)[J]. Journal of Geophysical Research: Biogeosciences, 2016, 121(6): 1581-1592.
18 KRAWCZYK D W, WITKOWSKI A, JUUL-PEDERSEN T, et al. Microplankton succession in a SW Greenland tidewater glacial fjord influenced by coastal inflows and run-off from the Greenland Ice Sheet[J]. Polar Biology, 2015, 38(9): 1515-1533.
19 蒋兴伟, 林明森, 张有广, 等. 海洋遥感卫星及应用发展历程与趋势展望[J]. 卫星应用, 2018(5): 10-18.
20 PEARCE D M, MAIR D W F, REA B R, et al. The glacial geomorphology of upper Godthåbsfjord (Nuup Kangerlua) in southwest Greenland[J]. Journal of Maps, 2018, 14(2): 45-55.
21 GRØNKJÆR P, NIELSEN K V, ZOCCARATO G, et al. Feeding ecology of capelin (Mallotus villosus) in a fjord impacted by glacial meltwater (Godthåbsfjord, Greenland)[J]. Polar Biology, 2019, 42(1): 81-98.
22 MORTENSEN J, BENDTSEN J, LENNERT K, et al. Seasonal variability of the circulation system in a west Greenland tidewater outlet glacier fjord, Godthåbsfjord (64°N)[J]. Journal of Geophysical Research-Earth Surface, 2014, 119(12): 2591-2603.
23 MASCARENHAS V J, ZIELINSKI O. Hydrography-driven optical domains in the Vaigat-Disko Bay and Godthabsfjord: Effects of glacial meltwater discharge[J]. Frontiers in Marine Science, 2019, 6. DOI: 10.3389/fmars. 2019. 00335.
24 ZIELINSKI O, BRAUN A, HENKEL R, et al. Physical oceanography during Maria S. Merian cruise MSM65 (GREENHABⅡ) [EB/OL]. [2021-10-05]. https://doi.org/10.1594/PANGAEA.886181.
25 GOOD S, FIEDLER E, MAO C Y, et al. The Current configuration of the OSTIA system for operational production of foundation sea surface temperature and ice concentration analyses[J]. Remote Sensing, 2020, 12(4): 720. DOI: 10.3390/rs12040720.
26 STARK J D, DONLON C J, MARTIN M J, et al. OSTIA : An operational, high resolution, real time, global sea surface temperature analysis system[J]. OCEANS 2007-Europe, 2007: 1-4. DOI: 10.1109/OCEANSE. 2007. 4302251.
27 DONLON C J, MARTIN M, STARK J, et al. The Operational Sea Surface Temperature and Sea Ice Analysis (OSTIA) system[J]. Remote Sensing of Environment, 2012, 116: 140-158.
28 KRAVITZ J, MATTHEWS M, BERNARD S, et al. Application of Sentinel 3 OLCI for chl-a retrieval over small inland water targets: Successes and challenges[J]. Remote Sensing of Environment, 2020, 237: 111562.
29 DOERFFER R, SCHILLER H. The MERIS case 2 water algorithm[J]. International Journal of Remote Sensing, 2007, 28(3/4): 517-535.
30 BROCKMANN C , DOERFFER R , PETERS M , et al. Evolution of the C2RCC neural network for Sentinel 2 and 3 for the retrieval of ocean colour products in normal and extreme optically complex waters[J]. ESASP, 2016, 740-745.
31 VANHELLEMONT Q, RUDDICK K. Atmospheric correction of Sentinel-3/OLCI data for mapping of suspended particulate matter and chlorophyll-a concentration in Belgian turbid coastal waters[J]. Remote Sensing of Environment, 2021, 256: 112284.
32 MORTENSEN J, LENNERT K, BENDTSEN J, et al. Heat sources for glacial melt in a sub-Arctic fjord (Godthåbsfjord) in contact with the Greenland Ice Sheet[J]. Journal of Geophysical Research-Oceans, 2011, 116(C1): C01013.
Temporal and spatial variation of Chlorophyllconcentration in Nuup Kangerlua Fjord, Greenland
Tian Jiahui1, Cheng Xiao1,2,3,4, Zhang Yannan2,4
(1State Key Laboratory of Remote Sensing Science, College of Global Change and Earth System Science, Beijing Normal University, Beijing 100875, China;2School of Geospatial Engineering and Science, Sun Yat-sen University, Zhuhai 519082, China;3Southern Marine Science and Engineering Guangdong Laboratory (Zhuhai), Zhuhai 519082, China;4University Corporation for Polar Research, Beijing 100875, China)
The temporal and spatial distribution of Chlorophyll(Chl) concentration in Nuup Kangerlua Fjord is an indicator of climate change. The spatiotemporal distribution of Chlconcentration in this region from 2016 to 2020 was therefore analyzed by combining the Chldata obtained from Sentinel-3 OLCI inversion by C2RCC algorithm, Operational Sea Surface Temperature and Ice Analysis (OSTIA) sea surface temperature (SST) data, and data from a nearby meteorological station (at Nuuk). The results show that the distribution of Chlwas mainly affected by wind speed, wind direction and SST at the fjord inlet. The peak time of Chlconcentration was about one month later than that of SST and is most widespread when the winds are at their strongest in September. At the outlet of the glacier, in addition to being influenced by wind and water temperature, the spring bloom of Chlis followed by a second bloom in autumn, driven by meltwater from glaciers.
glacial fjords, Chlorophyll, Sentinel-3 OLCI, C2RCC
2021年11月收到来稿, 2022年3月收到修改稿
国家自然科学基金(41825002)资助
田佳慧, 女, 1997年生。硕士研究生, 主要从事极地水色遥感研究。E-mail: tianjiahui@mail.bnu.edu.cn
程晓, E-mail: chengxiao9@mail.sysu.edu.cn
10.13679/j.jdyj.20210086
