In vitro and in silico studies of salicylic acid on systemic induced resistance against bacterial leaf blight disease and enhancement of crop yield
2023-01-06WannapornTHEPBANDITNarendraKumarPAPATHOTIJayasimhaRayuluDADDAMNguyenHuyHOANGToanLETHANHChanonSAENGCHANKumraiBUENSANTEAI
Wannaporn THEPBANDIT ,Narendra Kumar PAPATHOTI ,Jayasimha Rayulu DADDAM ,Nguyen Huy HOANG ,Toan LE THANH ,Chanon SAENGCHAN ,Kumrai BUENSANTEAI
1 School of Crop Production Technology,Suranaree University of Technology,Nakhon Ratchasima 30000,Thailand
2 Department of Animal Science,Agriculture Research Organization,Rishon Lezion 7505101,Israel
3 Department of Plant Protection,Can Tho University,Can Tho 900000,Vietnam
Abstract Salicylic acid (SA) is an effective elicitor to promote plant defenses and growth.This study aimed to investigate rice(Oryza sativa L.) cv.Khao Dawk Mali 105 treated with salicylic acid (SA)-Ricemate as an enhanced plant protection mechanism against bacterial leaf blight (BLB) disease caused by Xanthomonas oryzae pv.oryzae (Xoo).Results indicated that the use of SA-Ricemate as a foliar spray at concentrations of more than 100 mg L–1 can reduce the severity of BLB disease by 71%.SA-Ricemate treatment also increased the hydrogen peroxide (H2O2) content of rice leaf tissues over untreated samples by 39–61%.Malondialdehyde (MDA) in rice leaves treated with SA-Ricemate also showed an increase of 50–65% when comparing to non-treated samples.The differential development of these defense compounds was faster and distinct when the SA-Ricemate-treated rice was infected with Xoo,indicating plant-induced resistance.Besides,SA-Ricemate elicitor at a concentration of 50–250 mg L–1 was correlated with a substantial increase in the accumulation of total chlorophyll content at 2.53–2.73 mg g–1 of fresh weight which suggests that plant growth is activated by SA-Ricemate.The catalase-and aldehyde dehydrogenase-binding sites were searched for using the CASTp server,and the findings were compared to the template.Chemsketch was used to design and optimize SA,which was then docked to the catalase and aldehyde dehydrogenase-binding domains of the enzymes using the GOLD 3.0.1 Software.SA is shown in several docked conformations with the enzymes catalase and aldehyde dehydrogenase.All three catalase amino acids (GLN7,VAL27,and GLU38) were discovered to be involved in the creation of a strong hydrogen bond with SA when SA was present.In this mechanism,the aldehyde dehydrogenase amino acids LYS5,HIS6,and ASP2 were all implicated,and these amino acids created strong hydrogen bonds with SA.In field conditions,SA-Ricemate significantly reduced disease severity by 78% and the total grain yield was significantly increased which was an increase of plant height,tiller per hill,and panicle in three field trials during Aug–Nov 2017 and 2018.Therefore,SA-Ricemate can be used as an alternative elicitor on replacing harmful pesticides to control BLB disease with a high potential of increasing rice defenses,growth,and yield components.
Keywords: bacterial leaf blight,crop yield,in vitro,in silico,rice defense,salicylic acid
1.Introduction
Rice (Oryza sativaL.) is one of the largest cultivated crops in the world.Rice production can be negatively affected by a variety of factors such as poor soil quality,nutrient deficiency,pests,and diseases which lead to a reduction in rice yield (Timsinaet al.2018).One of the most important diseases in the rice production system is the bacterial leaf blight (BLB) caused byXanthomonas oryzaepv.oryzae(Xoo).The BLB widely presents in South-East Asia and Japan causing average losses of 20–50% and up to 100% in severe conditions (Walterset al.2013).The BLB is considered to be one of the most dangerous rice diseases in Thailand due to favorable climate conditions (Kawasaki and Herath 2011).Reducing the intensity of BLB is a key priority to improve Thai rice production.The main existing methods used to control the BLB are focused on synthetic chemicals such as copper hydroxide,copper oxychloride,copper sulfate.However,these components potentially have adverse effects on humans and the environment.Therefore,it is important to perform research on effective green and healthy agrochemicals to control the BLB.Advanced control methods based on biotic and abiotic elicitors to activate plant defense mechanisms have been reported with plant growth-promoting rhizobacteria (PGPR),oligosaccharides,polymers of carbohydrate,derivatives of lipids,green agrochemicals,and chemical plant hormones (Aranega-Bouet al.2014).In recent years,abiotic elicitors have been extensively investigated as a means of controlling plant diseases based on the induced resistance (IR) principle,including induced systemic resistance (ISR),and systemic acquired resistance(SAR) (Thakur and Sohal 2013).IR contributes to the production of innate defenses against plant pathogens involving multiple biochemical pathways that can be activated by plant elicitors/activators/inducers (Thakur and Sohal 2013).A significant feature of IR is planting priming such as a foliar application or seed priming which enables the rapid expression of defense responses to pathogen infections (Le Thanhet al.2017).Salicylic acid(SA) is a phenolic compound that works by signaling for the acquisition of systemic resistance as a plant defense against several pathogens (Dempsey and Klessig 2017).In addition,SA plays an important role as a plant hormone in the growth and development of plants and as a plant promoter of innate immunity (Hayatet al.2013).The SA response in immune plants is associated with the oxidative burst and the induction of the reactive oxygen species (ROS).The oxidative burst response to SA has been recorded in a different number of plants such as potato,rice,tobacco due to the increased hydrogen peroxide (H2O2) production (Kauss and Jeblick 1995)which is close to the effects of the chitooligosaccharides.The increased levels of H2O2have led to the expression of glucanase and chitinase transcripts in rice which lead to the induction of plant resistance.It was suggested that H2O2development under IR resulted in functional formation and reinforcement of plant structural tissues(Leonet al.1995).Malondialdehyde (MDA) is a marker of oxidative lipid injury associated with programmed cell death (PCD) with varying concentrations in response to biotic and abiotic stresses (Zoelleret al.2012).The effects of SA treatments on H2O2generation,lipid peroxidation,and oxidative damage to proteins,as well as the formation of chlorophyll and carotene isomers,were all seen in the experiments.H2O2levels were shown to rise in response to SA,which was associated with enhanced activity of Cu-,Zn-superoxide dismutase but not with changes in catalase or ascorbate peroxidase activities.In addition,the use of SA induces retrograde signals derived from chloroplasts such as chlorophyll synthesis,encoded photosynthesis genes,and essential mediators of plant immunity (Nomuraet al.2012).The application of SA was investigated as a resistance elicitor against plant pathogen in greenhouse conditions based on the IR principle,whereas SA has never been evaluated on rice fields neither developed into a commercial formulation in Thailand.
Therefore,this work is focused on investigating the resistance mechanism caused by abiotic SARicemate elicitor formulation based on SA combined with inert ingredients for stimulating H2O2,MDA,and total chlorophyll content.This SA-Ricemate is also investigated for the reduction of BLB severity as well as growth parameters and rice yield of three consecutive crops,during Aug–Nov 2017 and 2018.
2.Materials and methods
2.1.Source of elicitor
The exogenous SA elicitor (SA-Ricemate 50,100,150,200,250,and 300 mg L–1concentrations used in this study) prototype is a product of the Bioactive Agro Industry Co.,Ltd.,Thailand;developed at the Plant Pathology &Biopesticide Laboratory,Suranaree University of Technology,Nakhon Ratchasima,Thailand.
2.2.Rice cultivar,bacterial strains,and culture conditions
A homogenous sample of Thai jasmine rice (O. sativaL.)seeds of susceptible BLB cultivar Khao Dawk Mali 105(KDML 105) was obtained from the Rice Seed Center in Thailand.TheXooSUT1-121 strain of virulence was used in this research and provided by the Prae Rice Research Institute,Thailand.The bacterial strain was grown in the nutrient glucose broth (NGB) at (28±1)°C followed by a continuous shaking of 180 r min–1for 48 h(Buensanteaiet al.2008;Krishnanet al.2014).
2.3.The assessment of SA-Ricemate against BLB under greenhouse conditions
TheXooSUT1-121 strain was inoculated on rice plants by the scissors dip technique.In this method,Xoo-soaked scissor tips cut the leaf tip (about 2–3 cm for seedling plants and 4–5 cm for older plants) away from the leaf.Growing temperatures ranged from 28–32°C (light,12 h) to 28–32°C(dark,12 h) with a relative humidity of 90 throughout the growing period (Keet al.2017).Rice seedscv.KDML 105 were soaked in water for 24 h before planting.The seedlings were transferred to 30-cm diameter plant pots containing 5 kg soil from farm of Suranaree University of Technology.The pots were held in greenhouse conditions under 12 h of photoperiod and 60–75% of moisture content.The rice plants were sprayed with 30 mL of the SA-Ricemate at 15,30,and 45 days after sowing,using different concentrations including 50,100,150,200,250,and 300 mg L–1;two positive control treatments were also used as copper hydroxide and commercial elicitor (COS®)4 000 mg L–1;the active ingredient of chitooligosaccharide.Finally,mock water was used as a negative control for the untreated samples.The suspension ofXooat 1×108CFU mL–1was inoculated on the upper-leaf by cutting the tip of the rice leaf;then,the plants were covered with plastic bags and incubated at 28°C for 24 h(Chithrashreeet al.2011;Le Thanhet al.2017).The severity of BLB disease was measured at 7,14 and 21 days of inoculation using the International Rice Research Institute (IRRI) method (IRRI 2017).The percentage of disease severity was calculated based on the following eq.(1).The reduction in disease severity was calculated using the eq.(2) (Ferreiraet al.2017;Le Thanhet al.2017).
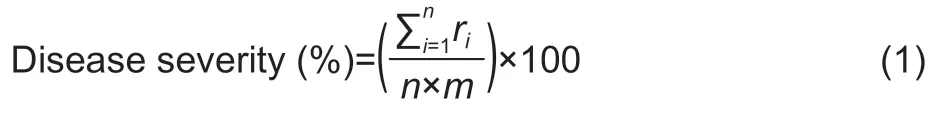
whereris the set of numerical ratings,nis the total of evaluations per sample,andmis the maximum value used for the evaluations.

where DSn is the calculated disease severity of untreated samples and DSt is the calculated disease severity of the elicitor-treated samples.
2.4.Determination of hydrogen peroxide (H2O2)
A total of 0.5 g of rice leaf tissues from each treatment was soaked in liquid nitrogen and homogenized with 5 mL of 0.1% trichloroacetic acid and then centrifuged at 14 000×g for 15 min under 4°C.The supernatant dosage at 0.5 mL was combined with 0.5 mL of 10 mmol L–1potassium phosphate buffer,pH 7.0,and 1 mL of 1 mol L–1KI.The absorbance was quantified at 390 nm by a microplate reader (Bio-Tek,USA).The content of H2O2was calculated based on the standard curve (Sahebani and Hadavi 2009).
2.5.Determination of MDA
A total of 0.5 g of rice leaf tissues of each treatment was soaked in liquid nitrogen and homogenized with 5 mL of 5% trichloroacetic acid,then centrifuged at 4 000×g for 10 min.A 2-mL dose of the supernatant was combined with 2 mL of 0.67% barbituric acid.Then,the mixture was boiled for 30 min at 100°C and cooled down in the ice.Next,the mixture was centrifuged at 3 000×g for 10 min.The absorption at 532,600,and 450 nm was quantified by a microplate reader (Bio-Tek,USA).The content of the MDA was calculated based on the following formula provided in Draper and Hadley’s research (Draper and Hadley 1990).
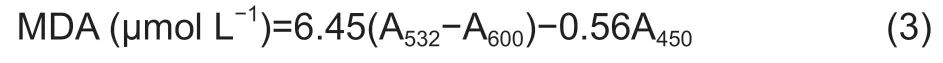
where A450,A532,and A600are the absorbance values at the wavelengths 450,532,and 600 nm,respectively.
2.6.In silico inhibitory studies of SA on H2O2 degrading enzyme catalase and MDA degrading enzyme aldehyde dehydrogenase
Complete molecular simulations have been carried out on AMD 64 bits dual processing hi-end Linux desktop.From the protein databank,the structure of catalase and aldehyde dehydrogenase fromO. sativa(Uniprot_KB accession ID: Q10S82 and Q84LK3) were acquired through lack of availability of 3D structure in the database,predicted models were collected in PDB format.For domain selection,the catalase and aldehyde dehydrogenase fromO. sativawere submitted to SBASE(pongor.itk.ppke.hu) server.Later,by molecular dynamics simulation studies,the protein was stabilized by adding hydrogens to the three-dimensional structure.With the help of the NAMD 2.8 and CHARMM27 force field,molecular dynamics (MD) simulations of the predicted model were performed.A multiple-time-stepping is the algorithm used where at every two steps,the long-range electrostatics was calculated,and at every level,shortrange forces were calculated.With the help of Hamilton’s equations of motion,the MD procedure is carried out in this study at the new positions to receive new velocities.A final model with advanced thermodynamic properties information is obtained,and in terms of RMSD,it is stabilized (Kurjogiet al.2018;Daddamet al.2020a).
In molecular dynamics studies,the structures of catalase and aldehyde dehydrogenase with lesser root mean square deviation (RMSD) is achieved,and to examine the stereochemical quality of protein structures;it is then figured out by Ramachandran plot,using PROCHECK server.Later,the environment profile is checked using the structure evaluation server ERRAT.To recognize the active site as well as to under docking to the SA;this protein is then can be utilized (Daddamet al.2020a;Harathiet al.2021).
2.7.Active site identification of catalase and aldehyde dehydrogenase
Depending upon the template’s structural comparison as well as the required model built,with the help of the CASTp server (http://www.sts.bioe.uic.edu/castp),the possible binding sites of catalase and aldehyde dehydrogenase fromO. sativawas checked after that the ultimate model was built.Using the SPDBV Program based on the template’s structure–structure comparison,the final refined model of the laccase domain was developed (Papathotiet al.2016;Shaiket al.2021).
2.8.Docking studies with SA
Using GOLD 3.0.1 Software,insight into the binding conformation of SA was gained by performing docking studies (Thepbanditet al.2021).In this method,the SA compound was docked to the active site of catalase and aldehyde dehydrogenase to study the possibility of inhibitory activity of the SA.After docking,the individual binding poses of each protein–protein complex were selected and their binding energies were studied.The most energetical conformation of the complex was selected and docking studies were analyzed (Daddamet al.2014).
2.9.Determination of chlorophyll content
Firstly,0.1 g of rice leaf tissues was homogenized with 5 mL of acetone and ethanol (4:1).The extraction was performed under dark conditions for 3 h and then centrifuged at 4 000×g for 5 min.The supernatant was quantified at 645 and 663 nm by the microplate reader(Bio-Tek,USA).The chlorophyll content was calculated based on the following formulas proposed by Aminot and Rey (2002).

where A645and A645are the absorbance values at the wavelengths 645 and 663 nm,respectively.
2.10.Field arrangement,cultivation,and the spray of elicitors
This experiment was based on the application of Randomized Complete Block Design (RCBD) with four treatments including SA-Ricemate,commercial elicitor of chitooligosaccharide;COS®,copper hydroxide 77% WP,and water as untreated control with four replications and three consecutive crops during Aug–Nov 2017 and 2018.All experiments were performed in the paddy field in Khok-Sung,Ubolratana District,Khon-Kaen Province,Thailand(DMS: 16°40´38.96´´N,102°41´57.23´´E).Treatments and replications were divided by 1 m of buffer strips.The individual plot size was 81 m2(9 m×9 m).The planting area was configured in 25 cm×25 cm row by row with 32 rows and 32 seedlings per row in each plot (Rice and Institute 1996).The margin rows were considered as boundary rows to prevent potential contamination of the samples.The magnitude of BLB disease growth parameters and yield production was assessed by quadrants(1 m2) as representative data for each treatment plot (Rice and Institute 1996).
The seedlings were prepared for transplantation as a seedbed in a small plot.The seedbed was prepared with farm soil and decomposed by 0.5 kg m–2,10 cm in deep.The rice seedscv.KDML 105 were soaked for 24 h and sown in seedbeds at 50 g m–2.The 20-days old seedlings were uprooted and arranged for replanting at 3 stems per hill.The field trial was plowed crisscross twice by using a tractor 7 days before transplantation.Decomposition at 500 kg rai–1(1 rai=0.4 acres) was applied 3 days before transplantation.The fertilizers used in the experiments were based on nitrogen,phosphorus,and potassium following the recommendations from the Land Department of Development (LDD),Ministry of Agriculture and Cooperatives,Thailand.The 16–20–0 fertilizer formula was added to the field for 10 days after transplantation(tilling stage) at a dose of 30 kg rai–1and 0–0–60 fertilizer formula at a dose of 10 kg rai–1.The 46–0–0 fertilizer formula was used for 50 days after transplantation (panicle initiation stage) at 3 kg rai–1in crop 1 (Aug–Nov 2017) and crop 2 (Aug–Nov 2018).A double dose of fertilizer like 16–20–0,0–0–60,and 46–0–0 was added to crop 3 (Aug–Nov 2018) to contribute to plant susceptibility to disease.Hand weeding was performed to suppress weeds.In addition,golden apple snails were removed by hand.
The elicitor was applied by spraying rice foliage(20 L rai–1) three times at 15,30,and 45 days posttransplantation.Water was used as a negative control treatment (untreated) while the commercial elicitor of chitooligosaccharide;COS®and copper hydroxide were used as two positive controls.
2.11.Disease severity,growth parameters,and BLB severity
The capability for management of the BLB was assessed against naturalXooinfection.The severity of the disease was assessed four times from 45 days posttransplantation.Percentage of disease severity and reduction was determined by using the eqs.(1) and (2)(Ferreiraet al.2017;Le Thanhet al.2017).
The plant parameters were observed in terms of plant height,number of tillers per hill,number of seeds per panicle,1 000-grain weight,and overall grain yield.Grain yield was calculated and reported as kg rai–1according to the eq.(7) provided in the research by Regalado and Ramos (2018).
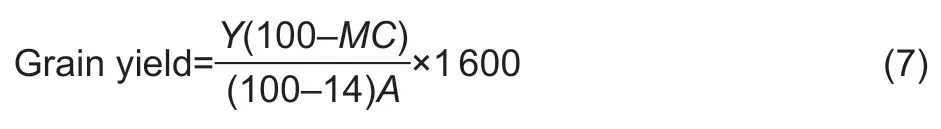
whereMCis the grain moisture content (%),Yis the net yield (kg) andAis the plot area (rai).
2.12.Statistical analysis
To demonstrate data consistency,the experiments under greenhouse conditions were repeated three times,and experiments underfield conditions were repeated two consecutive years.The experimental data were analyzed by one-way ANOVA using SPSS Software version 20.The mean differences were divided by Duncan’s multiple range test with a significance ofP=0.05.
3.Results
3.1.The efficacy of SA-Ricemate against BLB under greenhouse conditions
The use of SA-Ricemate at 100 mg L–1or higher concentrations substantially decreases the incidence of BLB disease when used as a foliage treatment at 15,30,and 45 days after sowing.BLB disease reduction of the rice plants that were treated with 100 mg L–1was 42.86,57.14,and 50.00% at 7,14,and 21 days after inoculation,respectively.Nevertheless,the protective efficacy of SARicemate at 150,200,250,and 300 mg L–1indicates a non-significant reduction in the BLB disease severity when compared with the 100 mg L–1treatment (Table 1).
3.2.The change of H2O2
The presence of H2O2in rice plants was increased at 24 h post-inoculation for all treatments.The H2O2contents of rice plants treated with SA-Ricemate were increased by 100,150,200,250,and 300 mg L–1and commercial elicitor;COS®with 39.31,46.48,44.25,54.36,61.10,and 62.84%,respectively.The copper hydroxide treatment had an improvement of 12.68% in H2O2relative to 9.63%in the untreated control (Table 2).
3.3.The change of MDA
Treated plants experienced a rise in the MDA content after 24 h from inoculation.Rice plants treated with commercial elicitor COS®and SA-Ricemate at 100,150,200,250,and 300 mg L–1showed a significantly higher increase of MDA with 69.69,60.98,54.59,50.58,65.04,and 53.06%,respectively,compared to untreated samples(33.01%).However,the MDA increase in rice treated with SA-Ricemate at 50 mg L–1showed an accumulation of 40.23% of MDA which was considered non-significant compared to untreated rice.Samples treated with copper hydroxide showed a significantly lower MDA of 23.75%(Table 3).
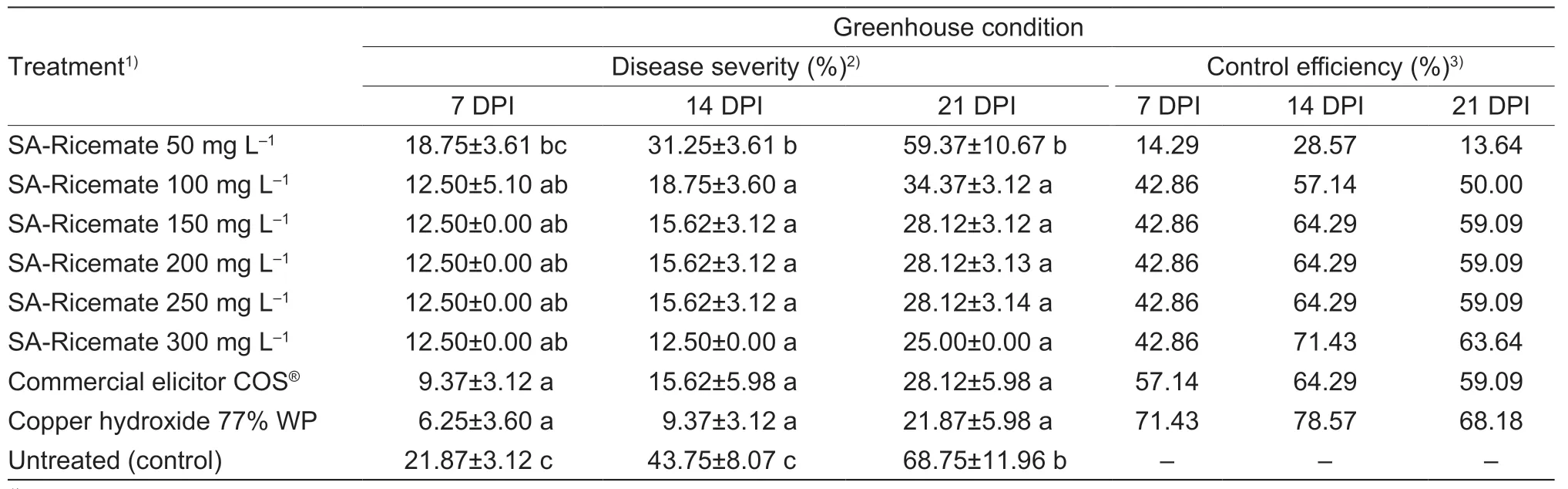
Table 1 Effectiveness of salicylic acid (SA)-Ricemate on the disease severity of bacterial leaf blight (BLB) in rice cv.KDML 105 caused by Xanthomonas oryzae pv.oryzae (Xoo) strain SUT1-121
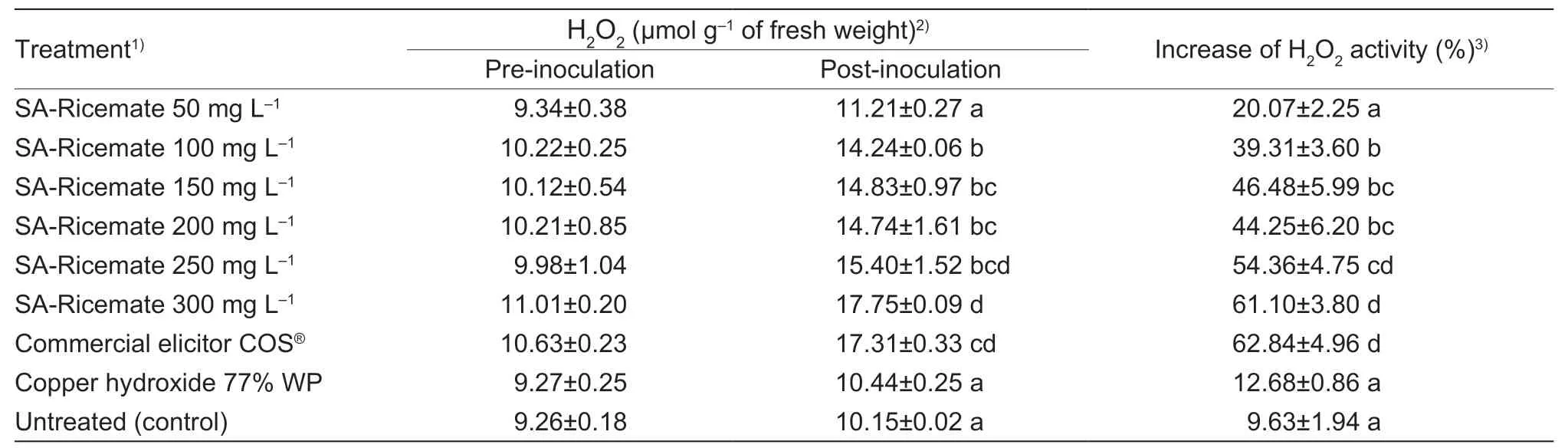
Table 2 Effectiveness of salicylic acid (SA)-Ricemate on the changes of hydrogen peroxide (H2O2) content in rice cv.KDML 105 infected by Xanthomonas oryzae pv.oryzae (Xoo) strain SUT1-121
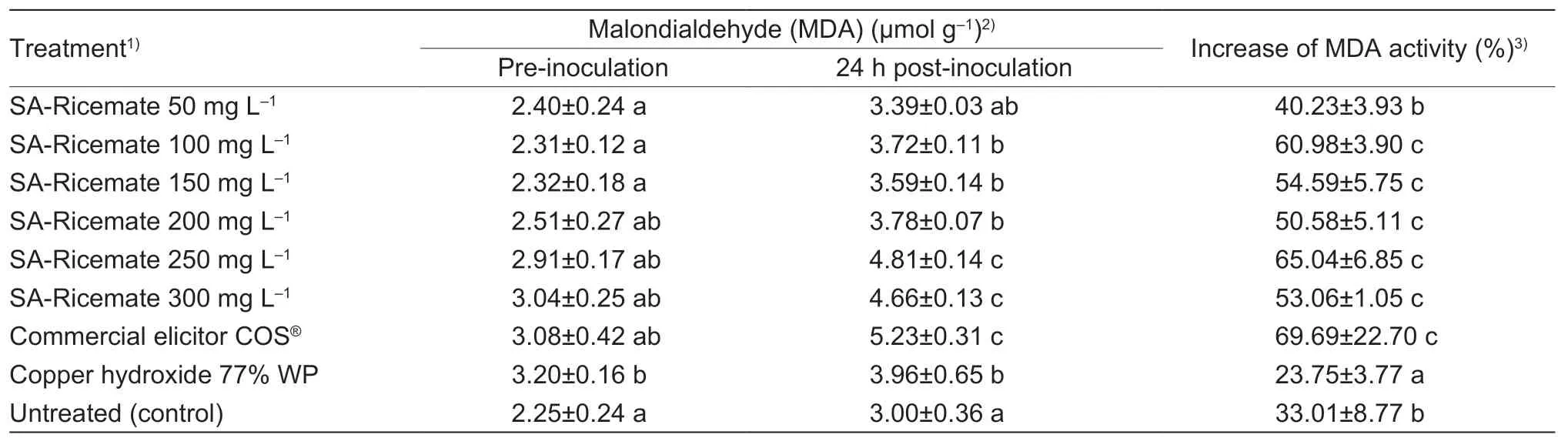
Table 3 Effectiveness of salicylic acid (SA)-Ricemate based on the changes of malondialdehyde (MDA) content in rice cv.KDML 105 infected by Xanthomonas oryzae pv.oryzae (Xoo) strain SUT1-121
3.4.Inhibitory activity of salicylic acid on catalase and aldehyde dehydrogenase by docking studies
In order to find the possible inhibitory activity of SA on catalase and aldehyde dehydrogenase fromO. sativa,a three-dimensional structure was obtained from proteindatabank and by molecular dynamics and validated using Ramachandranplot server using PROCHECK Program(Figs.1-A and 2-A).The binding sites of catalase and aldehyde dehydrogenase were searched using the CASTp server as well as comparing with the template(Figs.1-B and 2-B).The SA was designed and optimized by Chemsketch Software and docked to catalase and aldehyde dehydrogenase binding region using GOLD 3.0.1 Software.The selected docked conformations of SA with the catalase and aldehyde dehydrogenase binding site are depicted in Figs.1-C and 2-C.The catalase amino acids GLN7,VAL27 and GLU38 were involved in forming a strong hydrogen bond with SA (Fig.1-D).The aldehyde dehydrogenase amino acids LYS5,HIS6 and ASP2 were involved in forming a strong hydrogen bond with SA (Fig.2-D).Based on this,it is suggested that the inhibition of catalase and aldehyde dehydrogenase was due to binding of SA at active site and forming a complex with the active residues of catalase and aldehyde dehydrogenase.

Fig.1 Inhibitory studies of salicylic acid (SA) with catalase.A,structure of catalase with helices (red color) and sheets (blue color).B,active site of catalase in red color pocket in the structure.C,docking of S A (green color) with catalase.D,amino acids of catalase involved in docking with SA.
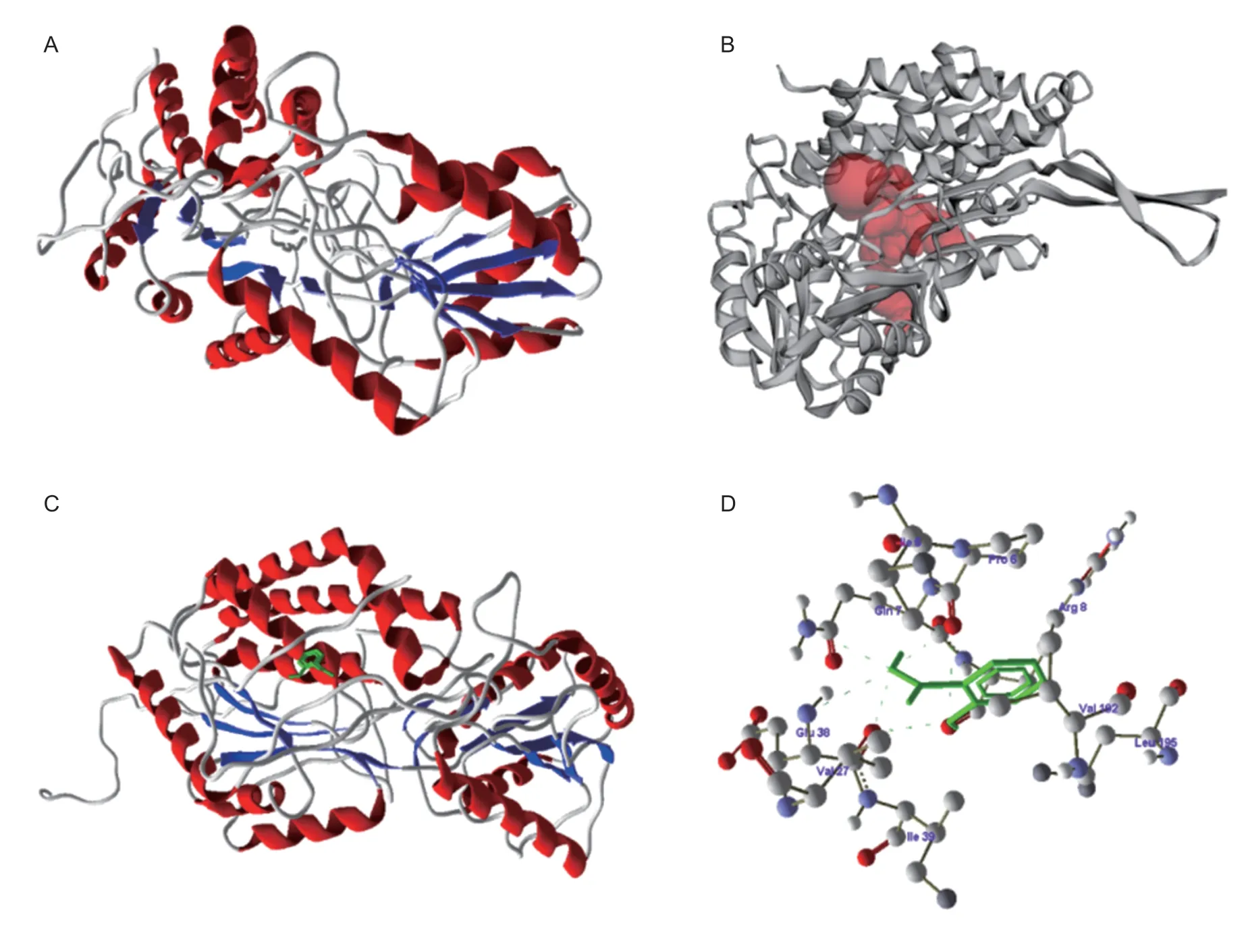
Fig.2 Inhibitory studies of salicylic acid (SA) with aldehyde dehydrogenase.A,structure of aldehyde dehydrogenase with helices(red color) and sheets (blue color).B,active site of aldehyde dehydrogenase in red color pocket in the structure.C,docking of SA (green color) with aldehyde dehydrogenase.D,amino acids of aldehyde dehydrogenase involved in docking with SA.
3.5.The change of chlorophyll content
The total chlorophyll content in rice leaves treated with SA-Ricemate increased significantly by 50,100,150,200,and 250 mg L–1,and commercial elicitor COS®chlorophyll content 2.65,2.66,2.73,2.62,2.53,and 2.29 mg g–1of fresh weight,respectively.The treatment with copper hydroxide and the treatment with 300 mg L–1of SARicemate were shown to be non-significant compared to the untreated (Table 4).
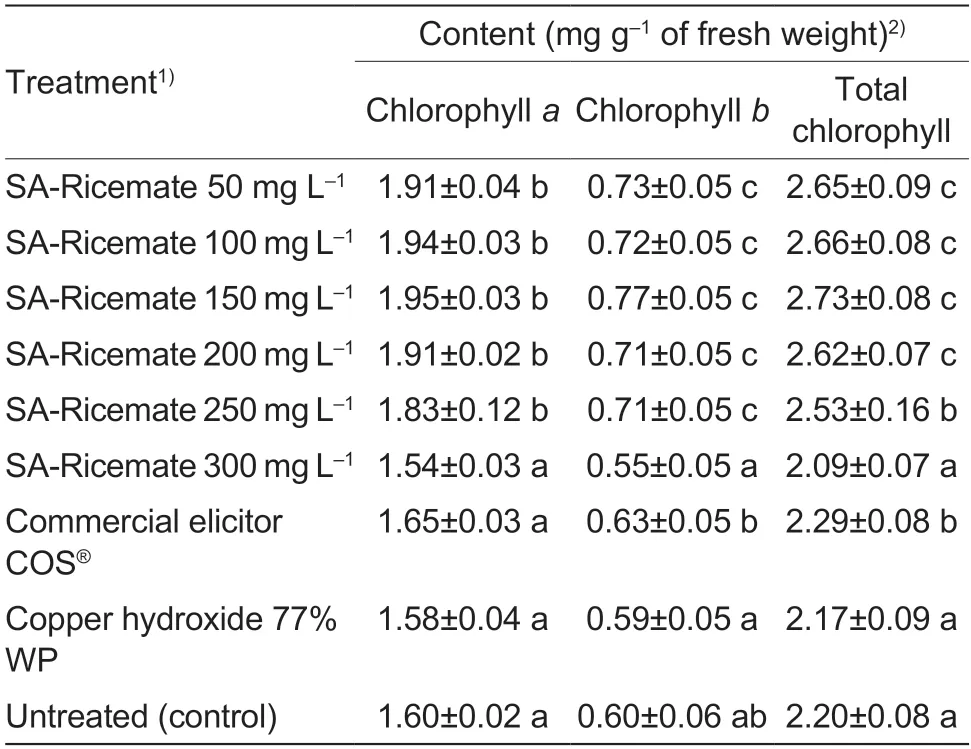
Table 4 Effectiveness of salicylic acid (SA)-Ricemate based on the accumulation of chlorophyll content in rice cv.KDML 105 infected by Xanthomonas oryzae pv.oryzae (Xoo) strain SUT1-121
3.6.The efficacy of SA-Ricemate against BLB under field conditions
The BLB reduction in SA-Ricemate treated paddy field was significantly higher compared with a negative control in all three crops which was reduced by 66.19% in crop 1,45.57% in crop 2,and 44.27% in crop 3.SA-Ricemate results were shown non-significantly different from the two positive controls.The commercial elicitor COS®showed BLB reductions of 66.92,51.72,and 40.03%,respectively.The copper hydroxide treatment showed a BLB reduction of 77.11,54.14,and 45.12%,respectively (Fig.3-A–C).
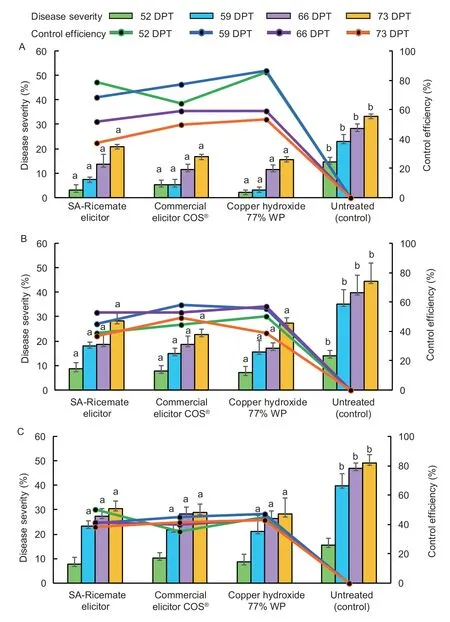
Fig.3 Efficacy of the testing elicitors on disease severity of bacterial leaf blight (BLB) disease on rice KDML 105 in the first crop season,during Aug–Nov 2017 (A),in the second crop season at location 1 during Aug–Nov 2018 (B),and in the second crop season at location 2 during Aug–Nov 2018 (C) at 52,59,66,and 73 days post-transplant (DPT).Data are mean±SD (n=5).Different letters (a,b) indicated significant difference via Duncan’s multiple range test at P=0.05.
3.7.The efficacy of SA-Ricemate in yield components and yield
Yield components,plant height,tillers per hill,panicles per hill,grains per panicle,1 000-grain weight,and yield were significantly increased by the SA-Ricemate treatment in all 3 crops during Aug–Nov 2017 and Aug–Nov 2018 for KDML 105 rice as shown in Fig.4-A–E.The data showed that the rice plant samples treated with SA-Ricemate were significantly different in plant height with 108 and 116 cm height at 45 and 75 days post-transplant (DPT) in crop 1;113,120,and 143 cm at 45 and 75 DPT in crop 2;123 and 145 cm at 75 and 105 DPT in crop 3.The highest number of tillers per hill was obtained from rice plants treated with SA-Ricemate with 14.4 tillers per hill in crop 1.In crop 2 and crop 3,the highest number of tillers per hill was obtained from rice plants treated with commercial elicitor COS®with 16.0 and 17.8 tillers per hill which were not significantly different compared to SA-Ricemate treated samples.The highest number of panicles per hill was obtained from rice plant treated with SA-Ricemate with 14.0,and 12.4 panicles per hill in crop 2 and crop 3,respectively;whereas the highest number of panicles per hill was obtained from rice plant treated with copper hydroxide in crop 1 with 12.8 panicle per hill which was not significantly different when compared to SA-Ricemate treated samples.Grain per panicle was significantly higher in SA-Ricemate treated samples in three crops with 142 grains per panicle when compared to the negative control.The 1 000-grain weight was significantly increased by the SA-Ricemate treatment in all three crops to 27.38 g.The highest 1 000-grain weight measurement was obtained from rice plants treated with commercial elicitor COS®in crop 1 with 28.44 g and crop 3 with 28.7 g,which was not significantly different compared to SARicemate treated samples.

Fig.4 Efficacy of the testing elicitors on rice KDML 105 growth parameters and yield component.DPT,days post-transplant.Data are mean±SD (n=5).Different letters (a,b,c) indicated significant difference via Duncan’s multiple range test at P=0.05.ns,non-significant.
4.Discussion
Rice plants treated with SA-Ricemate by foliar application as described in this study showed a significant reduction in the BLB disease severity of >50% in greenhouse conditions and >30% in field trials.Data obtained from multiple experiments such as H2O2,MDA,chlorophyll,growth parameters,and yield;reported that SA-Ricemate plays an important role in the protection of productivity as phytohormones in rice plants.Previously,the evidence of SA in plant protection has been appreciated in several research works (Waret al.2011;López-Gresaet al.2016;Le Thanhet al.2017).The elicitor is applied by foliage priming which induces host plants to participate in a process of protection against pathogen infection by triggering bio-signals that causes systemic acquired resistance (SAR) based on the IR principle (Penget al.2015;Gaoet al.2017;Tripathiet al.2019) studies has shown that the elicitor cause plant immunity by increasing resistance to fungal,bacterial,and viral pathogens in several plants such as tomato,rice,wheat (Hussainet al.2015;Adsset al.2017;Le Thanhet al.2017;Uedaet al.2019).The concentration of SA-Ricemate determines the efficacy of the disease control treatment having observed that,in greenhouse conditions,doses from 100 to 300 mg L–1are in the ideal concentration range for BLB disease control.The dose of SA-Ricemate elicitor is an important metric for stimulating plant defense enzymes.The results obtained are consistent with Ghazanfaret al.(2019) who found that 6 mmol L–1of SA produced promising antifungal activity by inhibiting mycelium growth in tomato diseases as well as sour rot,pink mold rot,and rhizopus soft rot.However,doses of 2 and 4 mmol L–1have not been considered to be effective for disease control(Ghazanfaret al.2019).Waret al.(2011) reported that chickpea (Cicer arietinumL.) reacted to SA treatment at 1.5 mmol L–1with higher induction of plant defense enzymes as peroxidase,polyphenol oxidase,H2O2,and defense protein activity than SA doses at 1 and 2 mmol L–1.These results suggested that SA at 1.5 mmol L–1can be used for plant defense induction (Waret al.2011).
H2O2as a reactive oxygen species is the earliest reaction of plant–pathogen interactions in plant cells(Bolwell 1999;Bastas 2014).H2O2acts as a local signal for the hypersensitive response (HR) that contributes to rapid localized cell death (program cell death;PCD)localized in pathogen-infected sections followed by defense enzymes that activate defensive genes (Alvarezet al.1998;Kuzniak and Urbanek 2012).In addition,H2O2is involved in the reinforcement processes of the plant cell wall,xylem,and cell structures (Dempsey and Klessig 1995;Kuzniak and Urbanek 2012).Our results showed a substantial increase in H2O2accumulation when using 100 mg L–1or more of SA-Ricemate (39–61%) at 24 h post-inoculation.This result indicates that the SA-Ricemate elicitor activates an internal signal of oxidative bursts needed for the induction of defense-related genes.Haoet al.(2014) stated that the defense induction ofSalvia miltiorrhizaby SA led to the enhancement of H2O2development.Similarly,Deenamoet al.(2018) reported that the treatment of priming rubber tree seedlings with 5 mmol L–1of SA post-inoculation ofPhytophthora palmivoraresulted in a substantial increase in H2O2content by 9.44-fold causing induced resistance (IR) to terminate the invading pathogen (Haoet al.2014;Deenamoet al.2018).Niu and Liao (2016)observed that the production of H2O not only contributed to stress-induced cell death,but also the growth of plant development and abiotic response (Niu and Liao 2016).MDA is one of the secondary lipid peroxidation products.It was commonly used as a biomarker for cell membrane damage (Corbineauet al.2002;Gawełet al.2004).Numerous studies have shown that the HR is linked to the accumulation of MDA.In this analysis,the observed MDA levels at 24 h post-inoculation are higher in samples treated with 100,150,200,250,and 300 mg L–1of SA-Ricemate compared to untreated samples.This indicates that superoxide radicals are generated in the SA-Ricemate elicitor treated samples which increases lipid peroxidation that can potentially kill cell membranes during the infection phase leading to cell death and significant symptoms of HR.These results are consistent with the work presented by Singh and Upadhyay (2014) who also found lipid peroxidation as well as MDA in tomato plants treated with fusaric acid to induce cell death which increased significantly after 8 h of treatment and peaked at 48 h (Singh and Upadhyay 2014).Similarly,Zhanget al.(2015) reported that a foliage application of 100 mmol L–1of glycerol enhanced disease resistance in cacao by stimulating the levels of pathogenesis-related (PR) genes and increased the level of MDA content (Zhanget al.2015).Chlorophyll is the main pigment used in photosynthesis to transform light energy into chemical energy for plant growth (Pallardy 2008).The increase in chlorophyll content would also improve photosynthesis.SA is also a part of the regulation of photosynthesis.Several previous studies have shown that the application of exogenous SA has affected plant growth and photosynthesis (Panchevaet al.1996;Hayatet al.2013).In an earlier study,Nazaret al.(2015) identified the use of SA in plants with and without stress,developing a significant increase in photosynthetic parameters and chlorophyll contents by proline synthesis,nitrate reductase activity ATP-sulfurylase,and inhibition of aminocyclopropane carboxylic acid synthase (Nazaret al.2015).In our results,after spraying rice plants with 50,100,150,200,and 250 mg L–1of SA-Ricemate elicitor three times,the chlorophyll content (chlorophylla,chlorophyllb,and total chlorophyll) was significantly increased in comparison to the untreated samples.These results are consistent with the work reported by Razmiet al.(2017) which found that the foliar application of SA to soybean plants at 0.4 mmol L–1significantly increased the chlorophyll content (15% chlorophyllaand 19% chlorophyllb) compared to the control (Razmiet al.2017).The work proposed by Moussa and El-Gamal(2010) stated that the application of 500 μmol L–1of SA to wheat (Triticum aestivum) seed priming increased the concentration of chlorophyll,CO2fixation,and phosphoenolpyruvate carboxylase (Moussa and El-Gamal 2010).In the same line,our greenhouse experiments have shown that the SA-Ricemate elicitor at 100,150,200,and 250 mg L–1was successful in controlling BLB disease and improving photosynthesis capability.
To further determine the effect of SA-Ricemate elicitor for controlling the BLB disease,growth parameters,and yield;two consecutive experiments were performed on field trials.SA has long been recognized as a regulatory agent for plant protection reactions based on the activation of systemic acquired resistance (SAR) in plants (Furniss and Spoel 2015).Several studies have shown that plant priming with SA can reduce the severity of pathogen attacks (Aranega-Bouet al.2014;Dempsey and Klessig 2017).In our research,the use of the SA-Ricemate elicitor at 100 mg L–1significantly decreased BLB disease severity with a control effect of 35–86% compared to untreated samples.This observation was consistent with the findings of Soodet al.(2013) who reported that the use of 50 mg kg–1of SA had a positive impact on sheath blight disease by the accumulation of defense-related enzymes including phenylalanine ammonia-lyase,superoxide dismutase,chitinase,β-1,3-glucanase,and phenols in rice leavescv.Pusa Basmati I (Soodet al.2013).Similarly,Hadi and Balali (2010) reported that the use of SA at 0.2 mmol L–1decreased the number of infected potato spots caused byRizoctonia solani(Hadi and Balali 2010).In our research,the application of the SA-Ricemate at 100 mg L–1significantly increased plant height relative to untreated samples.This reported change was not substantially different from the use of a commercial elicitor COS®treatment.This result confirms the findings of Sadeghipour (2012) which described pre-treatment for common bean (Phaseolus vulgaris) by soaking with 0.25–0.75 mmol L–1of SA leading to an increase in plant height under water stress (Sadeghipour 2012).Gorni and Pacheco (2016) also found that 0.5 mmol L–1of SA had a significant increase of 83.11% in the dry mass ofAchillea millefoliumL.(Asteraceae) roots (Gorni and Pacheco 2016).Similarly,Parasharet al.(2014) reported that 0.01 mmol L–1of SA applied toBrassica junceaplants increased the dry root mass shoot by 26 and 51%,respectively (Parasharet al.2014).It is well known that SA also encourages the division and elongation of plant cells (Hayatet al.2013).SA also acts as a stimulant for plant cells to stabilize plants under environmental stresses such as drought,extreme temperatures,and heavy elements (Mousaviet al.2009;Saleh and Abduljabbar 2019).Our field experiment results have shown that SA-Ricemate elicitor can effectively increase yield as tillers per hill,panicles per hill,grains per panicle,and 1 000-grain weight with an overall crop yield increase of 25–69%.The two key different functions of SA are plant defense and growth promotion which have contributed to a general increase in rice production (Silvermanet al.1995;Hayatet al.2013).Reddyet al.(1979) identified a linear relationship between the severity of BLB disease at the plant growth stage and the grain yield.The increase in BLB severity has been shown to have a reduced effect in terms of grain yield,1 000-kernel weight,and fertility(Reddy 1979).Similar to Nohet al.(2007),the authors reported a decrease in rice yield and brown head rice rate of more than 50% was observed at 29% disease of leaf infected diseases (Nohet al.2007).In addition,the effects of SA as a plant growth regulator function directly in metabolic pathways as a response to abiotic stress.This process may be triggered by SA signaling to other hormones such as jasmonic acid,ethylene,and auxin which affect plant growth and development (Smithet al.2017).These results are consistent with the work presented by Kareemet al.(2019) which stated that the exogenous application of SA at 1.44 mmol L–1can promote shoot growth,final wheat biomass,and yield components(spike drying weight,grain dry weight,and 1 000-grain dry weight.It is also suggested that a pre-treatment based on SA-Ricemate can increase the productivity of rice by improving tolerance to biotic and abiotic stress.
In the present study,we performed molecular docking of SA on catalase and aldehyde dehydrogenase related to lipogenesis.To study the interaction between two molecules,molecular docking is the widely used method in computational studies to develop structure-based drugs.Also in previous studies,SA inhibitory and binding studies were documented (Thepbanditet al.2021).However,the mechanism of catalase and aldehyde dehydrogenase inhibition by SA was not yet studied using docking methods.Using docking,we revealed that SA binds to amino acids in the active site of catalase and aldehyde dehydrogenase and hence may inhibit catalase and aldehyde dehydrogenase activity which may increase the levels of H2O2and MDA.In most of the docking algorithms rigid or flexible protein structure is considered for the interaction with compounds.Generally,inflexible docking,algorithm side-chain fluctuations are taken into consideration and multiple confirmations of compounds are used to find a better docking complex.For this,a better structure of the protein is needed to get good docking results and depends on the quality of the protein structure.The docking score of SA and the binding poses suggest that SA inhibits catalase and aldehyde dehydrogenase by binding to the amino acids involved in the active site.Even though no experimental data is available regarding structures,the use of molecular docking can simulate interactions and binding scores.To improve docking studies towards accurate results,molecular dynamics of the docked complex were used and this corrects the inaccurate structural conformational change.The complex formed by SA after docking was improved by molecular dynamics simulations by correcting the interaction of the complex.From the molecular interactions,it was confirmed that SA formed a good ligand-protein complex with catalase and aldehyde dehydrogenase showing the possible direct involvement in the mechanisms.This finding may indicate that SA directly inhibits catalase and aldehyde dehydrogenase.
5.Conclusion
The SA-Ricemate elicitor increased resistance toXooby emphasising a fast plant defence compound containing H2O2and MDA.In addition,the SA-Ricemate elicitor increased overall chlorophyll content and yield.As a result,SA-Ricemate is a viable option for reducing BLB disease and increasing productivity in rice fields.Catalase and aldehyde dehydrogenase-binding sites were found using the CASTp Software.SA was created and refined in Chemsketch before docking to catalase and aldehyde dehydrogenase-binding sites in GOLD 3.0.1.Docked SA conformations with catalase and aldehyde dehydrogenase-binding sites.The catalase amino acids GLN7,VAL27,and GLU38 formed a strong hydrogen bond with SA when SA was present.The amino acids involved in aldehyde dehydrogenase were LYS5,HIS6,and ASP2.
Acknowledgements
This work was supported by the Suranaree University of Technology,Thailand,the Thailand Science Research and Innovation (TSRI),and the National Science,Research and Innovation Fund,Thailand (NSRF) (90464).
Declaration of competing interest
The authors declare that they have no conflict of interest.
杂志排行

Journal of Integrative Agriculture的其它文章
- Less hairy leaf 1,an RNaseH-like protein,regulates trichome formation in rice through auxin
- Characterization of a blaCTX-M-3,blaKPC-2 and blaTEM-1B co-producing lncN plasmid in Escherichia coli of chicken origin
- Consumers’ experiences and preferences for plant-based meat food: Evidence from a choice experiment in four cities of China
- Farmers’ precision pesticide technology adoption and its influencing factors: Evidence from apple production areas in China
- Visual learning graph convolution for multi-grained orange quality grading
- lnfluence of two-stage harvesting on the properties of cold-pressed rapeseed (Brassica napus L.) oils