秋石斛钙依赖蛋白激酶基因的克隆及表达分析
2023-01-04林榕燕钟淮钦陈艺荃方能炎叶秀仙
林榕燕,钟淮钦,陈艺荃,方能炎,孔 兰,叶秀仙*
(1 福建省农业科学院作物研究所/福建省特色花卉工程技术研究中心,福州 350013;2 福建省农业科学院农业工程技术研究所,福州 350003)
石斛(Dendrobiumspp.)是兰科(Orchidaceae)石斛属(Dendrobium)多年生附生草本植物,根据开花时间和花序着生位置常将其分为春石斛和秋石斛[1]。秋石斛在秋季开花,花序着生于假鳞茎顶端,着花数5~20朵,其花姿优雅、花色艳丽、花期长,是观赏兰花中的重要切花和盆花,在国际花卉市场上扮演着重要角色[2]。秋石斛原产地为东南亚和西太平洋岛屿等热带亚热带地区,其在生长过程中对温度较为敏感,不适宜的温度会导致秋石斛花朵的产量与质量的下降,甚至停止生长。而在中国大部分地区,低温仍是限制秋石斛生长发育的重要逆境因子之一[3]。因此,提高秋石斛的抗寒性以降低低温伤害是秋石斛产业急需解决的主要问题,而获得高抗寒性的秋石斛品种则是解决该问题的关键。其中开展秋石斛抗寒基因的挖掘与鉴定,是培育高抗寒性秋石斛新品种的一项重要工作。
钙依赖蛋白激酶(calcium-dependent protein kinase,CDPK/CPK)是一类在植物中广泛存在的依赖钙离子的 Ser/Thr 型蛋白激酶,是一类重要的钙感受器,在植物的生长发育、应答胁迫反应中具有重要作用[4]。目前,模式植物和一些作物的CDPK基因家族成员已经被分离和鉴定,拟南芥[5]中存在34个CDPK基因家族成员,玉米[6]中存在40个CDPK基因家族成员。此外,铁皮石斛[7]、广东桑[8]和番茄[9]等多种植物中的CDPK基因也陆续被报道[10-13]。随着对CDPKs研究的深入,发现其能通过识别和参与细胞内Ca2+信号转导来响应低温、干旱、高盐等多种逆境胁迫的生物学功能,如过表达BnCPK5能增强油菜的干旱胁迫耐受性[14];玉米ZmCDPK6被证实不仅相应高温胁迫,还受到盐胁迫和低温胁迫诱导表达[15];蝴蝶兰PaCDPK1在应答低温、机械损伤和病原菌侵染时的转录水平提高[16]。
鉴于CDPK基因在植物响应逆境胁迫中的重要性,特别是在应对低温胁迫方面,推测秋石斛CDPK中可能有相似的生物学功能。因此,本研究中通过克隆秋石斛CDPK基因,对其生物信息学进行分析,并对其在低温胁迫下的表达情况进行研究,有助于后续秋石斛CDPK基因的功能验证,为探明秋石斛抗寒的分子机制奠定基础。
1 材料和方法
1.1 材 料
以生长状况良好的秋石斛品种‘水芙蓉’为供试材料,取适量叶片于液氮速冻后进行RNA提取,用于CDPKs基因克隆。分别取‘水芙蓉’的根、茎、叶、花用于CDPKs基因的组织表达分析;分别取长势相近的4个秋石斛品种(‘三亚阳光’、‘四面佛’、‘水芙蓉’及D128)叶片用于基因的表达分析和相对电导率的测定。取长势相近的‘水芙蓉’分别于0 ℃和8 ℃条件下处理12 h,以置于资源圃常规条件(25 ℃)下的植株作为对照,于相应时间点取叶片用于检测低温胁迫下基因的表达情况及相对电导率变化情况。

表1 秋石斛CDPK基因克隆及荧光定量PCR所用引物
1.2 试验方法
1.2.1 秋石斛总RNA的提取及cDNA第一链的合成参照林榕燕等[17]的方法,采用通用植物总RNA提取试剂盒(BioTeke)进行秋石斛总RNA提取,采用PrimeScriptTMⅡ 1st Strand cDNA Synthesis Kit试剂盒(TaKaRa)进行cDNA 第一链的合成。
1.2.2 秋石斛CDPK基因cDNA序列扩增从NCBI上下载大花蕙兰、铁皮石斛的CDPK基因序列,根据同源克隆的原则设计了CDPK基因序列特异引物CDPK-F/CDPK-R(表1),以合成的cDNA第一链为模板,进行PCR扩增。扩增体系为25 μL,包含了2×PCR buffer for KOD Fx 12.5 μL、2.0 mmol/L dNTP 5 μL,1 U/μL KOD Fx 0.5 μL,ddH2O 4 μL,100 ng/μL cDNA模板1 μL,10 μmol/L上、下游引物各1 μL。PCR扩增程序:98 ℃预变性 2 min;98 ℃变性 30 s,53 ℃退火30 s,68 ℃延伸2 min 30 s,共35个循环;68 ℃延伸7 min。PCR扩增产物通过琼脂糖(1%)电泳进行检测后,回收目的片段,经连接、转化,挑选阳性克隆送至测序公司测序完成。
1.2.3 生物信息学分析将测序获得的序列翻译成氨基酸序列后,利用ExPASy、CBS Prediction Servers、WoLF PSORT、Conserved Domain Search、SOPMA、DNAMAN、MEGA5.0等软件进行CDPKs 蛋白质的理化性质、跨膜结构、亚细胞定位、保守结构域、二级结构、序列多重比对分析及系统进化树构建。
1.2.4 秋石斛DenCDPKs基因定量表达分析以秋石斛不同品种不同处理材料的cDNA为模板,以Actin为内参基因,利用引物(表1),采用TB Green Premix Ex TaqTM试剂盒在7500 Real-Time PCR System仪(美国ABI公司)上检测DenCDPKs基因在不同材料中的表达情况。qRT-PCR反应体系为10 μL,包含TB Green Premix Ex Taq 5 μL,ROX Reference DyeⅡ 0.2 μL,上、下游引物0.4 μL,cDNA模板1 μL,ddH2O 3 μL。反应程序:95 ℃预变性 1 min;95 ℃变性 5 s,60 ℃退火30 s,72 ℃延伸 45 s,共40个循环。每个反应设3次重复,采用2-ΔΔCt法对数据进行定量分析。
1.2.5 秋石斛叶片相对电导率测定以秋石斛不同品种不同处理条件下的叶片为材料,对其相对电导率进行测定,具体方法参照徐新娟等[18]的方法,并做适当修改。先测量材料处于室温下浸泡24 h后的电导率(E1),而后将其置于100 ℃ 沸水浴中20 min,待冷却至室温后,再次测量电导率(E2),相对电导率(REC/%)=E1/E2×100%。
1.3 数据分析
试验数据采用Execl 2007软件进行分析,显著性分析采用SPSS 20.0软件进行。
2 结果与分析
2.1 秋石斛CDPK基因序列的获得
经PCR扩增,分别得到约2 000、2 000和2 300 bp左右的特异性条带(图1)。测序完成后,将结果提交至GenBank 数据库进行在线Blast,发现获得的序列与铁皮石斛的CDPK1、CDPK2、CDPK3基因序列的一致性分别达96.64%、95.89%和94.49%,说明所得片段序列为秋石斛CDPK1、CDPK2、CDPK3基因序列,命名为DenCDPK1(GenBank登录号为MZ322902)、DenCDPK2(GenBank登录号为MZ322903)、DenCDPK3(GenBank登录号为MZ322904)。DenCDPK1基因片段大小为1 934 bp,包含一个长度为1 605 bp的完整开放阅读框(219~1 823),编码534个氨基酸。DenCDPK2基因片段大小为1 971 bp,包含一个长度为1 626 bp的完整开放阅读框(220~1 845),编码541个氨基酸。

M. DL2000;1. DenCDPK1;2. DenCDPK2;3. DenCDPK3图1 秋石斛CDPK基因的克隆Fig.1 Cloning of CDPK gene in Dendrobium spp.
DenCDPK3基因片段大小为2 302 bp,包含一个长度为1 611 bp的完整开放阅读框(410~2 020),编码536个氨基酸。
2.2 秋石斛DenCDPK蛋白质生物信息学分析
秋石斛DenCDPK1、DenCDPK2、DenCDPK3蛋白质的理论分子量分别为59.59、60.49和60.44 kD,等电点分别为6.03、6.14和6.62,分子式分别为C2643H4182N714O808S22、C2648H4222N764O806S26和C2680H4269N747O803S20,总平均疏水指数(GRAVY)分别为-0.447、-0.480和-0.509,说明三者均为亲水蛋白质,且三者均不具有跨膜结构。二级结构预测结果显示,三者二级结构均以α-螺旋(44.01%、47.87%和45.34%)和无规则卷曲(36.70%、34.57%和36.38%)为主,β-转角(10.30%、9.61%和9.51%)和延伸链(8.99%、7.95%和8.77%)所占比率较低。
此外,DenCDPK1中Lys所占比重较大,比例达8.2%,Trp含量最低,仅为0.7%;定位于叶绿体的可能性最大;该蛋白质不稳定系数为42.55,表明它是一个不稳定蛋白质。DenCDPK2中Gly所占比重较大,比例达9.2%,Trp含量最低,仅为1.1%;定位于细胞质的可能性最大;该蛋白质不稳定系数为44.88,表明它是一个不稳定蛋白质。DenCDPK3中Lys所占比重较大,比例达9.7%,Trp含量最低,仅为0.9%;定位于细胞质的可能性最大;该蛋白质不稳定系数为35.18,将其归为稳定蛋白质。
2.3 秋石斛DenCDPK蛋白质序列一致性比对与进化分析
Blast比对结果显示,DenCDPK1氨基酸序列与铁皮石斛(Dendrobiumofficinale,AGA83664.1)的一致性为97%,与桃红蝴蝶兰(Phalaenopsisequestris,XP_020577380.1)的一致性为92%,与深圳拟兰(Apostasiashenzhenica,PKA45643.1)的一致性为84%。DenCDPK2氨基酸序列与铁皮石斛(Dendrobiumofficinale,XP_020684921.1)的一致性为99%,与桃红蝴蝶兰(Phalaenopsisequestris,XP_020594551.1)的一致性为96%,与深圳拟兰(Apostasiashenzhenica,PKA50602.1)的一致性为91%。DenCDPK3氨基酸序列与铁皮石斛(Dendrobiumofficinale,AGI21023.1)的一致性为97%,与桃红蝴蝶兰(Phalaenopsisequestris,XP_020574779.1)的一致性为94%,与深圳拟兰(Apostasiashenzhenica,PKA49856.1)的一致性为87%。利用DNAMAN软件进行序列多重比对分析,发现不同植物的CDPK之间具有较高的一致性,且这些序列都具有 CDPK 家族特有的保守结构域,从氨基末端开始有STKc_CAMK结构域(Ser/Thr蛋白激酶区)和4个EF-hand结构域(图2)。

Den.秋石斛;Do.铁皮石斛;As.深圳拟兰;虚线为Ser/Thr蛋白激酶区;实线为4个 EF-hand 结构图2 秋石斛与其他植物CDPK氨基酸序列多重比对Den. Dendrobium spp.; Do. Dendrobium officinale; As. Apostasia shenzhenica; Dotted lines indicate Ser/Thr protein kinase domain; Solid lines indicate four EF-hand structuresFig.2 Multiple alignment of the amino acid sequences of CDPK among Dendrobium spp. and other plants
从NCBI数据库中下载拟南芥的34个CDPK蛋白质序列及铁皮石斛的3个CDPK蛋白质序列,采用MEGA5.0软件构建CDPK系统进化树(图3)。参照洪旭升等[19]的研究报道,将构建的系统进化树分为4个组,其中DenCDPK1先与DoCPK1聚为一类,并于AtCDPK9和AtCDPK33的亲缘关系较近,位于第Ⅱ组;DenCDPK2先与DoCDPK3聚为一类,并与AtCDPK13的亲缘关系较近,DenCDPK3先与DoCPK3聚为一类,并与AtCDPK14和AtCDPK32的亲缘关系较近,且DenCDPK2和DenCDPK3位于第Ⅲ组。

Den.秋石斛;Do.铁皮石斛;At.拟南芥图3 DenCDPK系统进化树Den. Dendrobium spp.; Do. Dendrobium officinale; At. Arabidopsis thalianaFig.3 Phylogenetic tree of DenCDPK
2.4 秋石斛CDPK基因表达分析
以秋石斛‘水芙蓉’根、茎、叶、花的cDNA为模板,通过实时荧光定量PCR分析DenCDPKs基因的表达水平(图4)。DenCDPK1和DenCDPK3基因的相对表达量表现为叶>根>花>茎,DenCDPK2基因的相对表达量表现为叶>茎>根>花,三者在叶中的相对表达量均显著高于在其他组织中的相对表达量,其中DenCDPK3基因在叶中的相对表达量是其在茎中的10倍。
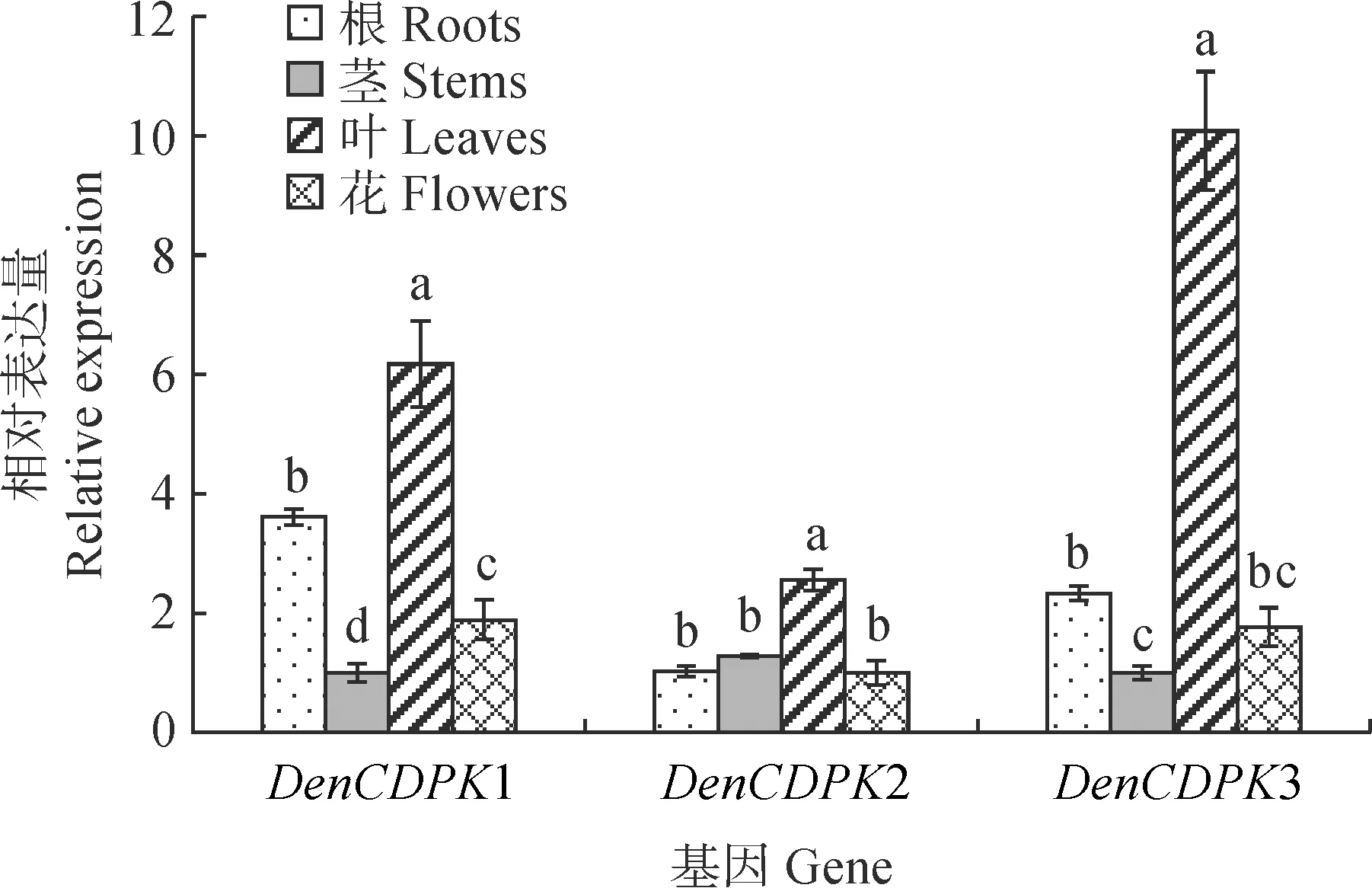
不同小写字母代表差异显著(P<0.05),下同图4 秋石斛DenCDPK在不同组织中的相对表达量Different normal letters represented significant difference (P<0.05); the same as belowFig.4 Expression levels of DenCDPK in different tissues of Dendrobium spp.
以秋石斛不同品种‘水芙蓉’、‘三亚阳光’、D128和‘四面佛’叶片的cDNA为模板,通过实时荧光定量PCR分析DenCDPKs基因的表达水平(图5)。DenCDPK1基因的相对表达量表现为‘水芙蓉’>‘四面佛’>‘三亚阳光’>D128,其中‘水芙蓉’叶片中该基因的相对表达量是D128的12倍。DenCDPK2基因的相对表达量表现为‘水芙蓉’>D128>‘三亚阳光’>‘四面佛’,其中‘水芙蓉’叶片中该基因的表达量是‘四面佛’的100倍。DenCDPK3基因的相对表达量表现为‘四面佛’>D128>‘水芙蓉’>‘三亚阳光’,其中‘四面佛’叶片中该基因的表达量是‘三亚阳光’的88倍。

图5 秋石斛DenCDPK在不同品种中的相对表达量Fig. 5 Expression levels of DenCDPK in different varieties of Dendrobium spp.
以秋石斛‘水芙蓉’在常规条件(25 ℃,对照组)和低温处理(8 ℃、0 ℃)12 h后的叶片cDNA为模板,通过实时荧光定量PCR分析DenCDPKs基因的表达水平(图6)。DenCDPK1和DenCDPK2基因的相对表达量随着处理温度的降低表现为先上升后下降的趋势,在处理温度为8 ℃时,基因的相对表达量显著高于对照组和0 ℃处理组。DenCDPK3基因的相对表达量随着处理温度的降低表现为不断上升的趋势,在处理温度为8 ℃时,DenCDPK3基因的相对表达量显著高于对照组;在处理温度为0 ℃时,DenCDPK3基因的相对表达量显著高于对照组和8 ℃处理组。

图6 秋石斛DenCDPK在不同条件下的相对表达量Fig.6 Expression levels of DenCDPK under different conditions of Dendrobium spp.
2.5 不同条件下秋石斛叶片中相对电导率的比较
通过比较秋石斛不同品种叶片在常温下的相对电导率(图7),发现相对电导率在4个品种中的大小依次为‘四面佛’、‘三亚阳光’、D128和‘水芙蓉’,且各品种间的相对电导率呈显著差异。

图7 不同品种秋石斛叶片的相对电导率Fig.7 Relative electrical conductivity of leaves in different varieties of Dendrobium spp.
通过比较秋石斛‘水芙蓉’在常规条件(25 ℃,对照组)和低温处理(8 ℃、0 ℃)12 h后叶片的相对电导率(图8),发现经低温处理12 h后,叶片中的相对电导率均显著提高,且随着处理温度的降低,叶片的相对电导率呈上升趋势,在处理温度为0 ℃时,显著高于对照组和8 ℃处理组。
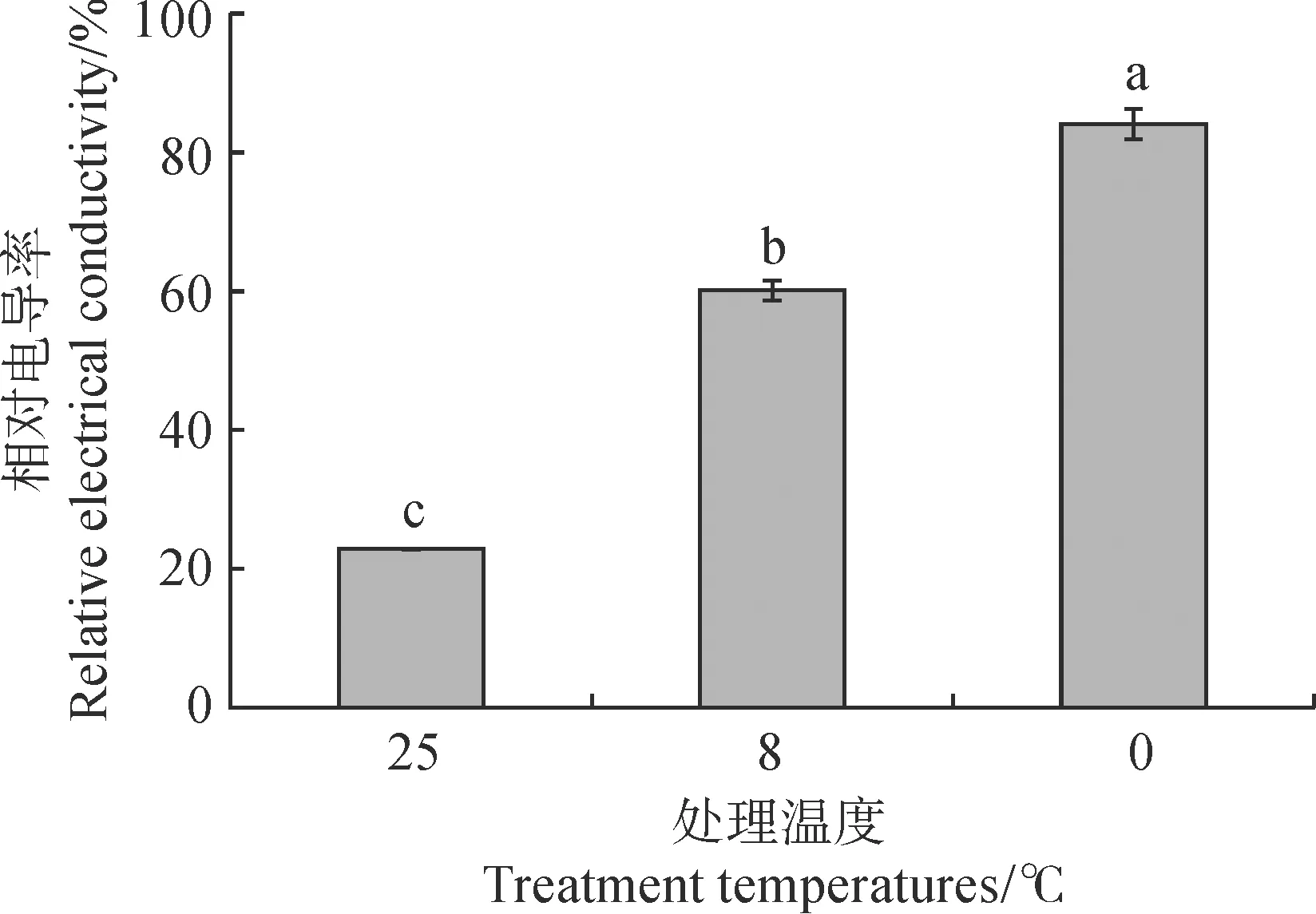
图8 不同处理条件下秋石斛叶片的相对电导率Fig.8 Relative electrical conductivity of leaves under different conditions in Dendrobium spp.
3 讨 论
钙依赖蛋白激酶(CDPK)作为钙结合蛋白之一,是植物细胞信号转导途径中的重要一环。近年来,越来越多的植物CDPKs基因被分离鉴定,并被报道广泛参与植物的生长发育和抵抗逆境胁迫的调控反应[20-22]。本研究在秋石斛‘水芙蓉’叶片中克隆获得了3个CDPK基因,将其命名为DenCDPK1、DenCDPK2和DenCDPK3,分别编码534、541和
536个氨基酸。分析3个基因编码氨基酸的理化性质与结构特征,发现三者的基本理化性质与结构特征极为相似,均为亲水蛋白质,且存在STKc_CAMK结构域,这与报道的铁皮石斛[23]、青花菜[24]等植物的序列结构特征一致。三者的差异在于DenCDPK1定位于叶绿体的可能性最大,而DenCDPK2、DenCDPK3定位于细胞质的可能性最大,这与在细胞膜、细胞质、线粒体、叶绿体、染色体和细胞核中均有 CDPKs存在的现象相符[25]。
CDPKs基因的表达存在组织特异性,辣椒CaCDPK09、CaCDPK18等基因在各组织中均有较高表达量,CaCDPK28、CaCDPK04则在各组织表达量都很低,而CaCDPK02、CaCDPK03等基因只在花芽和花中表达,CaCDPK07、CaCDPK14等只在根、茎、叶中高表达[26]。本研究中DenCDPKs在根、茎、叶、花中均有表达,DenCDPK1和DenCDPK3基因在叶中高表达,在茎中低表达;DenCDPK2基因同样在叶中高表达,但在花中低表达。
CDPKs基因表达还受温度、盐胁迫、植物激素等的影响,羊草LcCDPK在高浓度盐碱处理条件下上调表达[27];朝鲜淫羊藿EkCDPK在干旱胁迫后15 h内表达量显著高于对照组[28];马铃薯StCDPK基因的表达量在4℃处理12 h时出现峰值[29]。本研究中,DenCDPK1、DenCDPK2和DenCDPK3基因分别在处理温度为8 ℃、8 ℃和0 ℃时相对表达量显著高于对照组,说明了DenCDPKs受到低温诱导表达,推测DenCDPKs基因参与到秋石斛低温胁迫响应过程。
有研究表明,相对电导率在一定程度上反映了膜脂的损伤程度,与抗寒性呈负相关,可在一定程度上体现植物的抗寒性[30]。本研究中叶片的相对电导率随温度降低呈上升趋势,在0℃时相对电导率最大,这一变化趋势与DenCDPK3基因在低温处理下的表达模式一致,说明DenCDPK3基因可能参与秋石斛低温胁迫的应激反应。研究还发现相对电导率在4个品种中的大小为‘四面佛’>‘三亚阳光’>D128>‘水芙蓉’,初步判断4个秋石斛品种的抗寒性大小为‘水芙蓉’>D128>‘三亚阳光’>‘四面佛’,这一结果与前期通过LT50评价法和隶属函数分析法综合评价的秋石斛抗寒性大小一致[31],也与DenCDPK2基因在不同品种叶片的表达模式相符,说明DenCDPK2基因的表达与秋石斛的抗寒性有一定的相关性。此外,系统进化树分析发现,DenCDPK2与AtCDPK13的亲缘关系较近,而前人的研究证实了AtCDPK13在拟南芥保卫细胞中的过表达抑制了光诱导的气孔开放,推测DenCDPK2可能有相似功能[32]。
以上结果均表明,本研究克隆获得的DenCDPKs是可能参与秋石斛低温胁迫应对的钙依赖型蛋白激酶,特别是DenCDPK2可能在秋石斛抗寒过程中发挥着重要作用,但其在低温胁迫下的调控机制和功能还需进一步研究,后续将通过分子生物学手段调节CDPK基因的表达,分析下游基因调控网络,揭示CDPK基因的作用机理,进而为研究秋石斛抗寒机制奠定基础,同时也为今后的品种创新提供基因资源。
