Context dependent song-flight performance and timing in the Black-headed Bunting(Emberiza melanocephala)
2023-01-03SnjBrisiDvorCikoviVesnTutiJelenKrljHerertHoi
Snj Briˇsi′c,Dvor′Cikovi′c,Vesn Tutiˇs,Jelen Krlj,Herert Hoi
a Institute of Ornithology,Croatian Academy of Sciences and Arts,Zagreb,Gunduli′ceva 24,10000,Zagreb,Croatia
b Konrad Lorenz Institute of Ethology,Department of Interdisciplinary Life Sciences,University of Veterinary Medicine Vienna,Savoyenstraße 1A,1160,Vienna,Austria
Keywords:Fertilisation probability Mate attraction Moth song-flight Towering song-flight
ABSTRACT Many songbirds produce song-flights;however,the function,vocal and motor characteristics,as well as the diel and seasonal variation of song-flight in songbirds remain not well understood.Here,we studied two types of songflight in male Black-headed Buntings(Emberiza melanocephala),the Moth—a standard,i.e.,perch song produced during a horizontal flight,and the Towering—a specifically structured song produced during a complex ascending and descending flight.While perch song,used during Moth flight,has already been described,here we provide the first description and sonogram representation of the more elaborate and less stereotyped Towering song.While males started to perform Moth song-flights as soon as they arrived at the breeding site,Towering did not start before the female arrival.Males usually delivered spontaneous Moth song-flights before chasing rivals or undertaking aerial fights,suggesting Moth is directed towards conspecific males and serves as a threat display.Furthermore,playback of conspecific perch songs triggered males to approach the loudspeaker with Moth songflights.The Towering started after females arrived at the breeding site,suggesting the Towering is directed towards conspecific females.We detected a seasonal difference in the peak Towering rate between forenoon and afternoon,suggesting that Towerings performed at different times of the day have different functions.Forenoon Towerings were strongly positively related to the number of fertile females at the breeding site,while afternoon Towerings peaked very early in the breeding season and coincided with the number of females at the breeding site before the onset of nest-building.These results provide evidence that birds can direct different types of song-flight towards different receivers and suggest the possibility that time of the day determines the context of the display,namely to attract social mates or solicit copulation.Further work is needed to account for the effect of between male variability in song-flight rate,as well as in height/steepness of flight or song quality.
1.Introduction
Females use various clues to assess the quality of males,including ornaments,vocalisations and behavioural displays(Andersson,1994).If such traits are costly to produce,they constitute a handicap and can be regarded as honest signals(Iwasa et al.,1991;Walther and Clayton,2005).In this context,differences in behavioural performance could reflect how well a male is able to cope with the handicap and hence demonstrate its quality(Byers et al.,2010).Males in many species can concurrently demonstrate their quality by performing multimodal signals.Different components of a multimodal signal could relay either distinct messages or the same message about the signaller's quality(Møller and Pomiankowski,1993;Candolin,2003).Males can also use different components of a multimodal signal and/or signal in different environmental settings to convey distinct information to receivers.
In birds,a song-flight is a multimodal signal as it combines a vocalisation with a flight display.Furthermore,body postures during the flight can emphasise male ornaments in some species,i.e.,long tails or intense colours of specific body parts(Emlen,1957;De Benedictis,1966).Flight displays are thought to be energetically costly(Hedenstrom and Møller,1992)or physically demanding(Usherwood,2008)and may impose a cost through increased predation risk(Zuk and Kolluru,1998).Hence,courtship flight performance could reliably signal a male's innate quality and condition(Simmons,1988;Mather and Robertson,1992).Courtship flight performance could also be used as a determinant for specific skills,for example,a male's foraging ability(Halupka and Borowiec,2006)and its qualification for the care of offspring(Ritchison,1991;Balsby,2000;Lang,2011).In addition,take-off abilities,which are paramount in predator avoidance,are condition-dependent,i.e.,heavier individuals are hampered by their weight(Krams,2002).Following the same argument,heavier individuals would need more muscle power to perform the song-flight or would need to spend more energy on performance.
Even though many bird species possess a song-flight display,only a few studies have investigated song flight performance and looked at its function(Meril¨a and Sorjonen,1994;Sorjonen and Merila,2000;Pereyra and Morton,2010).One way to infer the function of a display is to examine its timing and the frequency in the breeding cycle(Ballentine et al.,2003;Taff et al.,2014).For example,if a flight display is used to advertise territorial claims,it should be expected to peak immediately after male arrival at the breeding grounds.On the other hand,if a flight display is used for mate attraction,it could peak upon female arrival or coincide with a peak in female fertility.Female fertility can be defined as the likelihood of fertilisation resulting from the insemination at a given time.It increases up to the onset of egg-laying(Birkhead,1998)and can vary diurnally depending on the timing of oviposition(Birkhead et al.,1996).Likewise,singing activity has pronounced temporal variability(Catchpole and Slater,2008).In line with this,a concurrence of peaks in male song-flight rates and female fertility could suggest their relationship.
Black-headed Bunting(Emberiza melanocephala)males perform two types of song-flight,namely the Moth song-flight(Almond,1946;Rokitansky,1969)and the Towering song-flight(Almond,1946).Song-flights in Black-headed Buntings have only been briefly mentioned in the literature(Cramp and Perrins,1994),and little is known about their function,variability,song structure,flight trajectory or body posture.The Moth song-flight,a level flight with shallow wingbeats and dangling legs,is a more common type,similar to a flight display of Corn Bunting(Emberiza calandra),while a towering type of song-flight,incorporating almost vertical ascent and“parachute”descent,occurs less commonly(Cramp and Perrins,1994).A similar towering display in Red-headed Bunting(Emberiza bruniceps)is associated with a peak courtship,while in the Black-headed Bunting it was reported only twice:in South-West Iran and Lebanon(Cramp and Perrins,1994).
The focus of this study was:(i)to describe vocal and motor characteristics of the song-flight displays of male Black-headed Buntings,and(ii)to evaluate potential functions of song-flights based on the social contexts and/or the timing of male song-flight behaviour in relation to female fertility.As Moth song-flight was a relatively simple display incorporating level flight and male standard,i.e.,perch song already described in Bariˇsi′c et al.(2018).Furthermore,it was straightforward to determine the context in which the Moth song-flight occurred(see Results).Consequently,more attention was given to the Towering song-flight.
2.Materials and methods
2.1.Bird species and study area
Black-headed Buntings mostly inhabit scrubland and arable areas in lowlands in their Mediterranean breeding area.They are long-distance protandrous migrants wintering in India(Cramp and Perrins,1994;′Cikovi′c et al.,2021).The Black-headed Bunting is a polygynous species with a very low and facultative paternal contribution to offspring care(authors'unpublished data).Males arrive at the study area in the first week of May and females approximately a week later(authors’unpublished data).Each male sings a single individual-specific song strophe;short and stereotyped,yet rather complex(Bariˇsi′c et al.,2018).
The study was conducted from 2011 to 2016 in Ravni Kotari,a geographic region in Croatia adjacent to the Adriatic Sea(44°10′N,15°25′E-43°45′N,15°50′E).We selected our main study plot(3 km2)in the shallow valley near the village of Dazlina,based on a study of Black-headed Bunting distribution and abundance carried out by the authors in 2010.The study plot supported a dense population of Blackheaded Bunting and represented the typical habitats of the region:traditional small-scale mosaic Mediterranean agricultural habitats intersected with natural vegetation in different stages of succession(eumediterranean garrigues and maquis and submediterranean deciduous shrubs).From 2013,the study was extended to cover an additional study plot near the village of Ivinj,5 km from the main study plot.This study plot also had a dense population of Black-headed Bunting and similar habitats as in the main study plot,but its vegetation was in later succession stages.
2.2.Fieldwork
2.2.1.Unstandardised observations,trapping and sound recording
The fieldwork started upon arrival of the birds at the breeding grounds(the beginning of May),until the end of the breeding activity(from the middle to the end of July,depending on the year).Work was carried out daily,usually from dawn to midday and from 17:00 to dusk.In case of heavy rain or strong wind,the fieldwork session was abandoned or truncated.Observation sessions lasted on average 5 h in the forenoon and 3 h in the afternoon.Conspecific song playbacks were broadcasted from loudspeakers to lure and catch males into mist nets.To increase trapping success,we used song playbacks of several males and,based on the male's reaction to the playback,occasionally switched between different songs.During the process of trapping,the focal male's behaviour was continuously monitored.Females were caught using custom made mist nets near the nest.We caught and colour-ringed 99 males and 26 females in total(2011–2016:Nmale=15,12,8,25,15,24;Nfemale=2,6,0,1,3,14).Furthermore,125 nests of Black-headed Buntings were located and monitored(2011–2016;Nnest=35,30,30,0,0,30).We repeatedly inspected the territories of marked males and visually verified the identity of territory owner.We also recorded all territory owners'audio.Trapping of Black-headed Buntings and recording of their vocalisation took place from 2011 to 2016,while from 2011 to 2013 and in 2016,we conducted unstandardised observations of marked birds'behaviour and monitored nests.
2.2.2.Focal male-level standardised observations
In 2011,20-min standardised observations(N=200)of focal males were conducted in the morning.The focal male's behaviour was noted in detail,including observations of the song-flights.These standardised observations were done after we finished trapping males(from 14 May-30 June),as we could not control for the effect of playback used to lure males.The number of observed song-flights per 20-min observation was low,and therefore it was not possible to differentiate seasonal and diurnal changes in male song-flight rates.
2.2.3.Neighbourhood-level standardised observations
In 2016,instead of documenting Towering song-flights of only one focal male at a time,we documented all Towerings detected in the hearing range of a researcher as well as the time spent by the researcher on fieldwork.An effort was made to allocate approximately the same amount of time to each male's territory.In 2016,we also monitored nests and related female fertility.The Towering rate per hour was calculated as the total number of Towerings detected over the observation session,divided by the number of hours sampled.To examine diurnal variation,observed Towering song-flights were divided into those that occurred in forenoon(from dawn to noon)and those that occurred in afternoon(from noon to dusk).Based on our observations,we estimated that females arrive at the breeding grounds approximately six days(6.3±1.0 days,N=15 nests)before they start to build the nest and that it takes about five days(4.6±0.2,N=10 nests)to complete the nest(Table 1).We compared female fertility(determined by the stage of the female nest)to the song-flight rate at the level of the studied population(i.e.,regardless of the male's identity).With this approach,we avoided relying on documenting specific behaviour related to mating,such as copulation or solicitation,given that these were difficult to observe in closed habitats.In addition,we avoided the need to find all the nests in a male'sneighbourhood.Unfortunately,Moth song-flight could not be distinguished by sound only(see Results)so the same approach could not be applied to document Moth song-flights.However,documenting the context in which the Moth song-flights were delivered was straightforward,which enabled us to infer its function.

Table 1 Schematic timetable representation of female Black-headed Buntings(Emberiza melanocephala)breeding stage variables.
All observations of Towering song-flights in the hearing range of a researcher were also documented in 2011–2013,but could not be adjusted for the observation time,hence the Towering rate per hour could not be calculated.Nevertheless,to account for the possible year effect,we compared Towering song-flight observations in 2011–2013 with the peaks in female fertility,i.e.,the number of females in the pre-laying stage(N2011=34,N2012=30,N2013=30).
2.3.Sound analysis of song-flights
To describe vocal characteristics of song-flight,sound recordings were made with a Roland R-05 digital audio recorder and a Telinga Pro-4 PIP microphone and parabola.Recordings were saved in WAV format at 44.1 kHz sampling frequency and 16-bit resolution.Altogether,over 70 h of audio recordings were analysed.Recordings were visualised in Avisoft-SASLab Pro 5.2.08(Specht,2014)with the following set of parameters:fast Fourier transform length 512,frame size 100%,window FlatTop,and temporal overlap 75%.These parameters enabled a frequency resolution of 86 Hz and a temporal resolution of 2.9 ms.
All original recordings were checked to identify and extract songs produced during flights.The same recordings were already used to describe a male's standard,i.e.,perch song(Bariˇsi′c et al.,2018)and flight songs were compared to a perch song of individual males.The Moth song could not be audibly distinguished from the perch song and thus was not further analysed.The Towering song differed from a perch song.Sounds used in the Towering song were checked against all other calls identified from the recordings.Towering songs produced by the same male were identified and visually compared to account for eventual intra-male variability.A quantitative description of the Towering song structure is based on 32 recordings of Towerings performed by 23 individual males recorded between 2011 and 2016.In most cases,only one Towering performance per individual male was recorded.Three males were recorded on two occasions,one male on three occasions,and one male on five occasions in two successive years.The duration of Towering song segments and the occurrence of different call types were quantified.These parameters were averaged among recordings of the same male.
2.4.Statistical analysis
2.4.1.Towering song intra-and inter-male variability
To examine within male variability in comparison to between male variability in the Towering song,we calculated the overall mean and standard deviation for the number of syllables in the Relax segment of the Towering song(see Results).This segment was selected as it showed the greatest complexity of syllable types.We determined the between-male coefficients of variation(CV=SD/mean).We calculated the withinmale coefficients of variation based on mean and SD values from five males whose Towering performance was recorded more than once.We then calculated the ratio of between-male to within-male coefficients of variation(CVb/CVw);a ratio>1 suggests that variation is larger between than within individuals,and a ratio>2 suggests that analysed parameter may be used in individual recognition(Christie et al.,2004;Vignal et al.,2008).
2.4.2.Seasonal variation in song-flight rates
To examine seasonal variation,the number of song-flights observed in May were compared to the number observed in June,for both the Moth and Towering.Seasonal Moth rates from 2011 were visualised in comparison to the number of females in different breeding stages(Fig.1).There was no indication of a relationship between Moth rates and the number of females in either of the breeding stages,thus was not explored further.Seasonal Towering rates from 2016 were also visualised in comparison to the number of females in different breeding stages(Fig.2).As there was an indication of a relationship between both the forenoon and afternoon Towering rates and the number of females in the early breeding stages,their potential relations were explored accordingly.
Mean and SE were reported unless otherwise indicated.Statistical tests were declared significant when p<0.05.
2.4.3.Forenoon and afternoon towering rates in relation to female breeding stage

Fig.1.The seasonal pattern in Moth song-flight rate(Moths/hour)in male Black-headed Buntings(Emberiza melanocephala)from Croatia in 2011 averaged across a 3-day interval,starting from May 14,2011(MF,Moth song-flight rate,primary y-axis),related to the number of females(secondary y-axis)in different breeding stages(PL,pre-laying stage;L,laying stage;I,incubation stage;N,nestling stage;F,fledgling stage).
Forenoon and afternoon Towering rates from 2016 were used to explore relations between the Towering rates and the female breeding stages.We tested separately whether the forenoon Towering rate(forenoon)or the afternoon Towering rate(afternoon)is related to:(1)the number of newly arrived females at the study plot where arrival day was on average six days before the start of the nest construction(farr),(2)the number of females in the pre-nest-construction stage,i.e.,an averaged six-day period between female arrival and the start of the nest construction(farr6),(3)the number of females in the pre-laying stage,i.e.,nest construction stage that lasted on average five days(PL),(4)the number of females in the laying stage,where the stage duration depended on the number of eggs laid(L),(5)the number of females in either pre-laying or laying stage(PLL).Variables representing incubation,nestling and fledgling stage were not used as predictors in modelling,as they peaked later in the breeding season(Fig.2B).It took approximately three days to inspect all territories in both study plots.Thus,all the variables have been averaged across a three-day interval.Due to the sequential nature of the data,they represent time series variables.Time series regression usually differs from a standard regression analysis because the residuals tend to be serially correlated(Metcalfe and Cowpertwait,2009).When this correlation is positive,which it is in our case,the estimated standard errors of the parameter estimates will tend to be underestimated,which may cause type I error,leading to rejecting a null hypothesis when it is actually true.Therefore,we used autoregressive integrated moving average(ARIMA)models to account for the autocorrelated structure of time series data(Metcalfe and Cowpertwait,2009).Computing was performed with R(R Core Team,2020)using an auto.arima function of the‘forecast’package,v8.11(Hyndman and Athanasopoulos,2018).The auto.arima function in R uses a variation of the Hyndman-Khandakar algorithm,which combines unit root tests,minimisation of the AICc and Maximum Likelihood estimation to obtain an ARIMA model(Hyndman and Khandakar,2008).The auto.arima function also handles regression terms via the xreg argument.The user specifies the predictor variables to include,but auto.arima selects the best ARIMA model for the errors.The AICc is calculated for the final model,and this value can be used to determine the best predictors.That is,the procedure should be repeated for all subsets of predictors to be considered,and the model with the lowest AICc value selected(Hyndman and Athanasopoulos,2018).Ranking of models using Akaike's Information Criterion(AICc;corrected for small sample size)was performed with‘MuMIn’package,v1.43.15(Barto′n,2019).The relative likelihood of each model was determined using model weights wi(Burnham and Anderson,2004).We used forenoon and afternoon Towering rates as response variables and all other variables as predictor variables.Autocorrelation of residuals of the best models was checked by visually inspecting acf and pacf functions and by performing the Box-Ljung test of the R Stats Package,v3.6.3.

Fig.2.The seasonal pattern in Towering song-flight rates(Towerings/hour)in male Black-headed Buntings(Emberiza melanocephala)from Croatia in 2016 averaged across a 3-day interval,starting from May 1,2016,related to the number of females in different breeding stages.(A)Towering rates(primary y-axis)observed in the afternoon(afternoon)and the forenoon(forenoon)related to the number of females(secondary y-axis)in the early breeding stages(farr,newly arrived;farr6,pre-nest-construction stage;PL,pre-laying stage;L,laying stage;PLL,either pre-laying or laying stage);(B)Towering rates(primary y-axis)observed in the afternoon(afternoon)and the forenoon(forenoon)related to the number of females(secondary y-axis)in the late breeding stages(I,incubation stage;N,nestling stage;F,fledgling stage).
3.Results
3.1.Towering and Moth flight description
Males by far most commonly delivered their songs from exposed perches,and rarely during horizontal Moth or vertical Towering songflights.Moth song-flight was observed prior to female arrival at the breeding grounds,while Towering was not.
A typical Towering song-flight is depicted in Fig.3A and is structured as follows(the corresponding vocalisation is given in brackets;see Towering song description below).A male calls from a perch and continues calling while it steeply ascends to an estimated 5–10 m above the take-off point(Introduction).At the peak of the ascending flight,the bird stalls shortly(Pre-rattle,Standard song,Post-rattle)before descending to a nearby perch,or occasionally,the starting point(Relax).The Towering song continues throughout all flight phases.While performing the Towering,the bird flies with shallow quivering wingbeats,lowered wings,raised tail,ruffled plumage and dangling legs.
Compared to the Towering flight,the Moth flight is much simpler.A bird typically flies in level flight from one song post to another,progressing more slowly than in normal flight with a similar body posture and wing action as in the Towering,and singing a single song strophe(Fig.3B).
3.2.Towering and Moth song description
Songs produced during Towering flights had a specific structure and could be divided into the following segments:Introduction,Pre-rattle,Standard song,Post-rattle and Relax(Fig.4).The Introduction comprised a medley of typical calls,i.e.,calls typically used in various other contexts(Fig.5),with pit and tseep being most frequent(Table 2).The Introduction lasted from 4 s to 5 min(Table 3).A few may have been even longer,as some Towering song-flights were not recorded from the very beginning.The Pre-rattle was typically present(in 93%of the cases),and was a succession of rattling syllables,usually of one type,but sometimes of two or three different rattle types.These rattle syllables(Fig.6),except the pook call,were specific to Towering songs as they were not recorded in any other context.The Standard song segment of Towering was a standard,i.e.,perch song strophe of a male or a somewhat shorter version with the last part absent.The Post-rattle was present in almost half of the cases(46%)and was constructed of the same types of rattle syllables as the Pre-rattle segment.The Relax segment,like the Introduction,was composed of typical calls but produced in a brief burst.Here,the pook call was the most frequent,the tseep was rarely used,and the teu was not recorded at all(Table 2).On four occasions,rattling was inserted into the Relax segment.Aside from the Standard song segment,Towerings of an individual male differed in duration and occurrence of calls.

Table 3 Duration(s)of Towering song-flight segments in male Black-headed Buntings(Emberiza melanocephala)from Croatia.
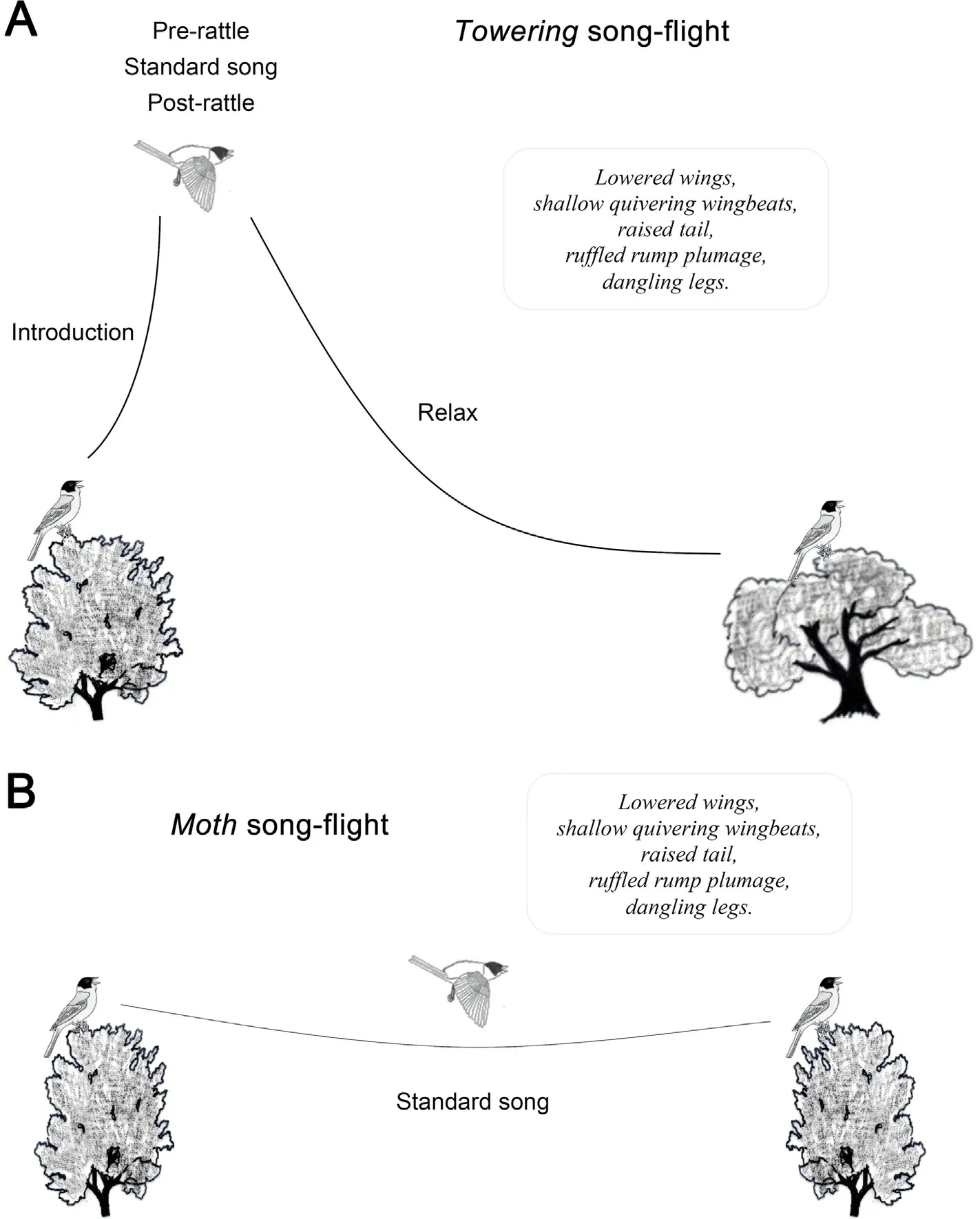
Fig.3.Schematic illustration of song-flights in male Black-headed Buntings(Emberiza melanocephala).(A)Towering song-flight,(B)Moth song-flight.

Fig.4.Sonograms of Towering song-flight of four Black-headed Bunting(Emberiza melanocephala)males from Croatia.Different segments of song-flight are indicated by frames;only the end of the Introduction is shown(512 FFT-length,Frame size=100%,Window=FlapTop,Temporal overlap=75%).

Fig.5.Sonograms of extracted typical calls in Black-headed Buntings(Emberiza melanocephala)from Croatia,with different versions of the same call type(512 FFTlength,Frame size=100%,Window=FlapTop,Temporal overlap=75%).

Table 2 The occurrence of different calls in the Introduction and the Relax segments of Towering song-flight in male Black-headed Buntings(Emberiza melanocephala)from Croatia.
The song produced during the Moth flight was the same as a songdelivered from a perch.Occasionally males produced silent Moth flights.
3.3.Towering song intra-and inter-male variability
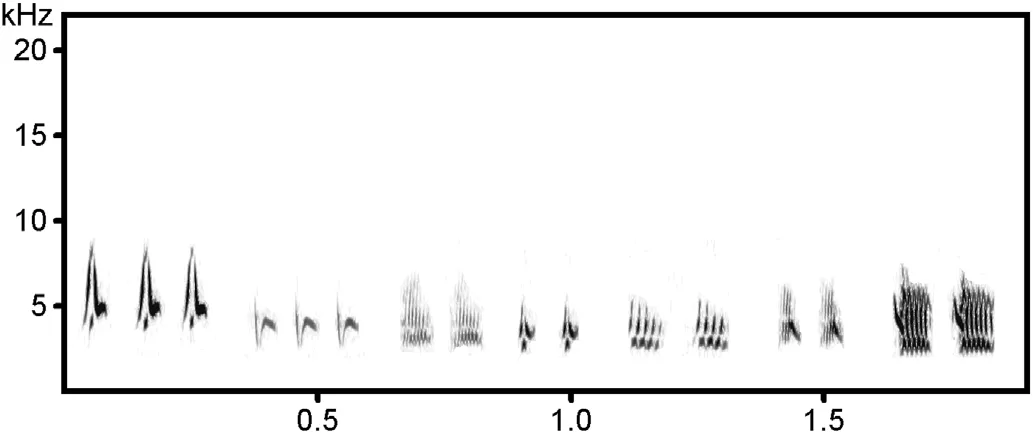
Fig.6.Sonogram of extracted different types of rattle syllables produced during the Towering song-flight by Black-headed Bunting(Emberiza melanocephala)males from Croatia.These types of rattle syllable were only recorded during Towering song-flights(512 FFT-length,Frame size=100%,Window=FlapTop,Temporal overlap=75%).
The variability in the number of syllables in the Relax segment of Towering song within the male that was recorded on five occasions over two years was higher than variability between the males(CVb/CVw=0.71).This individual,for example,had post-rattle and rattle syllables inserted into the Relax segment in one recording and not in the others.For other four males,the variability was slightly lower within than between the individuals(CVb/CVw=1.21,1.14,1.27,1.22).The results are inconclusive but suggest that the purpose of the Relax segment is not individual recognition.
3.4.Seasonal variation in song-flights rates
Even though Moth song-flights were observed immediately upon the males’arrival,they were infrequently seen during the standardised 20-min observation protocols throughout the season.Their frequencies did not show strong seasonal variations(in May 0.4±0.2 Moth flight/h,N=38 protocols;in June 0.2±0.06 Moth flight/h,N=162 protocols;Mann-Whitney U test,Z=0.6,p=0.5;Fig.1).Males were often observed performing Moth song-flights towards the loudspeaker broadcasting conspecific songs.Spontaneous Moth song-flights were all documented in the morning(N=27);during inter-male conflicts(41%),usually before chasing rivals or undertaking aerial fights,but also in other contexts including after engaging in duet-singing with neighbouring males(30%),while accompanying fledglings(11%),or while intruding into a neighbouring territory(7%).
Towering song-flights were more frequently detected than Moth songflights and were documented 59 times in 2011,59 times in 2012,73 times in 2013 and 111 times in 2016.The maximum observed Towerings per individual male was nine times in one breeding season.Males typically performed just one Towering per observation session.However,the maximum per session was four Towerings per individual male,performed approximately every 24 min.The minimum observed time between two successive Towerings by the same male was 5 min.
Towering song-flights were also infrequently seen during the standardised 20-min observation protocols,but their frequencies showed strong seasonal variations(in May 0.5±0.2 Towerings/h,N=38 protocols;in June 0.04±0.03 Towerings/h,N=162 protocols;Mann-Whitney U test,Z=3.6,p<0.001).The results were similar when considering all documented Towering song flights in the hearing range of a researcher,i.e.,the Towering rate dropped dramatically as the breeding season progressed(in May 0.6±0.08 Towering/h,N=47 observation sessions;in June 0.1±0.03 Towering/h,N=54 observation sessions;Mann-Whitney U test,Z=4.8,p<0.001;Fig.2).In May,the Towering rate was lowest at dawn and peaked in the late morning,while in the second part of the day it was highest in the late afternoon and decreased towards dusk(Fig.7).In June,the hourly pattern did not show strong variation,possibly due to a much smaller sample size.
3.5.Forenoon and afternoon towering rates in relation to female breeding stage
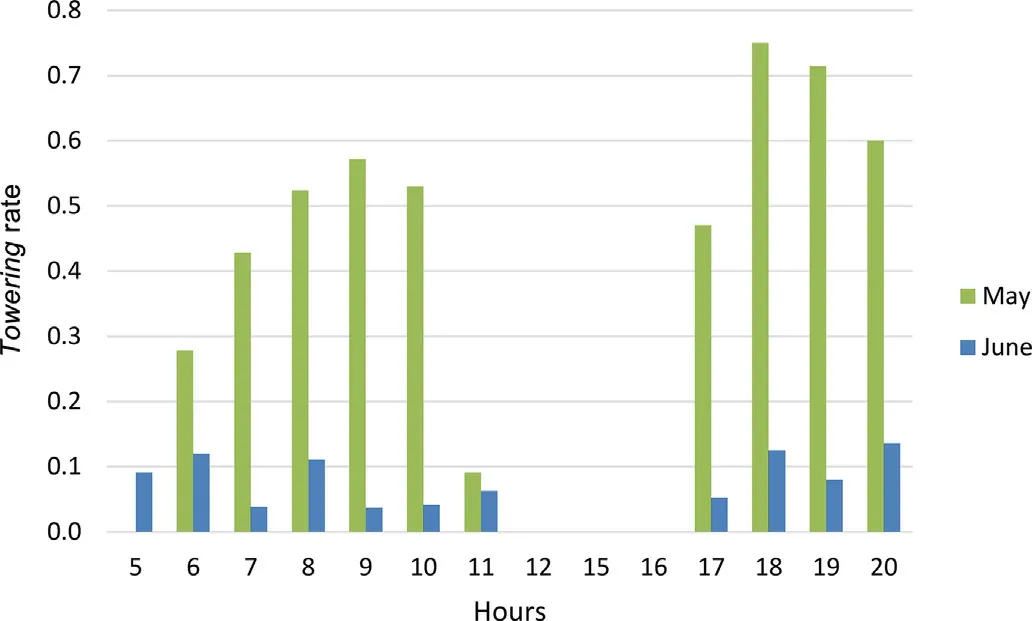
Fig.7.The diurnal pattern in Towering song-flight rate(Towerings/h)in male Black-headed Buntings(Emberiza melanocephala)from Croatia in May and June 2016.The period from 13:00 to 15:00 was not sampled.
Following model selection,the best models explaining variation in male Towering rates revealed that forenoon Towering rates were positively related to the number of females in the pre-laying stage,whereas afternoon Towering rates were positively related to the number of females in the pre-nest construction stage(Fig.8;Table 4).Compared to other models,these models had strong support,with an Akaike weight of 0.86 for the forenoon Towering rate and 0.99 for the afternoon Towering rate.There was no indication of the autocorrelation of residuals(Box-Ljung test for best models;forenoon:p=0.99,afternoon:p=0.23).
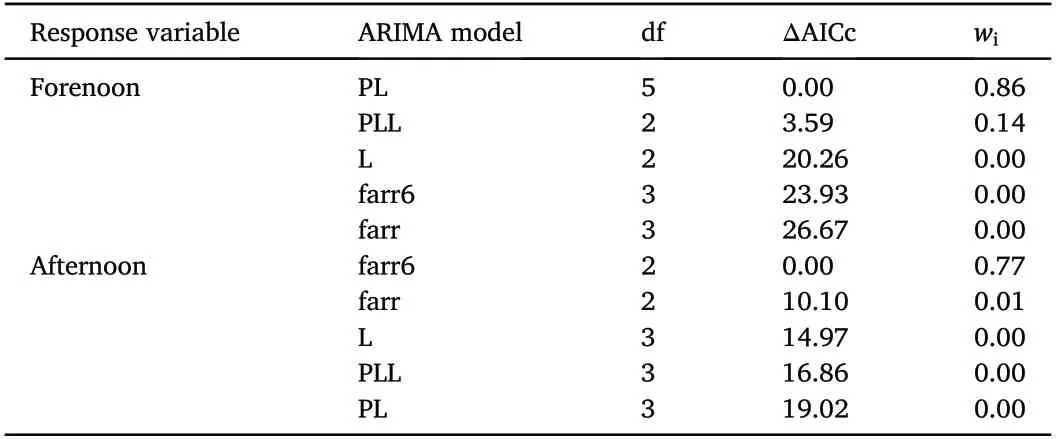
Table 4 Summary of model selection results for the forenoon and afternoon Towering rates of Black-headed Buntings(Emberiza melanocephala)from Croatia in 2016.

Table 5 Predictors estimates,standard errors(SE),z and p values of forenoon and afternoon Towering rates of male Black-headed Buntings(Emberiza melanocephala)from Croatia in 2016 from the best models(see Table 4).
To account for a possible year effect,the number of observed Towerings per three-day period was compared to the number of females in the pre-laying stage for years 2011–2013.The number of females in the prelaying stage was significantly positively correlated with the forenoon Towering occurrence for each year(Spearman correlation:2011,r=0.69,p=0.002,N=18;2012,r=0.52,p=0.03,N=18;2013,r=0.60,p=0.004,N=21)but not with the afternoon Towering occurrence for either year(Spearman correlation:2011,r=0.17,p=0.5,N=18;2012,r=0.36,p=0.1,N=18;2013,r=0.36,p=0.1,N=21).
4.Discussion
This study confirmed the existence of two types of song-flight display in male Black-headed Buntings:Moth and Towering song-flights.The Moth song-flight was relatively simple,incorporating level flight and perch song.The Towering song-flight had both elaborate vocal structure and locomotor behaviour.Towerings were more frequently detected than Moth song-flights,but detection probabilities varied considerably between the two since Towerings could be detected by sound,while Moth song-flights could only be identified visually.To our knowledge,this is the first study of the Towering song-flight of the Black-headed Bunting with a detailed description and sonogram representation.Song-flight behaviour exists in some other Emberiza buntings.Similar displays have been visually described for the Red-headed Bunting,Emberiza bruniceps,a closely related species with a similar breeding system(Cramp and Perrins,1994).The Black-headed Bunting's Towering song-flight structure is also similar to the song-flight structure of the Ortolan Bunting(Emberiza hortulana;Lang,2011).It also has the initiation and relaxation segments comprising typical calls and rattles before and after the Standard song.Apart from the Standard song segment,each Towering vocal performance of an individual Black-headed Bunting male was different,leading to the conclusion that Towering song is stereotyped in neither duration nor call type occurrence.The same is true for the Ortolan Bunting's song-flight(Lang,2011).
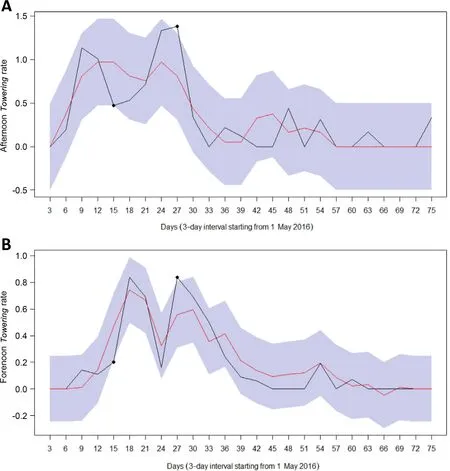
Fig.8.Observed(black line)and predicted(red line)afternoon(A)and forenoon(B)Towering rates(Towerings/h)in male Black-headed Buntings Emberiza melanocephala from Croatia in 2016 based on the best model parameter estimates in Table 5.Black dots represent values of the response variable outside the modelled 95%confidence intervals(grey polygons).
It has been shown in many bird species that song is important both in territory defence and mate attraction.This distinction is especially pronounced in bird species that have songs of different types(Catchpole and Slater,2008).Black-headed Bunting males sang immediately upon arrival,well before female arrival,indicating that perch song,at least in this period,is directed towards other males(Bariˇsi′c et al.,2018).Perch song repertoire in the Black-headed Bunting is constrained to only one song strophe,which is individually distinct and remains constant throughout the male's lifespan,and thus is likely used in individual recognition of males(Bariˇsi′c et al.,2018).Several neighbouring males typically share the same song type,and the similarity in the perched song is likely to be selected through inter-male competition(Bariˇsi′c et al.,2018).Males reacted to the playback of conspecific songs either by singing from perch or by approaching the loudspeaker with Moth song-flight.This is further evidence that perch song is used in territory defence and indicates that Moth song-flight serves as a threat display,especially as males usually exhibit spontaneous Moth song-flights before engaging in chasing rivals or undertaking aerial fights.Our study showed that the primary function of Towering song-flight is mate attraction.A song produced during Towering is more elaborate and variable than a perch song,thereby allowing an assessment of the male's song-flight repertoire.Compared to the perch song,which always stayed the same and could therefore be used in individual recognition,the Towering song incorporated both the constant part,i.e.,the perch song and the variable part,i.e.,the Relax segment.By adding the Relax segment to the perch song,a male can perhaps simultaneously demonstrate its identity and quality.The Towering does not start before females arrive,and its performance rate is highly related to the presence of receptive females.The Towering song-flight of the Black-headed Bunting is a multimodal signal integrating acoustic(song),motor(flight)and possibly visual(bright ruffled plumage)components.To perform Towering flight,a Black-headed Bunting male needs to ascend steeply into the air.Song-flights could be physically demanding(Usherwood,2008)and/or energetically costly(Hedenstrom and Møller,1992),thus allowing precise estimation of a male's condition and abilities.Furthermore,the relative infrequent occurrence of Towering displays might indicate that it imposes a high cost on performing males either in terms of energetic demands,physical exhaustion,or greater conspicuousness to predators.
In many bird species,dawn choruses are directed toward males and serve in territory defence while singing at other times of the day appears to play a role in attracting mates(Klit,1999;Liu et al.,2007;Moran et al.,2019).The Black-headed Bunting perch song peaked at dawn and gradually decreased into the late morning(authors’unpublished data).In contrast,the Towering rate was lowest at dawn and in the second part of the day also decreased towards dusk.Males might avoid performing Towering song-flight in low visibility conditions that would impair the reliable transmission of the intended message.Additionally,the Towering rate could be constrained by the demands of other behaviours,such as perched singing that is used in territory defence.
The Towering rate dramatically dropped as the breeding season progressed.Furthermore,there was a clear difference in peak Towering rates between forenoon and afternoon,suggesting that forenoon and afternoon Towerings have different functions or convey different messages.Our findings from the 2011–2013 dataset supported the same conclusion,thus excluding a year effect.
The female fertile period depends on the sperm storage duration(Birkhead and Møller,1993a)and is in most studies defined as starting a few days before laying the first egg and universally ending the day the penultimate egg is laid(Birkhead and Møller,1992;Kempenaers et al.,1995;Moran et al.,2019).Female fertility increases as egg-laying approaches(Birkhead,1998).However,once the egg-laying starts(or usually the day before),female fertility is somewhat hindered,as the passage of sperm up the female reproductive tract is to some extent blocked by the developing egg(Birkhead and Møller,1993b).In most passerine species,copulation peaks in the morning(Birkhead et al.,1987;Sheldon and Burke,1994).The closed habitats and secretive behaviour of Black-headed Bunting females meant that we were only able to observe copulation events on two occasions:at 06:55 and 09:45;both females were in the pre-laying stage.Assuming that copulatory behaviour in the Black-headed Bunting peaks in the morning and given the strong relationship between forenoon Towering rate and the period of high female fertility,forenoon Towering displays could be directly linked to copulation events and are likely to be timed to coincide with high fertilisation probabilities.To our knowledge,the reasons for the variation in diurnal timing of copulation are unknown,so we can only speculate about the underlying reasons.For example,since it has been shown in some species that sperm depletion in males can occur after only a few copulations(Birkhead,1991;Westneat et al.,1998),females may solicit copulation in the morning to secure ejaculates with a high sperm count.
Afternoon Towering peaked very early in the breeding season and coincided with the number of females present at the breeding site before the onset of the nest-building activity.As this is unlikely a period of high female fertility(fertility rises toward the start of egg-laying)it can be argued that afternoon Towerings are not related to fertilisation likelihood but serve to attract social mates.Song-flight performance depends on wingspan,wing area and body mass(Hedenstrom and Møller,1992).Although wingspan and wing area stay more or less the same throughout the breeding season,the bird's body mass has strong seasonal and diurnal variation(Blem,1976;Haftorn,1989;Witter and Cuthill,1993).Flight cost in birds increases allometrically with body mass(Pennycuick,1989).Performing Towering presumably becomes more demanding as the day progresses and the bird accumulates body fat reserves.Thus,a song-flight could be regarded as a condition-dependent sexual signal as its cost changes depending on the physical condition of the performing male.If the signal is more costly at a particular time,it is also likely to be more honest(Hedenstrom and Møller,1992).It may be that only high-quality males can perform well with some extra weight or can afford to understock their fat reserves due to superior foraging abilities.In conclusion,female Black-headed Buntings may prefer to choose their social mates based on the afternoon Towering performance as a more reliable signal of male quality.Condition-dependent traits are considered important in sexual selection,as they can help a female assess a male's quality in a refined way(Byers et al.,2010;Jacobs and Zuk,2010).
It was proposed in the Whitethroat(Sylvia communis)that the purpose of song-flight is to gain a better view of the surroundings(Balsby and Dabelsteen,2003).In the Ortolan Bunting,it is thought that song-flight serves as the‘last warning’to females and offspring about an immediate threat from predators(Lang,2011).It is unlikely that the sole purpose of an elaborated and‘overemphasised’Towering song-flight in the Black-headed Bunting is to warn females and offspring or to gain a better view of surroundings as this can be done in a more efficient and less physically demanding way.Furthermore,a protection function of nest and offspring is unlikely,given that there is no relationship between the Towering rate and the number of nests in the incubation or nestling stage.
5.Conclusions
Our data provide evidence that birds can direct different types of song-flight towards different receivers.Towering song-flight is directed towards the female,whereas Moth song-flight is used in male-male interactions.Furthermore,the time of the day seems to be a determinant whether Towering is used in social pairing or to increase fertilisation probabilities.A displaying male could be constrained by either energetic demands or by physical abilities,and a female could base her choice on how often or how well a male display.Future work is needed to account for the effect of individual male variability in Towering displays on within-pair and extra-pair mating success.Furthermore,a question that remains largely unanswered is which parameters,e.g.,rate,height or trajectory of flight or even song quality,females use in their choice of mates.In contrast to the straightforward measurement of Towering frequency,measuring the quality of Towerings(height,steepness)would be a challenging endeavour due to its rarity and unpredictability and the difficulty in observing a displaying male in dense habitat.
Authors’contribution
SB conceived the study.SB,D′C,VT,JK and HH carried out the fieldwork and collected the field data.HH supervised the study.SB analysed and interpreted the data and wrote the first draft of the manuscript.SB,D′C,VT,JK and HH reviewed and edited the manuscript.All authors read and approved the final manuscript.
Ethics statement
This study followed the institutional and national ethical guidelines for scientific research.Bird capture and ringing were performed under a ringing licence(issued by the Institute of Ornithology,Croatian Academy of Sciences and Arts and authorised by the Ministry of Environment and Energy).The study was approved by the Croatian Ministry of Nature Protection(UP/I-612-07/:11–33/0571,12–33/0536,13–48/53,14–48/24,15–48/86,16–48/17).
Declaration of competing interest
The authors declare that they have no competing interests.
Acknowledgements
We thank all the inhabitants of our study area who kindly welcomed us and allowed us to work on their land.We are most grateful to the family of Zdravko and Anka Mui′c for their hospitality and support in the fieldwork.We are indebted to Marko Bariˇsi′c for his help with the fieldwork and especially for making high-quality sound recordings and producing a schematic illustration of song-flight in the Black-headed Bunting.We thank Michael Schaub for his crucial help with statistical analysis,along with constructive comments and improvements to the manuscript.Finally,we would like to thank the editor and the reviewers for their thoughtful comments and efforts in improving the quality of the manuscript.This work was partially funded by the Croatian Environmental Protection and Energy Efficiency Fund.
杂志排行

Avian Research的其它文章
- Use of 3D-printed animal models as a standard method to test avian behavioral responses toward nest intruders in the studies of avian brood parasitism
- Habitat selection and food choice of White-naped Cranes(Grus vipio)at stopover sites based on satellite tracking and stable isotope analysis
- Year-round multi-scale habitat selection by Crested Tit(Lophophanes cristatus)in lowland mixed forests(northern Italy)
- Definition of spatial positions within foraging Greater White-fronted Geese flocks from an individual's perspective:Cost-benefit dynamics change with the distance to flock edge
- Urban avifauna distribution explained by road noise in an Eastern European city
- Demographic and behavioral responses of the White-winged Dove to human disturbances during winter in northwestern Mexico