Definition of spatial positions within foraging Greater White-fronted Geese flocks from an individual's perspective:Cost-benefit dynamics change with the distance to flock edge
2023-01-03MengzhenWangWenwenChenChunlinLiJinmingZhao
Mengzhen Wang,Wenwen Chen,Chunlin Li,Jinming Zhao
a School of Life Science,Huaibei Normal University,Huaibei,235000,China
b School of Resources and Environmental Engineering,Anhui University,Hefei,230601,China
Keywords:Distance to edge Foraging time Intake rate Spatial position within a group Vigilance time
ABSTRACT The costs and benefits of group living are not evenly distributed among individuals that occupy different spatial positions within a group(central or peripheral).However,the definitions of central and peripheral positions may artificially mask the true dynamics of cost and benefit of individuals.Using wintering foraging Greater Whitefronted Geese(Anser albifrons)flocks as our study system,we measured individual spatial position within a flock as distance to the flock edge.First,we tested the effects of distance to edge on the antipredation cost and foraging benefit of individual goose using generalized linear mixed models(GLMMs).Flock size and number of neighbors and their interactions with distance to edge were included as fixed effects in GLMMs.Then we used piecewise regression to determine the threshold value of distance to edge to classify central and peripheral individuals.Our results indicated distance to edge significantly affected percent of time spent on vigilance(vigilance time),percent of time spent on foraging(foraging time)and intake rate(pecks/min),whereas other factors had no effects.We found that the critical distance that predicted changes in behavior was around 2.5 m.Birds 2.5 m or less from the edge of the group were more vigilant,and had less efficient foraging than more central individuals.Behavioral differences between central and peripheral individuals were more easily detected when the birds were classified by the method introduced in this study than by the traditional minimum convex polygon method.Our research provides researchers a dichotomous classification scheme that places individuals into peripheral and central positions from an individual's perspective.
1.Introduction
Understanding the evolution of group living is a long standing question in ethology(Ioannou,2017a).Animals join in groups to acquire benefits such as food acquisition,predator avoidance,mate choice,energy saving,travel efficiency and information sharing(Kopachena,1987;Romey and Emily,2008;Killen et al.,2011;Ioannou,2017b;Gil et al.,2018),but incur the costs of competition with conspecifics(Sansom et al.,2008;Hirsch,2011b;Zhao et al.,2020).Benefits acquired through group living are not shared homogeneously by all group members due to phenotypic variation in age,sex,hierarchy,satiety and body size,among other factors(Stankowich,2003).Even within groups composed of individuals of very similar phenotype,however,benefits acquired by group members still vary with spatial position within a group(Krause,1994).Individuals on the periphery of a group may suffer a higher predation risk than those in central positions(marginal predation)because they are more exposed to predators or merely because they are closer to predators(Duffield and Ioannou,2017),therefore,peripheral individuals often devote more time to vigilance directed to detect potential predators(edge effect;Fern′andez-Juricic and Beauchamp,2008).Furthermore,central individuals may have a lower food intake rate because the abundance of food resources may be decreased after the consumption or disturbance of the peripheral individuals(Hirsch,2007a;Morrella and Romey,2008).A central-edge effect is likely to emerge when animals occupy central or peripheral positions within a group according to the tradeoffs between the benefits and costs associated with these positions(Quinn and Cresswell,2006;Hirsch,2007b).
The presence of a central-edge effect on the benefits and costs accrued by individuals occupying different spatial positions within a group has well been established in a variety of studies on animal groups(Krause and Ruxton,2002;Christman and Lewis,2005).However,a general problem in existing studies is that researchers have generally categorized central and peripheral positions arbitrarily and from a human's perspective.Stankowich(2003)summarized four methods to categorize central and peripheral individuals within a group:(1)polygon vertices or minimum convex polygon method(MCM);(2)layers of protection;(3)concentric circle;and(4)mean distance to center,with the MCM as the commonly accepted and most widely used method by researchers.In the MCM method,the smallest closed convex polygon is drawn by connecting all of the individuals(considered to fall within a group)that are furthest from the center of the group.Those individuals that fall at the vertices of the convex polygon are considered to be peripheral,and all others are considered central.However,for some group shapes,animals considered‘central’can fall very close to the peripheral lines or vertices.The classification of central in this case may be too arbitrary and incorrect because these individuals may actually face a much higher risk of predation than those far from the edges.Some researchers defined spatial positions based on their own experience.For example,some cutoff values describing individuals'distances from the outmost edges of groups were set by investigators to dichotomize central and peripheral positions,and these values were varying based on the researchers'experience.Categorizing central and peripheral positions in an inappropriate way may be too artificial in some cases and contribute to a misunderstanding of the true cost-benefit dynamics along the peripheral-center gradient(Stankowich,2003).
A more promising methodology is to measure spatial position within a group as a continuous variable and explore the ways in which costs and benefits change along a gradient of group positions(Stankowich,2003).There are two methods to continuously quantify individual spatial position within a group.One method is to measure the distance between an individual and the geometrical center of a group.The other method is to mark an individual's spatial position according to its distance to the outmost peripheral of the minimum convex polygon encompassing all group members.The former method requires an overhead(aerial)picture of the group(to get the positions of all individuals in a group)and time-consuming calculations to determine the center of a group,which is not methodologically possible for many field studies.Furthermore,it will become harder when dealing with moving groups than stationary groups(Krause,1994).In contrast,applying the latter method is easier(no need to know the positions of all group member but the outmost individuals or lines between them in a local scale)and more appropriate to test the hypotheses related to individual spatial position within a group such as the marginal predation.
In this study,we used wintering Greater White-fronted Geese(Anser albifrons)flocks to examine the relationship between cost-benefit dynamics of individuals and the continuous distance from the periphery to the center of flocks.First,we tested whether an individual's spatial position within a group would influence its costs and benefits received through joining groups.We used the percent of time spent on vigilance(vigilance time)as a surrogate of predation risk because researchers have found that animals'vigilance time is highly related to their perceived predation risk(Saout et al.,2015;Beauchamp,2017).We used time spent on foraging and intake rate to calculate the benefits obtained by animals.We measured individual distance to the edge of the minimum polygon(a continuous variable)as the spatial position of an individual within a group.We also incorporated flock size and number of neighbors and their interactions with individuals'spatial position when we test the effect of distance from the flock edge.Flock size has been found to be one of the most important flock characteristics affecting the benefits and costs received by individuals(Beauchamp,2008;Sansom et al.,2008).And previous studies found number of neighbors(within a 5-m quadrat centered by the focal individual)significantly influences individuals'vigilance and foraging behaviors(Zhao et al.,2019,2020).Then we tried to dichotomized central and peripheral individuals based on a cutoff value of distance to edge determined by abrupt changes of cost paid and benefits received.Finally,we compared the behavioral differences between the two groups of central individuals and between the two groups of peripheral individuals classified by the cost-benefit dynamic methods(CDMs)and the MCMs,respectively.
2.Methods
2.1.Study area and species
We conducted our study at Babaizhang,the lower part of Shengjin Lake(30°15′–30°30′N,116°55′–117°15′E;Fig.1),a shallow freshwater lake connected with the Yangtze River through the Huangpen sluice.The open water area of the lake shrinks from approximately 140 km2in summer to about 34 km2in winter as the water level fluctuates seasonally.The annual rainfall is about 1600 mm at this site.The mean temperature is about 6°C in December when the study was conducted.Detailed descriptions about the study area were presented in Zhao et al.(2013,2015).
Wintering Greater White-fronted Geese migrate to the shallow lakes along the middle and lower reaches of the Yangtze River flood plain from October to November and depart for breeding grounds from March to April each year.Although wintering Greater White-fronted Geese forage mainly in abandoned fields in Europe and North America(Fox et al.,2016),they mainly forage in natural habitats and consume sedges in China(Carex spp.;Zhao et al.,2018).Sedges are the dominant vegetation and are distributed almost evenly along the littoral zones of many shallow lakes in the Yangtze River flood plain,including the Shengjin Lake.Geese flocks often change their habitat use along the littoral zones,according to the depletion and renewal of the local sedges.Flock sizes change daily or more frequently as geese move about.There are no large carnivores that can prey upon wintering geese at Shengjin Lake.However,some raptors such as Peregrine(Falco peregrinus)and Bonelli's Hawk Eagle(Hieraaetus fasciatus)have been found to prey upon geese.In addition,human activities(illegal fishing,fishery inspection,grazing,agriculture activities along the lake shores)and disturbance by domestic dogs often trigger a flight reaction in geese flocks.
Shengjin Lake suffered a severe drought in 2019,the water level decreased to low levels a month earlier than ordinary years.This led to the overgrowth of sedges,which was unsuitable for the migrating geese to feed on when they arrived in November.The total number of geese at Shengjin Lake decreased from more than 13,000 in December 2018 to no more than 1500 individuals in December 2019.Most of the geese abandoned the former foraging sites(Zhao et al.,2019)at the upper part of the lake and were found to forage on a small island at Babaizhang in 2019 when this study conducted.No more than 200 individuals were occasionally found to forage at the traditional wintering sites at the upper part of Shingjin Lake.
2.2.Data collection and digitization
We recorded the behaviors of geese with the aid of a camera(Coolpix P1000,Nikon Corp)between 7:00–11:30 and 13:00–17:00,when these geese were foraging extensively,from December 3 to December 30,2019.We collected data on sunny days with clear field of view from a dike 10 m above the ground.Individuals were considered as belonging to the same group when they were separated by less than 20 m.
Before recording the behavior of geese,we firstly estimated the flock size using increments of 1,10,50,100 and 500 individuals according to a preliminary estimation of flock size(Zhao et al.,2019).Then we randomly selected one individual from one end of the flock and recorded its spatial position as the distance to the edges of the minimum polygon.Distance to edge was estimated by the bird lengths(0.7 m/bird)or based on the wreckages on the sedge meadows(Zhao et al.,2020).The spatial positions of individuals at the vertices of the polygons were recorded as 0 m.We videoed the behaviors of selected individual for 5 min and then recorded the spatial positions again.The mean values of the two spatial positions were used in the following analyses.We selected another individual that was about 20 m from the first one and recorded its spatial position and behaviors using the methods described above.We continued to select focal individuals until we reached the other end of a flock.This procedure enabled us to minimize the risk of recording the behaviors of the same individual.We kept the focal individual at the center of the camera lens and included the nearby neighbors in the lens coverage during the videoing process.
Fig.1.Location of Shengjin Lake and the study site.
The same flock was only sampled once in a day.We did not collect behavior data when the flock contained>5%of other geese species.We stopped recording behavior data when the flock size changed during an observation period and excluded these data from subsequent analyses(n=9).To control for the confounding effect of age,we collected data only from adult geese,which were distinguished from juveniles based on the white patch on forehead and the mottled/scaly plumage pattern on flanks and breast(Koffijberg,2006).
Back in the laboratory,we digitized the behavior data of focal individuals.For each observation period,we defined the following behaviors of geese:vigilance(neck stretched vertically with its head horizontal and eyes scanning the surroundings),foraging(sudden,short range movement of the beak toward the ground),competition(chasing or threating or attacking between a focal individual and other companions with their neck stretched horizontally),drinking,resting(crouching on the ground or standing with its head under the wing),and preening(Kotrschal et al.,1993).We recorded vigilance time(quantified as the percentage of total time spent on vigilance),foraging time(quantified as the percentage of total time spent on foraging)and intake rate(quantified as the mean number of pecks per min during an observation session).
We defined neighbors as other individuals within a 5-m quadrat centered at the focal individual from the video(Zhao et al.,2020).We estimated the 5 m distance as 7 body lengths from the left-right dimension,and estimated the 5 m front-rear distance based on preliminary observations with high accuracy(Zhao et al.,2020).We recorded the number of neighbors within the quadrat at 0,1,2,3,4,and 5 min,and used the mean value for further analyses.
2.3.Statistical analysis
We examined the effects of flock size,number of neighbors and spatial position within a flock on vigilance time,foraging time and intake rate separately using generalized linear mixed models(GLMMs)fitted by maximum likelihood(Favreau et al.,2010),with flock identity as a random variable.GLMMs have been proved to be reliable for analyzing non-normal data with random effects(Bolker et al.,2008).We also included the interaction terms of flock size with spatial position within a flock and number of neighbors with spatial position within a flock.Therefore,we had five variables as the fixed effects in GLMMs.We used a Gaussian distribution with an identity link function when analyzing variables influencing vigilance time,foraging time and intake rate.Vigilance time was square root transformed while foraging time was inverse sine transformed to meet the normal distribution.GLMMs were performed using the“lmerTest”package in R 3.6.2(www.r-project.org).Flock size was log10transformed before analyses.
We used locally weighted regressions(LOESSs)to explore the relationship between spatial position within a flock and vigilance time,foraging time and intake rate(Cleveland,1979).The advantage of this regression is that it does not require the specification of a function to fit a model to all of the data in the sample,and thus to examine the correlation between the value changes of cost and benefit with the distance to flock edge(Pujol et al.,2018).LOESSs were conducted in“ggplot2”package in R 3.6.2.Based on the results obtained from the LOESS models,we set 3.5 m to the flock edge as the threshold value to classify individuals’spatial positions.The value of 3.5 m was roughly estimated by eyes according to the changes of cost and benefit as the distance to flock edge varied.Then we used the piecewise regression to refine the threshold values in“segmented”package in R 3.6.2.
Based on the results obtained using the above methods(see Results),we set 2.5 m to the flock edge as the threshold value to classify individuals’spatial positions as central and peripheral.Then we compared the vigilance and foraging behaviors of central and peripheral individuals classified by our method(cost-benefit dynamic method,CDM)and the traditional MCM using one-way analysis of variance(ANOVA).Between group differences were examined by computing Tukey honest significant differences(TurkeyHSD)in“multcomp”package.
Some individuals changed their spatial positions at the end of the observation sessions.We examined the behavior change of these individuals between their first halves(2.5 min)and the second halves(2.5 min)of the observation sessions using Wilcoxon rank test.All results were presented as means±SE.Two-tailed statistical significance was set to 0.05.
3.Results
In total,we videotaped 457 individual geese from 88 different flocks.The mean values of flock size and number of neighbors were 432±74(range:3 to 4400)and 3.5±0.1(range:0 to 15.2).
With flock identity as a random factor,general linear mixed models indicated that increasing distance to edge positively affected foraging time and intake rate,while negatively affected vigilance time(Table 1).Flock size and number of neighbors and their interactions with distance to edge did not have significant effects.Further analysis revealed that instantaneous intake rate(the mean number of pecks observed across 5 min,1 min intervals)was not influenced by flock size(-2.015±1.258,z=-1.602,P=0.109),number of neighbors(-0.011±0.215,z=-0.049,P=0.961)or distance to edge(-0.053±0.075,z=-0.699,P=0.484).
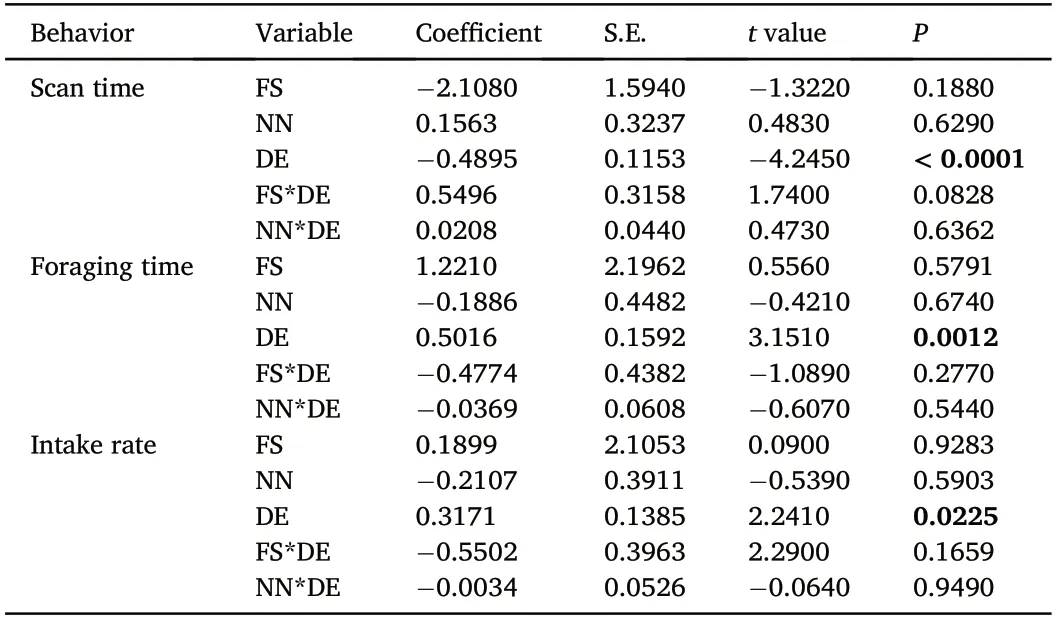
Table 1 Fixed effects of flock size,number of neighbors,and position within a flock on vigilance and foraging behaviors of individual Greater White-fronted Goose,analyzed using generalized linear mixed models,with the effects of flock identity controlled.
Vigilance time decreased dramatically as distance to edge increased from 0 to 3.5 m and levelled off thereafter(Fig.2A).Intake rate and foraging time increased sharply as distance to edge increased from 0 to 3.5 m,with no obvious change when distance to edge exceed 3.5 m(Fig.2B and C).Using piecewise regressions,we found that the refined turn point values(or threshold values)were 2.43 m for vigilance time,2.57 m for foraging time and 2.59 m for intake rate.As the three refined values were similar,we examined the correlations between the vigilance time,foraging time and intake rate using spearman correlation test.Foraging time and vigilance time were negatively correlated(r=-0.75,P<0.001),foraging time and intake rate were positively correlated(r=0.86,P<0.001).
We used a mean value of 2.5 m as the threshold value to classify central and peripheral individuals.There were significant differences in vigilance time(15.97±0.75 vs 25.51±1.66,P<0.001),foraging time(75.41±1.19 vs 64.75±2.00,P<0.001)and intake rate(58.57±1.10 vs 50.72±1.67,P<0.001)between central(n=296)and peripheral(n=161)individuals classified by CDMs(Fig.3).In contrast,there were significant differences in vigilance time(18.40±0.78 vs 28.07±3.38,P=0.0012)and foraging time(72.64±1.09 vs 62.41±4.09,P=0.022),but no difference in intake rate(56.51±0.97 vs 49.20±3.47,P=0.095)between central(n=413)and peripheral(n=44)individuals classifiedby MCMs.Moreover,there were no differences in vigilance time,foraging time and intake rate between central individuals(all P>0.05)classified by CDMs and MCMs,and no differences were found between peripheral individuals(all P>0.05).
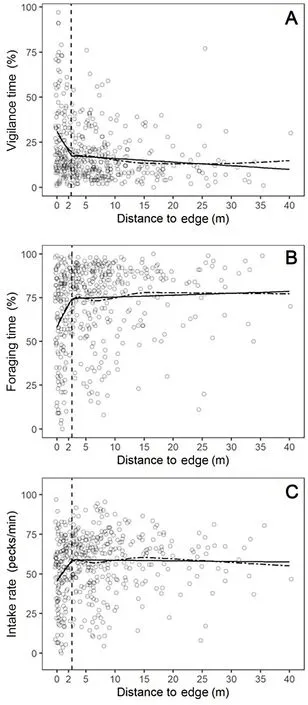
Fig.2.The changes of(A)vigilance time,(B)foraging time and(C)intake rate of individual Greater White-fronted Goose with the distance to edge.Dashed lines:fitted by LOESS models,straight line:fitted by piecewise regressions.
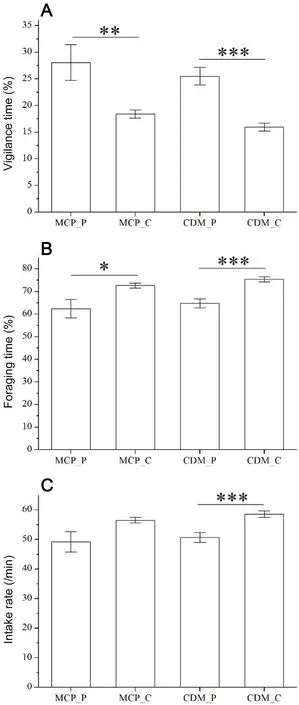
Fig.3.The behavioral difference between peripheral and central individuals classified by the traditional minimum convex polygon method(MCM)and the cost-benefit dynamic method(CDM)used in this study.*:0.01≤P<0.05;**:0.001≤P<0.01;***:P<0.001.
Forty-one geese that initially occupied peripheral positions moved to central positions at the end of the observation sessions and 24 geese moved from central to peripheral positions classified by CDMs.There were no changes in vigilance time(0.17±0.02 vs 0.14±0.02,P=0.193),foraging time(0.82±0.02 vs 0.83±0.02,P=0.268)and intake rate(64.26±0.45 vs 64.49±0.24,P=0.644)when geese moved from peripheral to central positions.As for geese that moved from central to peripheral positions,intake rate(60.34±3.55 vs 50.02±5.05,P=0.044)decreased and vigilance time(0.21±0.04 vs 0.21±0.04,P=0.974)and foraging time(0.74±0.04 vs 0.63±0.06,P=0.067)did not differ between the first and second halves of the observation sessions.
4.Discussion
Our results suggest that geese near the flock edges paid the cost of heightened vigilance,but that was not counterbalanced by foraging benefits.More importantly,there was an abrupt change of cost paid and benefit gained as the distances of individuals from the edge increased from 0 m to ca.2.5 m,providing researchers a dichotomous classification scheme that places individuals into peripheral and central positions from an individual's perspective than the traditional methods that based on researchers'subjective experiences or arbitrary choices.Moreover,behavioral differences between central and peripheral individuals were more easily detected when spatial positions were classified by the costbenefit dynamic method.
Animals that occupy the peripheral positions of groups are often more exposed to predators and have a higher risk of predation than those in center positions(Inglis and Lazarus,1981).These edge effects have been found in many animal groups where peripheral individuals devoted more time to vigilance than central individuals(Matson et al.,2005;Dias,2006;Shi et al.,2011;Beauchamp,2015).The higher vigilance levels of individuals near the flock edges found in this study is consistent with the predictions of the edge effect,and most other studies on the costs and benefits of animal group dynamics.
Although peripheral individuals pay greater antipredator costs than central ones,some studies have suggested that these costs could be counterbalanced by foraging benefits,implying a trade-off between predation risk and foraging efficiency(Morrella and Romey,2008;Hirsch,2011b;Hansen et al.,2016).Contrary to the findings of these studies,we found that individuals at peripheral positions spend less time on foraging and had a lower intake rate than those in central positions,indicating individuals did not gain a higher foraging benefit by occupying peripheral positions.Sedges were generally evenly distributed along the littoral zones and might be abundant enough to preclude peripheral geese to deplete these food resources(Hall and Fedigan,1997;Hirsch,2011a).A previous study at the same study system also found that peripheral geese had a lower intake rate than central ones(Zhao et al.,2020).Peripheral individuals might be poorer competitors that spend more time monitoring not only potential predators but also nearby neighbors.Our data preclude us to test this assumption because only 21 birds(unpublished data)were observed to have agnostic competition behaviors in the 457 individuals observed in this study.Another method to verify this assumption is that capturing and marking some geese to examine whether an individual’spatial position within a flock was correlated with its social status.Moreover,detailed studies should also be conducted to examine whether peripheral individuals could gain foraging benefits by taking higher quality(more nutritional)parts of the sedges.
The most important finding of our study is that there was a clear spatial boundary beyond which there was no obvious changes in costs paid and benefits gained for individual geese.The outmost peripheral individuals paid the largest antipredator vigilance cost and gained the lowest food intake benefit.Initially,the cost decreased and benefits increased as the individual distance to flock edge increased from 0 to 2.5 m,but levelled off thereafter.Our results agree with Stankowich's(2003)suggestion that,to some extent,individual spatial position within a group should be considered to be a continuum.Arbitrary classifications used in many other studies might have masked the true cost-benefit dynamics when individuals choose different spatial positions(Christman and Lewis,2005;Hirsch and Morrell,2011).This was confirmed by our study which revealed more obvious behavioral differences between central and peripheral individuals classified by the CDMs.When researchers try to compare risk of predation between peripheral and central individuals using the“polygon vertices”,they might categorize individuals near flock edges who face a similar predation risk to outmost individuals as central ones that might have a much lower risk of predation.Similarly,researchers might wrongly categorize safer central individuals as peripheral ones using“concentric circles”.We recommend researchers to use a continuous measure of individual spatial positions within a group and categorize central and peripheral individuals based on the cost and benefit change from an animal's perspective when necessary.For example,researchers could dichotomize individuals to central and peripheral ones using the CDMs instead of categorizing them based on the MCMs.Furthermore,the estimated threshold values might show big differences according to the variables used,which might be different from the results of this study where we found similar values.
It was not surprising that flock size did not influence the vigilance and foraging behaviors of geese because flock sizes in our study were larger than most of other studies that compared individuals’behavior performance in relatively small groups or between solitary and aggregated individuals(Beauchamp,1998).Animals might not be able to estimate the number of conspecifics and adjust their behaviors accordingly when group size was beyond a certain value(Emmerton and Delius,1993).However,individuals joining large aggregations can still gain an antipredator benefit passively through risk of dilution and confusion effect(Landeau and Terborgh,1986;Santos et al.,2016;Hogan et al.,2017).
We found that number of neighbors did not influence the vigilance or foraging of geese.These results contrast with those from previous studies on the same study system which showed significant effects of number of neighbors(Zhao et al.,2019,2020).The inconsistent results might result from the fact that geese were under different predation risks between this and the previous studies.Using a simulation model,Hirsch and Morrell(2011)found that at short attack distances,number of neighbors would pose a more significant influence on a focal animal’behaviors than relatively long attack distances.Our study site was an ephemeral island surrounded by water at the lower part of the Shengjin Lake.Therefore,this site was under more threat from the aerial avian predators(mostly pursuit predators)attacking from relatively long distances,but safe from mammal predators(mostly adopt ambush tactics)attacking from short distances.In contrast,previous studies were conducted at sites of the upper part of the Shengjin Lake where both mammal and avian predators both could pose great threats on the foraging geese.Another possibility is that geese might concentrate more effort to increase food intake rate to cope with the severe shortage of sedges resulting from the severe drought in 2019.Geese devoted less effort to competition with neighbors(25 vs 281 agonistic interactions,unpublished data)and had a slightly lower mean intake rate(55 vs 59)than geese in previous studies(Zhao et al.,2019,2020).
5.Conclusions
Our study supports the hypothesis that animals have significant different cost and benefits when they choose different spatial positions within a group.Moreover,dichotomously categorizing animals into peripheral and central individuals is logically reasonable but should be based on the cost-benefit tradeoff from an individual's perspective.We suggest researchers use a continuous measurement of spatial position when animals aggregate in large flocks and explore the exact boundary for the dichotomous classification.The boundary for classification may be different between different animal species or different populations of the same species with different environmental conditions,such as food abundances and quality,intensity of intraspecific competition and human disturbance and predator communities.
Authors'contribution
JMZ and WWC conceived the idea and designed the study.JMZ collected the data.MZW digitalized and analyzed the data.JMZ,MZW and CLL wrote the manuscript.All authors read and approved the final manuscript.
Ethics statement
All the observations were conducted under the natural status of geese flocks and have no influences on the behaviors of birds.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
This work was supported by the National Natural Science Foundation of China(grant numbers 31800325,31970500,31770571),and the Anhui Provincial Natural Science Foundation(grant numbers 1808085QC59,1908085QC128).
杂志排行

Avian Research的其它文章
- Use of 3D-printed animal models as a standard method to test avian behavioral responses toward nest intruders in the studies of avian brood parasitism
- Context dependent song-flight performance and timing in the Black-headed Bunting(Emberiza melanocephala)
- Habitat selection and food choice of White-naped Cranes(Grus vipio)at stopover sites based on satellite tracking and stable isotope analysis
- Year-round multi-scale habitat selection by Crested Tit(Lophophanes cristatus)in lowland mixed forests(northern Italy)
- Urban avifauna distribution explained by road noise in an Eastern European city
- Demographic and behavioral responses of the White-winged Dove to human disturbances during winter in northwestern Mexico