香蕉单宁的提取、结构及功能研究进展
2022-12-30曾祥权李倩倩姜微波李学杰惠博文
曾祥权,李倩倩,姜微波,杨 倩,奚 宇,李学杰,李 岩,惠博文,李 健,*
(1.北京工商大学食品与健康学院,北京 100048;2.北京市农林科学院质量标准与检测技术研究所,北京 100097;3.中国农业大学食品科学与营养工程学院,北京 100083;4.云南农业大学食品科学技术学院,云南 昆明 650201)
单宁属于植物次生代谢产物,是酚类家族的重要成员,其生物学作用主要与保护植物免受病菌、真菌、昆虫和食草动物的侵害有关[1]。通常将相对分子质量在500~3 000的植物多酚定义为单宁[2],其在自然界中分布广泛,通常存在于高等植物的根、茎、叶、花、果实、种子和树皮中,因具有多个酚羟基,易被氧化,通常呈米黄色、棕色或褐色无定形粉末状[3]。许多谷物(如大米、高粱和大麦等)、水果(如葡萄、柿子和莓类等)、饮料(如葡萄酒、茶和可可等)和可食海藻(如深海褐藻等)中都含有丰富的单宁[4]。研究表明,单宁具有抗氧化、抗肿瘤、抗炎症、抗菌、降血糖、降血脂和减缓金属对机体损伤等活性[5-7],且已经被成功应用于制革、食品、医药、饲料、包装等行业[8]。
根据化学结构的不同,单宁通常被分为水解单宁和缩合单宁两大类[9]。如图1所示,水解单宁是由葡萄糖和没食子酸/鞣花酸结合形成的多元酯类化合物,包括没食子酸吡喃葡萄糖、没食子单宁、鞣花单宁及其衍生物[10]。其中,水解单宁的重要特征之一是在酸、碱及酶的作用下发生水解,破坏其原有的化学结构。缩合单宁也被称为原花色素,是由黄烷-3-醇结构单元通过A型或B型连接形成的寡聚体或多聚体(图2)[11]。根据黄烷-3-醇结构单元的不同,缩合单宁能够被分为原花青素、原翠雀素、原菲瑟素和原刺槐亭醇[12]。缩合单宁的化学结构十分复杂,与其黄烷-3-醇结构单元的种类、连接方式、数量及C环C2、C3和C4的立体构型密切相关[13]。常见的黄烷-3-醇结构单元包括儿茶素、表儿茶素、表没食子儿茶素和表没食子儿茶素没食子酸酯等。缩合单宁结构单元的数量(即聚合度)对其功能活性同样具有重要影响。在特定条件下,具有更高聚合度的缩合单宁可能比低聚合度的缩合单宁具有强的生理功能。
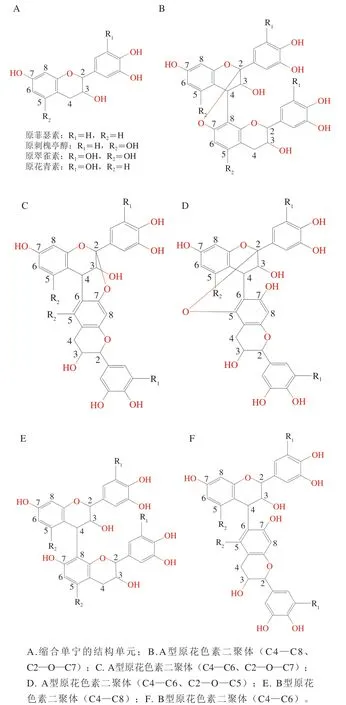
图2 缩合单宁的结构单元及不同类型的原花色素二聚体[11-12]Fig. 2 Structural units of condensed tannins and different types of proanthocyanidin dimers[11-12]
香蕉是芭蕉科(Musaceae)芭蕉属(Musa)的大型单子叶多年生草本植物,其果实是一种典型的呼吸跃变型水果,风味独特,深受消费者喜爱[14]。根据植株的外形特点和经济特征,香蕉主要分为香牙蕉、大蕉、粉蕉、龙牙蕉和贡蕉五大类,而目前市场上消费者普遍意义上说的香蕉多指香芽蕉[14]。作为世界四大水果之一,香蕉年产量超过1亿 t,产量排名居世界第二,仅次于柑橘[15]。香蕉喜高温多湿的环境,主要分布在南北纬度30°以内的热带、亚热带地区,全世界共有120个香蕉生产国和地区,产量排名前5的国家分别是印度、中国、印度尼西亚、巴西和厄瓜多尔[16]。香蕉富含多种营养功能成分,如糖类、维生素、多酚、矿质元素、膳食纤维、蛋白质和脂肪等[17],经常食用香蕉不仅有利于维持体内电解质平衡及酸碱代谢平衡,促进肠道蠕动,还可预防忧郁症、神经退行性疾病和心血管疾病。此外,香蕉中含有一定量的单宁[18],香蕉单宁具有抗氧化、抗菌、降血糖、降血脂、降胆固醇、清除有机磷农药、结合二价金属离子等多方面的生理活性[19-25],但是鲜有综述介绍香蕉单宁近年来的研究进展。基于此,本综述旨在对香蕉单宁的提取工艺、结构及功能活性研究进展进行系统性概述,为后续扩大香蕉单宁在食品行业中的应用提供一定的理论指导。
1 香蕉单宁的提取工艺
目前常见的单宁提取方法有溶剂提取法,超声波辅助提取法、微波辅助提取法和酶辅助提取法等,其中溶剂提取法和超声波辅助提取法的应用范围最广,溶剂提取法对原料、设备、时间和能耗等成本的需求低于其他方法,经济性和便捷性最高。但在提取率方面,与超声波辅助提取法(0.5%~1.3%)、微波辅助提取法(0.8%)和溶剂提取法(0.02%~0.50%)相比,酶辅助提取法(1.7%)提取香蕉单宁的效果最好。
1.1 溶剂提取法
溶剂提取法的原理是相似相溶,基于活性成分与杂质在不同溶剂中的溶解度差异从而得到目标产物,具有便捷和经济的优点。但与此同时,溶剂提取法也存在提取率低、耗时长和产品安全性低等缺点。对于香蕉单宁来说,常用水、丙酮、乙醇、甲醇和乙酸乙酯等溶剂进行提取。Kyamuhangire等[26]对4个品种香蕉果肉中的单宁进行水提,发现‘Kayinja’香蕉果肉中的水溶性单宁含量要远高于‘Katalibwambuzi’‘Musakara’和‘Kibuzi’香蕉,且未成熟和成熟‘Kayinja’香蕉果肉中单宁含量分别为0.841、4.799 g/kg。杨贤松等[27]利用丙酮溶液作为提取剂对鲜香蕉皮中单宁进行优化提取,当丙酮体积分数为50%、料液比为1∶12、提取温度为50 ℃、提取时间为2 h时,鲜香蕉皮中的单宁提取效果最佳。而Wu Tingru等[28]则利用响应面法优化得到香蕉皮中单宁的最佳提取工艺为提取温度62.85 ℃、乙醇体积分数75.44%、料液比1∶35.34、提取时间7.94 h,在此条件下单宁提取率为57.42%。与前两者不同的是,鲍金勇等[29]对比了不同体积分数的乙醇、丙酮和甲醇水溶液分别对香蕉皮单宁的提取效果,确定在料液比1∶14、提取温度40 ℃、提取时间10 h的条件下,采用体积分数60%的丙酮溶液作为提取剂,单宁的得率最高,约为0.901%。Laeliocattleya等[30]研究体积分数均为96%的乙醇、乙酸乙酯溶液对香蕉果肉中单宁的提取效果,结果表明采用乙醇溶液提取得到的香蕉单宁含量更高,约为209.83 mg/kg。
1.2 超声波辅助提取法
单宁属于细胞内产物,而细胞壁是从植物细胞中提取单宁的主要障碍。超声波辅助提取法能够通过超声波的机械、空化和热效应有效破坏植物组织的细胞壁,显著增强有机溶剂的穿透力,加速胞内物质的释放、扩散和溶解,具有省时、能耗低、效率高、不破坏活性成分等优点,因此在单宁提取方面具有广阔的应用前景[31]。赖晓琳等[32]对香蕉叶中缩合单宁的超声提取工艺进行研究,发现当利用体积分数85%乙醇溶液为提取剂、料液比为1∶28、提取时间25 min时,提取效果最佳,缩合单宁提取率为7.15 mg/g。刘涛等[33]借助超声波细胞破碎法提取香蕉皮粉末中的水解单宁,响应面优化最佳工艺条件为超声功率120 W、丙酮体积分数50%、料液比1∶30、提取时间40 min,此条件下水解单宁的提取率为78.11%。Vu等[34]优化香蕉皮中缩合单宁的提取工艺参数,结果表明当超声功率150 W、丙酮体积分数60%、料液比1∶12.5、超声温度30 ℃、超声时间5 min时,缩合单宁的得率最高,可达到13.11 mg/g。
1.3 微波辅助提取法
微波辅助提取法的原理主要是利用穿透力极强的微波渗透到植物细胞基质内部,使极性物质定向排列,导致细胞内温度急剧上升,同时内压增大,细胞壁和细胞膜形成裂纹或孔洞从而促进单宁的溶出[35]。微波辅助提取法不仅具有提取效率高、提取物质纯、能够有效保护单宁的优点,还可以有效降低溶剂消耗及废物产生量[31]。刘宁等[36]对香蕉皮粉末中的水解单宁进行微波提取,结果表明丙酮体积分数70%、料液比1∶40、提取温度50 ℃、提取4 min时水解单宁的提取率最高,约为72.37%。而王虹玲等[37]对香蕉皮单宁的微波提取工艺进行正交优化,结果表明体积分数75%乙醇溶液、微波功率375 W、料液比1∶30、提取时间80 s时,单宁的提取率高达87.04%。
1.4 酶辅助提取法
酶辅助提取技术是基于酶反应的高度专一性,通过利用纤维素、半纤维素或果胶酶对植物细胞壁进行水解,从而使单宁溶出,具有条件温和、选择性强的特点[31]。贾宝珠等[38]采用酶解辅助提取物法优化香蕉皮中的缩合单宁提取工艺,并评价单宁在不同反应体系下的稳定性。当复合植物水解酶添加量0.8%、酶解温度50 ℃、酶解45 min,缩合单宁的提取率可达到1.655%。随着贮藏温度和贮藏时间的增加,香蕉皮缩合单宁的稳定性逐渐降低。此外,VC、柠檬酸和苹果酸的添加能够提高香蕉皮缩合单宁的稳定性,而苯甲酸钠、山梨酸钾和钠离子、钙离子、钾离子、铜离子、铁离子等金属离子对其则具有明显破坏作用。
2 香蕉单宁的化学结构
如表1所示,目前对香蕉单宁结构的研究主要围绕香蕉果皮、果肉和茎秆中的水解单宁或缩合单宁展开,借助红外光谱法、凝胶渗透色谱、核磁共振技术、高效液相色谱法、液相色谱-质谱连用技术和质谱技术等对其结构单元、分子质量分布范围、聚合度范围、主要化学组成和结构单元连接方式等进行表征。徐树英等[39]测定香蕉茎秆中水解单宁的相对分子质量,凝胶渗透色谱的结果表明香蕉茎秆水解单宁的Z均相对分子质量、重均相对分子质量和数均相对分子质量分别为44 300.9、38 829.6、34 354.5,属于高分子多羟基苯酚酯类化合物。与之相比,Rebello等[40]鉴定香蕉皮缩合单宁的化学结构,结果表明香蕉皮缩合单宁主要由B型原花青素和原翠雀素组成,平均聚合度为19.2,结构单元包括儿茶素、表儿茶素、没食子儿茶素和表没食子儿茶素。其中,表没食子儿茶素是扩展单元的主要成分,物质的量分数约为70%,而表儿茶素是末端单元的主要成分,物质的量分数约为65%。对于香蕉果肉缩合单宁来说,Santos等[41]利用液质联用仪对27个品种绿熟期香蕉果肉中缩合单宁间苯三酚水解产物进行测定,发现表没食子儿茶素和表儿茶素是香蕉果肉缩合单宁的主要结构单元,其中表没食子儿茶素物质的量分数占88%~98%,并且鲜食香蕉缩合单宁中的表没食子儿茶素与表儿茶素的物质的量比(25.39~64.36)要普遍高于烹食香蕉(6.41~45.73)。而本课题组前期研究[20]发现从绿熟‘巴西蕉’果肉中提取得到的缩合单宁为相对分子质量866~1 522、聚合度3~5的B型原花青素和原翠雀素寡聚体,其中三聚体为主要成分,原翠雀素含量比原花青素高。此外,表儿茶素和表没食子儿茶素是香蕉果肉缩合单宁的主要结构单元,且表没食子儿茶素和表儿茶素的物质的量比约为2.64。
常见水果单宁的化学结构如表2所示,包括从柿子、葡萄、酸石榴、蓝莓、黑桑葚、蔓越莓、苹果和龙眼中提取得到的单宁[42-49]。其中,柿子果肉缩合单宁的结构单元组成和连接方式最复杂,且具有最高的聚合度,酸石榴果皮水解单宁的结构单元数量较低,组成相对简单。通过对比可以发现香蕉水解单宁的相对分子质量远高于酸石榴果皮和黑桑葚籽水解单宁,但化学结构尚不清楚,有待进一步解析。而香蕉缩合单宁的化学结构比较明晰,包含2~4种基本结构单元,主要化学组成是原翠雀素,其相对分子质量与聚合度与蔓越莓果肉缩合单宁较为接近,结构单元连接方式与葡萄籽和苹果果肉缩合单宁相同,均为B型连接。
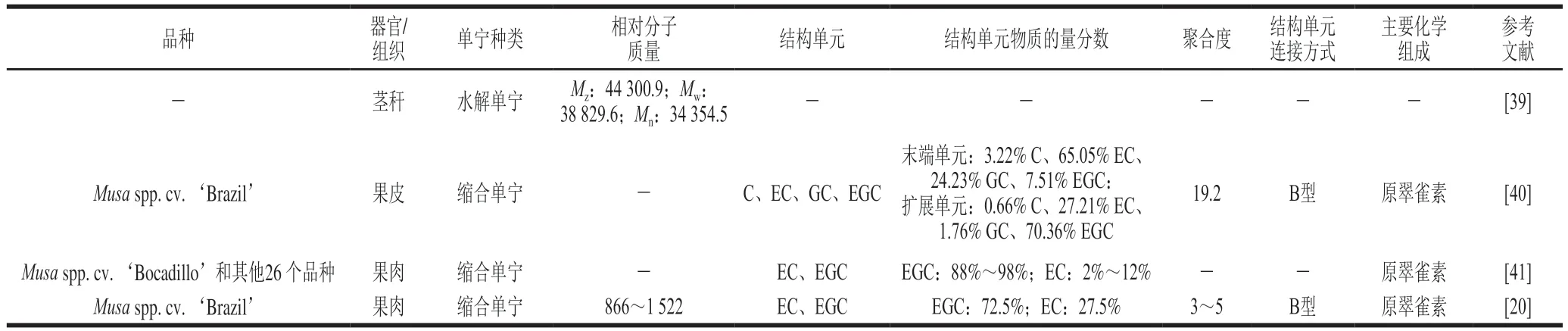
表1 香蕉单宁的化学结构Table 1 Chemical structures of banana tannins
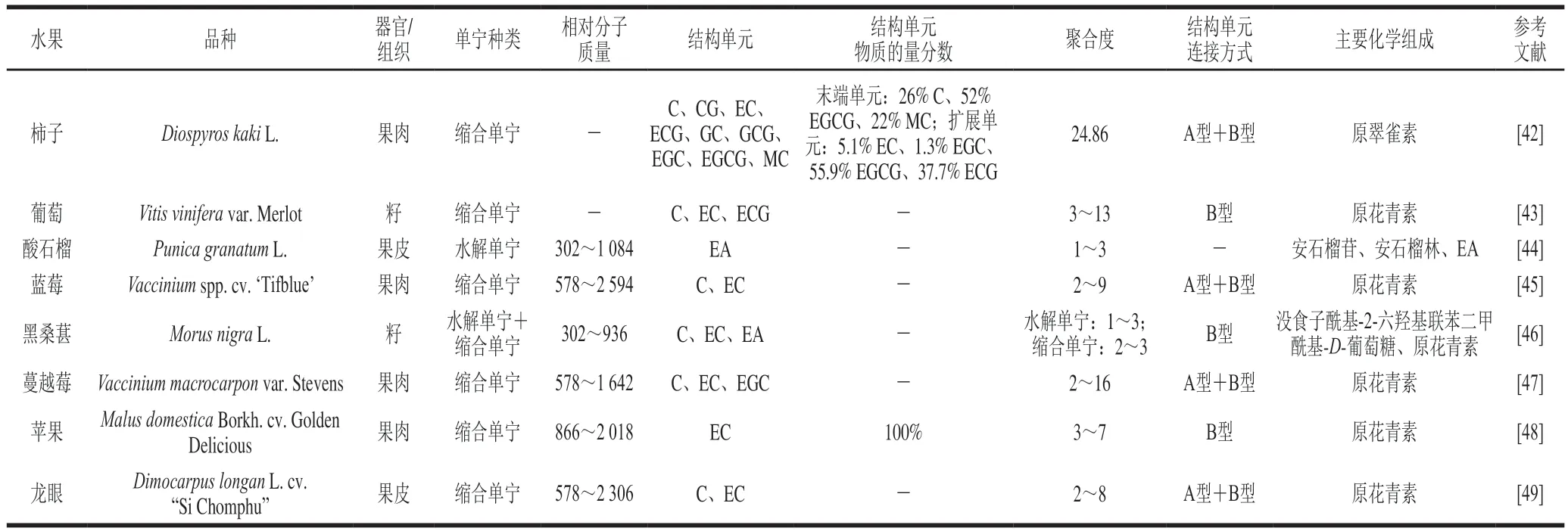
表2 常见水果单宁的化学结构Table 2 Chemical structures of tannins from common fruits
3 香蕉单宁的功能活性
单宁的功能活性与其多元酚结构所具有的独特理化性质(如蛋白质结合能力、金属离子络合能力、自由基清除能力和还原性等)密不可分。研究人员目前已经报道了香蕉单宁的抗氧化、抗菌、降血脂、降血糖、降低农药对机体毒性、吸收紫外线和结合金属离子等作用。
3.1 抗氧化活性
自由基能够通过攻击生物大分子破坏细胞的结构和功能诱导机体的氧化应激。单宁具有大量酚羟基,使其具有强还原性,从而能与自由基反应实现清除体内过剩自由基的目的,延缓机体的衰老。李晓静等[50]采用超声波法提取香蕉皮单宁,对其抗氧化活性进行评价,结果表明香蕉皮单宁对1,1-二苯基-2-三硝基苯肼(1,1-diphenyl-2-picrylhydrazyl,DPPH)、超氧阴离子自由基和羟自由基具有明显的清除能力,且半数抑制质量浓度(50% inhibitory concentration,IC50)分别为0.300、1.185、0.730 mg/mL,其中DPPH自由基和羟自由基IC50均低于VC对照组。此外,香蕉皮单宁对大豆油的自氧化具有很好的抑制作用,当体系中香蕉皮单宁添加量为0.04%(质量分数)时大豆油的过氧化值最低。因此,香蕉皮单宁具有被开发为新型油脂氧化抑制剂的潜力。Bennett等[18]分析从‘Terra’和‘Mysore’香蕉果肉中提取得到的缩合单宁的抗氧化活性,发现两种香蕉果肉缩合单宁提取物在体外均能有效清除DPPH自由基,其中‘Mysore’的清除能力显著高于‘Terra’,可能是由于‘Mysore’中缩合单宁含量(204.7~259.4 μg/g(原花青素寡聚体当量))远高于前者(37.5~46.3 μg/g(原花青素寡聚体当量))所致。Rosales-Reynoso等[51]对比了墨西哥4个品种绿熟香蕉果肉中的缩合单宁和水解单宁含量差异及其2,2’-联氮-双(3-乙基苯并噻唑啉-6-磺酸)(2,2’-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid),ABTS)阳离子自由基清除能力,结果表明,‘Morado’香蕉果肉中的缩合单宁(44.65 mg/g(没食子酸当量))含量最高,缩合单宁的ABTS阳离子自由基清除能力为55.87 μmol/g(Trolox当量)。与之相比,‘Enano’香蕉果肉中的水解单宁(19.65 mg/g(没食子酸当量))含量最高,水解单宁的ABTS阳离子自由基清除能力为13.14 μmol/g(Trolox当量)。
从香蕉不同组织和器官中提取得到的单宁抗氧化活性存在一定的差异。在最近的一项研究中,Bashmil等[52]分别测定6个澳大利亚品种成熟和未成熟香蕉果皮和果肉单宁含量及总抗氧化能力。对于果皮,成熟‘Ducasse’香蕉皮中单宁含量(3.34 mg/g(儿茶素当量))最高,但总抗氧化能力最强的是从成熟‘Plantain’香蕉果肉中提取得到的单宁,约为0.25 mg/g(VC当量)。但就果肉而言,未成熟‘Ladyfinger’香蕉果肉中的单宁含量及单宁总抗氧化能力均显著高于其他果肉,分别为1.52 mg/g(儿茶素当量)和1.03 mg/g(VC当量)。而Sasipriya等[53]比较了Musaspp.Ensete superbum和Musa paradisiacavar.Monthan香蕉花和果肉丙酮提取物中单宁的含量和总抗氧化能力。其中Ensete superbum香蕉花提取物中的单宁含量为276.2 mg/g(单宁酸当量),DPPH自由基、ABTS阳离子自由基清除能力和铁还原能力分别是441.0 mol/g(Trolox当量)、125.7 mol/g(Trolox当量)和64.4 mol/g(Fe2+当量),均比其他提取物高。
3.2 抗菌活性
研究表明,单宁能够通过影响细菌细胞壁结构的完整性干扰细菌的黏附力从而发挥抗菌效应[54]。与传统的抗菌药物(如抗生素和磺胺类药物)相比,单宁具有安全和无毒副作用的优点,因而具有被开发为新型抑菌剂的潜力。目前,关于香蕉单宁抗菌活性的研究大多围绕从香蕉皮中提取得到的单宁展开。赵立[55]测定了香蕉皮单宁对不同细菌、霉菌和酵母菌的体外抑制活性,结果表明香蕉皮单宁对大肠杆菌和金黄色葡萄球菌的最小抑制剂量(minimal inhibitory concentration,MIC)为0.1%(质量分数,此段同),对枯草芽孢杆菌的MIC为0.2%,对黑曲霉和黄曲霉的MIC均为1%,对啤酒酵母几乎没有抑制作用。此外,香蕉皮单宁对细菌的抑制作用强于山梨酸钾,表明其可能是一种具有开发前景的新型细菌抑制剂。而李丽等[56]研究表明香蕉皮单宁对细菌、霉菌和酵母菌均具有明显抑制效果,且对酵母菌的抑制作用最强,抑菌效果为啤酒酵母>米曲霉>根霉>大肠杆菌>枯草芽孢杆菌,这可能与提取得到的香蕉皮单宁化学组成和结构差异有关。此外,李丽等[56]还考察了温度、pH值和盐质量分数等因素对香蕉皮单宁抑菌性能的影响,发现单宁的抑菌能力不受高温处理影响,但随着pH值的增加(2.0~8.0)逐渐减弱,随着盐质量分数的增加(1%~7%)显著增强。一方面,香蕉皮单宁属于酸性物质,可能在碱性环境中被氧化从而失去抑菌活性;另一方面,盐的存在对于单宁的抗菌能力具有一定的协同作用。李海帆等[19]研究香蕉皮单宁提取物对大肠杆菌的抑菌机理,结果表明提取物对大肠杆菌的MIC为2.5 mg/mL,经MIC单宁处理后大肠杆菌菌液中的核酸质量浓度(0.054 mg/L)显著增加。因此,香蕉皮单宁对大肠杆菌细胞膜完整性的破坏对其抗菌活性具有重要贡献。
3.3 降血脂活性
心脑血管疾病是全球范围内高死亡率的一类疾病,而高血脂是心血管疾病的主要风险因子之一。近年来的研究结果表明许多单宁具有显著的降血脂活性,与其对脂质在胃肠道的吸收、血清和肝脏脂蛋白的分布、脂质代谢基因和相关酶表达、小肠胆固醇胶束溶解度和粪便脂肪排泄的多重作用有关[57-60]。Li Xiangxin等[23-25]对香蕉果肉缩合单宁的体内外降血脂活性开展了一系列研究。首先,在模拟小肠条件下分析香蕉果肉缩合单宁与模拟消化液及各组分之间的相互作用,并探究单宁对脂质消化的影响,结果表明,香蕉果肉缩合单宁能够使模拟消化液、胰脂肪酶、胆固醇酯酶、胆酸盐、胆固醇和氯化钙溶液的浊度和平均粒径增大,并使消化液及其成分溶液的Zeta电位和热力学特性发生改变,说明缩合单宁与上述组分之间发生絮凝和聚集。因此,认为香蕉果肉缩合单宁能够通过与消化液及各组分之间相互作用抑制脂质在小肠内的消化吸收。其次,探究香蕉果肉缩合单宁与胆固醇酯酶、胆固醇和4种胆酸盐的相互作用机制。缩合单宁能够通过混合竞争抑制模式降低胆固醇酯酶活性,其与酶结合形成复合物的过程是放热的且自发的,分子之间主要作用力为氢键、疏水相互作用和范德华力,且这种结合能够改变酶的二级结构组成。此外,香蕉果肉缩合单宁能够通过氢键、疏水相互作用和范德华力等非共价作用与胆固醇或胆酸盐发生絮凝和沉淀,这种结合是由熵、焓共同驱动的。最后,该研究团队以C57BL-6小鼠为研究对象,建立高胆固醇小鼠模型,探究香蕉果肉缩合单宁对小鼠体内胆固醇代谢的调节作用[61]。缩合单宁处理(200 mg/kg(原花青素B2当量),48 d)能够显著降低高胆固醇摄入组小鼠血液中谷丙转氨酶、谷草转氨酶、碱性磷酸酶和谷胱甘肽-S转移酶活力及肝脏中丙二醛含量,提高肝脏总抗氧化能力、总超氧化物歧化酶、谷胱甘肽、谷胱甘肽过氧化物酶和过氧化氢酶水平,减少肝脏和血液中总胆固醇、总甘油三酯、总胆汁酸、低密度脂蛋白胆固醇水平,同时增加高密度脂蛋白胆固醇含量,还能促进小鼠粪便中胆固醇的排泄。代谢组学分析结果表明,高胆固醇摄入组和胆固醇香蕉果肉缩合单宁复合处理组小鼠肝脏的差异表达基因数目分别为1 967、2 178个。相比于高胆固醇摄入组,胆固醇香蕉果肉缩合单宁复合处理组小鼠肝脏差异基因表达以下调为主。香蕉果肉缩合单宁能够通过调节CETP、LCAT、LDL-R、ABCG5/8、ABCA1、Hmgcr、CYP7A1、CYP8B1、Strada、Camkk2、Adra1a、Adipor2、Ppp2r5b和Ppp2r5e基因的表达调控机体内胆固醇代谢过程(胆固醇逆转运、胆固醇合成、胆汁酸代谢和腺苷酸活化蛋白激酶途径),进而减少胆固醇在小鼠肝脏中的蓄积。以上研究结果表明,香蕉果肉缩合单宁是一种能够有效减少脂质吸收和降低过量胆固醇摄入诱导代谢性脂肪肝病风险的功能性成分。
3.4 降血糖活性
单宁的降血糖活性与其对参与葡萄糖代谢相关基因表达和酶活性的影响、对胃肠道葡萄糖的吸收干扰及对蛋白质非酶糖基化的抑制作用有关[62-63]。de Angelis-Pereira等[64]评价富含单宁的绿熟香蕉皮粉和果粉对正常Wistar大鼠血糖水平的影响,其中香蕉皮粉和果粉中单宁的含量分别为1 259、1 670 mg/100 g。尽管香蕉皮粉和果粉干预15 d后对大鼠餐后血糖变化无明显影响,但在饲料中分别添加质量分数10%和15%香蕉果粉后大鼠的血糖指数仅为对照组的87.7%和88.2%。由此可知,摄入一定剂量富含单宁的绿熟香蕉果粉可能有助于预防糖尿病。虽然单宁在体内外的降血糖活性显著,但是依然存在黏附性差、溶解度低、苦涩味重、易氧化降解等缺点,很难在常见食品药品中直接添加应用,这是制约其在食品工业和医疗保健行业中应用的关键技术难题[65]。因此,将单宁改性进而扩大其应用范围的相关研究在近年来受到了一定关注。在最新的一项研究中,本研究团队借助自由基介导法将香蕉果肉缩合单宁接枝到菊糖上成功制备得到香蕉果肉缩合单宁-菊糖共价复合物,共价复合物的接枝比率和接枝效率分别为(357.54±2.98)g/kg和(74.57±1.44)%。抗氧化活性评价的相关数据显示香蕉果肉缩合单宁-菊糖共价复合物的ABTS阳离子自由基、DPPH自由基清除能力、铜离子和铁离子还原能力均显著高于菊糖。表征结果证明了香蕉果肉缩合单宁与菊糖成功接枝,且两者的结合位点可能在与菊糖C6位连接的羟基和香蕉果肉缩合单宁黄烷-3-醇结构单元C6/C8位之间。香蕉果肉缩合单宁在与菊糖接枝后贮藏稳定性显著增加,化学改性也明显提高了菊糖的体外降血糖活性。香蕉果肉缩合单宁-菊糖共价复合物的葡萄糖吸附能力和葡萄糖透析延迟指数均比菊糖高。同时,共价复合物还表现出明显的α-淀粉酶和α-葡萄糖苷酶抑制活性,且显著高于菊糖。香蕉果肉缩合单宁-菊糖共价复合物对α-淀粉酶的抑制作用是可逆的,但却是α-葡萄糖苷酶的不可逆抑制剂,共价复合物对两种酶的抑制均为混合型抑制模式,能够通过静态机制对酶的荧光进行猝灭,与酶的结合常数和结合位点数均大于菊糖[21]。基于以上研究结果认为,改性后的香蕉果肉缩合单宁在被开发为新型降血糖功能性成分方面具有一定潜力。
3.5 降低农药对机体的毒性
尽管农药在农作物生产上对控制害虫、杂草和病害的效果不可忽视,但近年来过度使用农药对生态环境造成的污染也逐渐成为棘手的问题。长期暴露于高浓度的农药会对机体造成多方面损害,如显著增加呼吸、神经、免疫和生殖系统疾病的发病风险等[66]。农药对机体产生的一系列损伤被证明能被许多天然产物缓解,其中就包括植物单宁。单宁可能通过抗氧化、抗炎症、抗凋亡、抗突变、抗抑郁、刺激免疫系统、调节神经递质和激素水平、脂质和能量代谢、促进农药排泄及改善器官功能等相关机制降低农药对机体的毒性[67]。因此,农药暴露人群可以通过摄入富含单宁的膳食从而促进机体损伤的修复。本研究团队通过研究香蕉果肉缩合单宁与草甘膦的体外直接相互作用,发现香蕉果肉缩合单宁能够通过和草甘膦分子在水溶液中形成不溶性颗粒从而实现对其的清除(表3)。随着缩合单宁含量的增加(0.25~1.00 mg/g(原花青素B2当量)),香蕉果肉缩合单宁的草甘膦清除能力逐渐增强(10.2%~59.14%)。静电相互作用、氢键和疏水相互作用在香蕉果肉缩合单宁的体外草甘膦清除能力中扮演着重要角色。此外,香蕉果肉缩合单宁-草甘膦共沉物的光谱学性质和表面形貌特征也证实了香蕉果肉缩合单宁和草甘膦分子之间的直接相互作用[22]。
类似的结果在香蕉果肉缩合单宁与百草枯和丙溴磷的相互作用研究中也被观察到[68-69]。研究人员发现百草枯和丙溴磷均能在体外与香蕉果肉缩合单宁反应产生絮状沉淀,可能与两者之间的非共价相互作用有关,且随着香蕉果肉缩合单宁浓度的增加,单宁对两种农药的清除率显著上升。而在体内,香蕉果肉缩合单宁通过促进百草枯排泄、调节氧化应激水平和改善肺纤维化水平从而减缓百草枯暴露引起的小鼠损伤。其中,香蕉缩合单宁百草枯复合处理组在24 h代谢实验第8小时收集的粪便中农药含量是百草枯对照组的1.16 倍,而肺损伤实验数据显示复合处理组小鼠肺组织中的羟脯氨酸和丙二醛含量分别比百草枯对照组低18.1%和21.7%,过氧化氢酶活性高5%,且肺纤维化程度明显减弱。丙溴磷在小鼠中的毒性也能被香蕉果肉缩合单宁明显缓解。一次性同时灌胃高剂量单宁(10.4 mg/kg(表儿茶素当量))和丙溴磷(89.5 mg/kg)组小鼠粪便中的农药含量比丙溴磷对照组高61.23%,预先1 h灌胃高剂量单宁同样使小鼠粪便中丙溴磷排出量较对照组增加48%。综上,香蕉果肉缩合单宁可能通过在小鼠胃肠道内沉淀丙溴磷和百草枯从而降低两种农药的生物毒性。
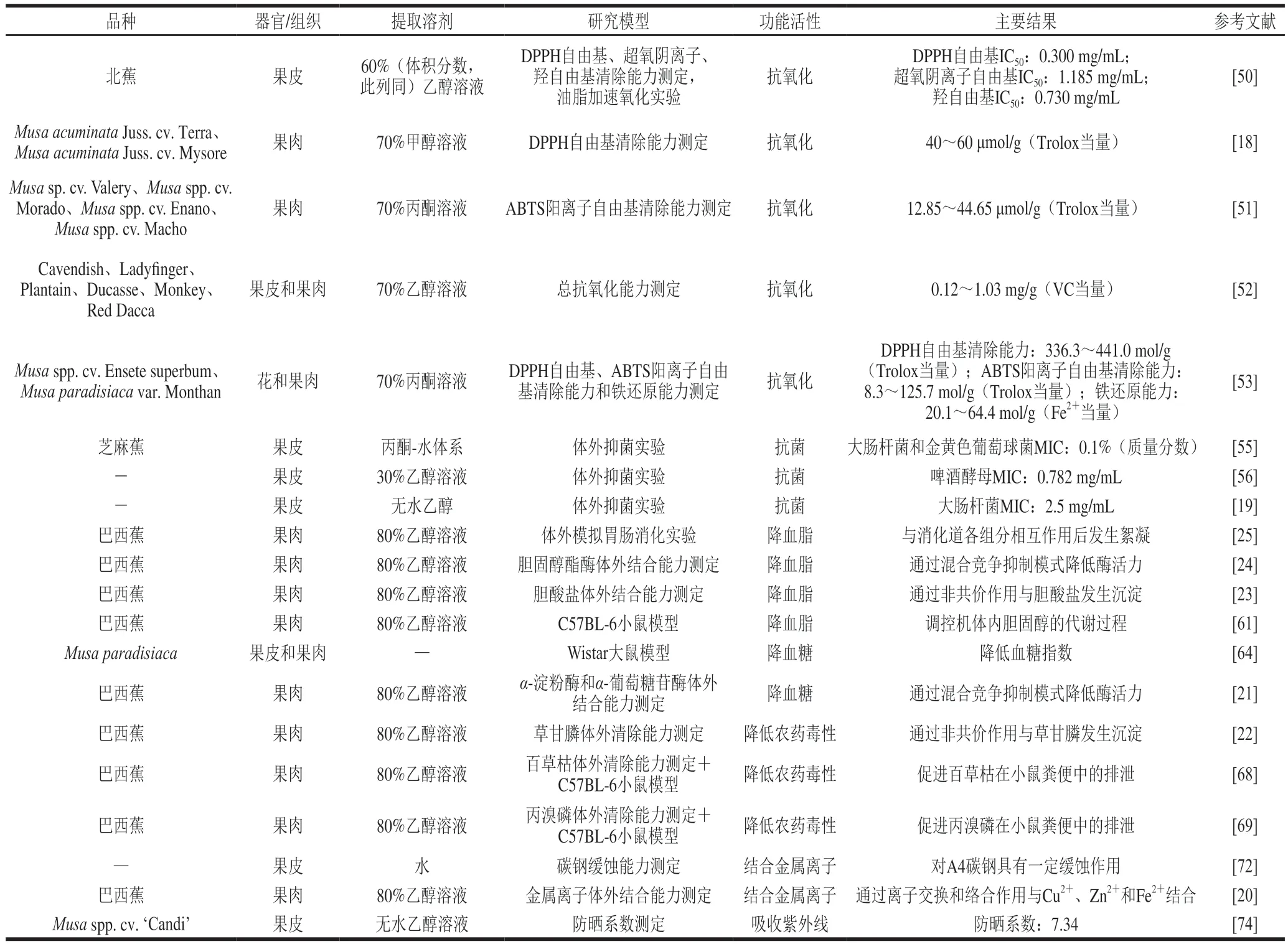
表3 香蕉单宁的功能活性Table 3 Functional properties of banana tannins
3.6 结合金属离子
作为优良的金属螯合剂,单宁被证明能够通过配位作用与金属离子发生结合[5,70]。而这种配位作用能够使单宁吸附在金属表面形成保护层从而表现出明显的缓蚀效果[71]。任雪峰等[72]首次评估了香蕉皮单宁对A4碳钢的缓蚀作用,结果表明当酸洗时间8 h、酸洗温度40 ℃、HCl浓度0.8 mol/L时,添加10 mL香蕉皮单宁提取液对碳钢的缓蚀率可达81.35%,缓蚀速率为18.13 g/(m2·h)。将香蕉皮单宁与乌洛托品和乙酸锌复配能够大大改善其对碳钢的缓蚀效果,且缓蚀率能超过90%。本研究团队采用光谱和质谱技术进一步研究香蕉果肉缩合单宁与3种二价金属离子(铜离子、锌离子和亚铁离子)之间的相互作用机制[20]。结果表明,二价金属离子对香蕉果肉缩合单宁荧光的猝灭模式为静态猝灭。香蕉果肉缩合单宁可以通过离子交换和络合作用与金属离子结合,但邻苯二酚和焦棓酚离子交换是主要的结合作用。香蕉果肉缩合单宁与金属离子相互作用的结合常数、结合位点数、吉布斯自由能绝对值、对金属离子结合能力大小均为香蕉果肉缩合单宁亚铁离子复合处理组>香蕉果肉缩合单宁铜离子复合处理组>香蕉果肉缩合单宁锌离子复合处理组,说明香蕉果肉缩合单宁对亚铁离子的亲和性高于铜离子和锌离子。对3种香蕉果肉缩合单宁-金属离子共沉物进行表征,相关数据进一步证明了香蕉果肉缩合单宁对二价金属离子的结合作用。研究结果有助于理解缩合单宁在与金属离子有关的生命系统中所发挥生物学作用,包括缩合单宁的自由基清除活性,对机体中金属暴露介导毒性的保护作用,对具有金属活性中心相关酶的抑制活性,以及对胃肠道中矿质元素吸收的抑制作用等。此外,还可能有助于缩合单宁-金属纳米粒子、基于缩合单宁用于识别金属离子的传感器及用于金属离子吸附膜材料的开发和利用。
3.7 吸收紫外线
过度紫外线暴露会导致皮肤色素沉积,提高机体患皮肤癌的风险。研究表明,植物单宁具有一定的紫外线保护作用,这与其结构中存在能够吸收紫外波长的苯环有关[73]。Laeliocattleya等[74]初步探究了‘Candi’香蕉皮单宁提取物对紫外线的吸收作用,发现提取物中单宁含量约为6.90 mg/g,防晒系数可达7.34,初步推测香蕉皮单宁提取物在防晒化妆品中具有应用潜力。
4 结 语
本文总结了香蕉单宁的提取工艺、化学结构及功能活性的研究进展,对推动香蕉功能性成分开发,加强香蕉资源的合理利用具有一定的指导意义。尽管如此,目前的研究依然存在一定的局限性。今后,研究人员可以围绕以下6个方面继续开展相关工作。首先,开发提取香蕉单宁的新工艺,包括超临界萃取法、离子沉淀提取法和吸附分离提取法等。其次,完善香蕉单宁的分离纯化方法。目前关于香蕉单宁的分离纯化研究十分鲜见,而香蕉单宁粗提物中通常含有蛋白、多糖、色素和脂类等杂质,因此需要采用大孔树脂吸附法、膜分离法、高速逆流色谱法和凝胶色谱法等对其分离纯化以便后续的结构分析或功能性研究。第三,进一步鉴定香蕉单宁的化学结构,包括水解单宁的种类及各组分含量、缩合单宁各寡聚体的物质的量比及扩展单元和末端单元的具体排列方式等。第四,探究香蕉单宁的其他潜在功能活性,包括抗肿瘤、抗炎症、抗病毒活性及对神经系统的保护作用等。第五,对香蕉单宁进行适当改性,如与不同蛋白或多糖共价接枝,解决单宁黏附性差、溶解度低、苦涩味重进而易氧化降解等问题。最后,扩大香蕉单宁在食品工业中应用,可以尝试将其开发成食品营养强化剂、防腐剂、抗氧化剂、饮料和酒类的澄清剂以及新型功能包装材料等。
