A Comparative Study on the Buoyancy of Resting Eggs from Different Artemia Populations, with Emphasis on Its Relationships with Habitat Salinity and Biometric Characters of Resting Eggs
2022-12-27ZHOUJiaxuanHUANGZhikeGUOYanandSUNShichun
ZHOU Jiaxuan, HUANG Zhike, GUO Yan, and SUN Shichun,
1) The Key Laboratory of Mariculture (Ministry of Education), and Institute of Evolution & Marine Biodiversity,Ocean University of China, Qingdao 266003, China
2) Xinjiang Uygur Autonomous Region Fisheries Research Institute, Urumqi 830002, China
Abstract The buoyancy of Artemia resting eggs is a feature with both biological and economic importance. Since the buoyancy of resting eggs is dependent on the specific weight of the eggs or salinity of ambient water, we suppose that habitat salinity may exert a selection pressure on resting egg buoyancy, and thereby lower habitat salinity may result in better floating capacity of resting eggs and vice versa. In this study, we compared the floating capacity of resting eggs from 25 Artemia populations. The results showed that the floating capacity of resting eggs varied greatly among different populations. The minimum salinity to float some eggs varied from 0 to 180. The salinity at which all resting eggs floated varied from 80 to 320. The FS50 (salinity with 50% eggs floating)of Zhundong and Yuncheng population was not detectable (over 50% eggs floating in distilled water), that of Kyêbxang Co population was 4.3,while the maximum value found in Dabancheng population was 234.5. In terms of the ‘apparent specific weight’ of resting eggs, 18 populations exhibited a single-peak distribution pattern, while the other 7 populations showed a multiple-peak or non-peak distribution. Negative correlations were found between FS50 and chorion thickness, and between FS50 and the volume percentage of the chorion in eggs, supporting a previous standpoint that shell thickness was a determinative factor for the floating capacity of resting eggs. A positive correlation was determined between FS50 and habitat salinity. The hypothesis that habitat salinity may cause a directional selection on the buoyancy of resting eggs seems to be true.
Key words brine shrimp; cyst; floating capacity; shell thickness
1 Introduction
The brine shrimpArtemiaLeach, 1819 (Crustacea: Anostraca)is a small planktonic crustacean living in hypersalinity habitats all over the world except the Antarctic (Vanhaeckeet al., 1987; Van Stappen, 2002). The genus contains several bisexual species and numerous parthenogenetic populations (Asemet al., 2020; Sainz-Escuderoet al., 2021).Both bisexual and parthenogeneticArtemiahave two reproductive modes: oviparity producing resting eggs (cysts)and ovoviviparity producing free-swimming nauplii in a single individual’s lifespan, and females have the ability to switch between the two reproduction modes (Lavens and Sorgeloos, 1987; Nambuet al., 2004; Wanget al., 2019). AsArtemianauplii are favorite living foods for larval fishes and crustaceans, and resting eggs are easy to store and circulate, resting eggs have great economic value and are commercially harvested and traded around the world. In addition to the large number of wild populations,Artemiaare also cultured in manmade brine ponds in countries like Brazil, Vietnam, China, Philippines and Thailand (Van Stappenet al., 2020).
Artemiaresting eggs, which are gastrulas and each contains some 3000 – 4000 cells (Clegg and Conte, 1980), represent the diapause stage inArtemialifecycle and allowArtemiato overwinter and survive harsh conditions (Lavens and Sorgeloos, 1987). Among brine shrimps,Artemia monicaVerrill, 1869 in Mono Lake (Lenz, 1980; Drinkwater and Crowe, 1991)and the parthenogeneticArtemiain Dabancheng Salt Lake (Renet al., 1996)were reported to produce benthic resting eggs (see further). All otherArtemiaspecies/populations produce resting eggs floating on the water surface, though a certain fraction of benthic resting eggs may coexist with the floating ones (e.g., Abatzopouloset al., 2006; Arashkevichet al., 2009; Litvinenkoet al., 2016). This is different from resting eggs of many other planktonic animals, which usually sink to the sediments after being laid,e.g., resting eggs of rotifers (Gilbert, 2017)and copepods (Tachibanaet al., 2019). The floating capacity of resting eggs brings obvious benefits to the life and procreation ofArtemia. For instance, they can be kept away from the lower oxygen of bottom waters, exposed to light, and blown leeward and aggregated along the shoreline, where they have more opportunities to encounter the low salinity water (from precipitation/runoff)and higher temperature in the next warming season. Moreover,floating eggs have more opportunities of dispersal by wind and aquatic migratory bird transportation (Proctor, 1964;Van Stappen, 2002; Eimanifaret al., 2006). Because it is difficult to catch resting eggs from sediments, the commercial harvestability ofArtemiaresting eggs is largely dependent on their buoyancy. Although the buoyancy ofArtemiaresting eggs is a character of both biological and economical importance, few studies have been documented. Sorgeloos (1996)and Abatzopouloset al. (2006)compared the resting egg buoyancy ofArtemia urmianaGünther, 1899(Urmia Lake, Iran)andArtemia franciscanaKellogg, 1906(Great Salt Lake, USA)and found that the former species had a higher fraction of sinking eggs than the latter species.Renet al. (1996)tested the buoyancy of resting eggs from Dabancheng Salt Lake (Xinjiang, China)and found they were not floating until increasing the salinity to 140. Litvinenkoet al. (2016)found that the hatching rate of floating resting eggs was significantly higher than that of sinking eggs in most Western Siberia lakes, but might not be so obvious in some seasons or lakes. Sura and Belovsky (2016)reported that selective harvesting of floating eggs had caused a decrease in the buoyancy of resting eggs in the Great Salt Lake, and the mortality of nauplii was negatively related with the buoyancy of resting eggs. Their experimental study also showed that parents with adequate food sources tended to produce floating eggs, while undernourished parents tended to produce sinking eggs even if they were hatched from floating eggs.
The habitat ofArtemiavaries considerably in many aspects such as ion composition, climatic condition, altitude,etc. (Triantaphyllidiset al., 1998). Since the buoyancy of resting eggs is dependent on the specific weight or the salinity of ambient water, we suppose that the habitat salinity may exert a selection pressure on the buoyancy of resting eggs, and thereby the buoyancy of resting eggs from differentArtemiapopulations may be different and is correlated with their habitat salinity. To test this hypothesis,we compared the buoyancy of resting eggs from differentArtemiapopulations. Moreover, previous studies have suggested that the absence of floating capacity in resting eggs ofA. monicawas a consequence of their thin shell (Drinkwater and Crowe, 1991), and the lower floating capacity ofA. urmianaresting eggs (compared with those ofA. fran-ciscana)was because of their narrower and more compressed alveolae in the shell (Abatzopouloset al., 2006). To further examine the relationships between the buoyancy and some intrinsic features of resting eggs, the dry weight of intact and decapsulated eggs, the chorion thickness, the weight and volume proportions of the chorion in the intact eggs were also studied in this paper.
2 Materials and Methods
2.1 Artemia Resting Eggs
Resting eggs were from 25 hypersaline habitats, including 21 inland salt lakes and 4 coastal solar salterns. Their locations, abbreviated codes, habitat salinities, sampling time,reproductive modes, and hatching percentages are shown in Table 1. Hatching percentages of resting eggs were determined at salinity of 30 and temperature of 25℃. For some populations, resting eggs were treated with 3% H2O2for 40 min to break diapause before determining the hatching rate.In a previous study, Sura and Belovsky (2016)accounted that resting egg buoyancy was unlikely influenced by cyst viability or the length of time the resting eggs were in diapause. The hatching rate of their oldest sample (stored for 23 years)was 40%. Accordingly, only resting egg samples with a minimum hatching percentage of 40% were used in the present study.
2.2 Buoyancy Experiments
The buoyancy ofArtemiaresting eggs was tested with solutions of gradient salinities (0, 20, 40, 60, 80, 100, 120,140, 160, 180, 200, 220, 240, 260, 280, 300, 320). To determine theFS50(salinity with 50% resting eggs floating; see below)precisely, resting eggs of some populations were tested under additional salinities,e.g., salinities of 10, 30 and 50 were used when testing the buoyancy of KXC resting eggs.Solutions with salinities of 0 to 260 were prepared by dissolving NaCl in distilled water, and higher-salinity solutions were prepared by dissolving MgCI2·6H2O in distilled water.
Buoyancy experiments were conducted in 150 mm (height)× 18 mm (diameter)test tubes each containing 10-cm deep experimental medium. At the beginning of each test, a small number (100 – 200)of non-damaged resting eggs were put into each test tube, and incubated at 0℃ for 12 h and then at 20℃ for 1 h. Resting eggs floating on the water surface and sinking to the bottom of test tubes were counted,respectively. Sometimes there were few resting eggs suspending in the water column, 50% of such eggs were counted as floating eggs and the other 50% as sinking eggs. Three replicates were performed for each test. The following parameters/terms were used:

Fraction of the ‘apparent specific weight’ of resting eggs(FSW): The difference of PF values between two neighboring experimental salinities. For example, if PFs were 25% and 45% at the neighboring salinities of 40 and 60 respectively, the FSW40–60would be: 45% - 25% = 20%.Here the ‘apparent specific weight’ is an ideal concept, and it is expressed by the salinity at which eggs are expected to suspend in the water column (in the above example, the‘apparent specific weight’ of the eggs of the fraction should range from 40 to 60).
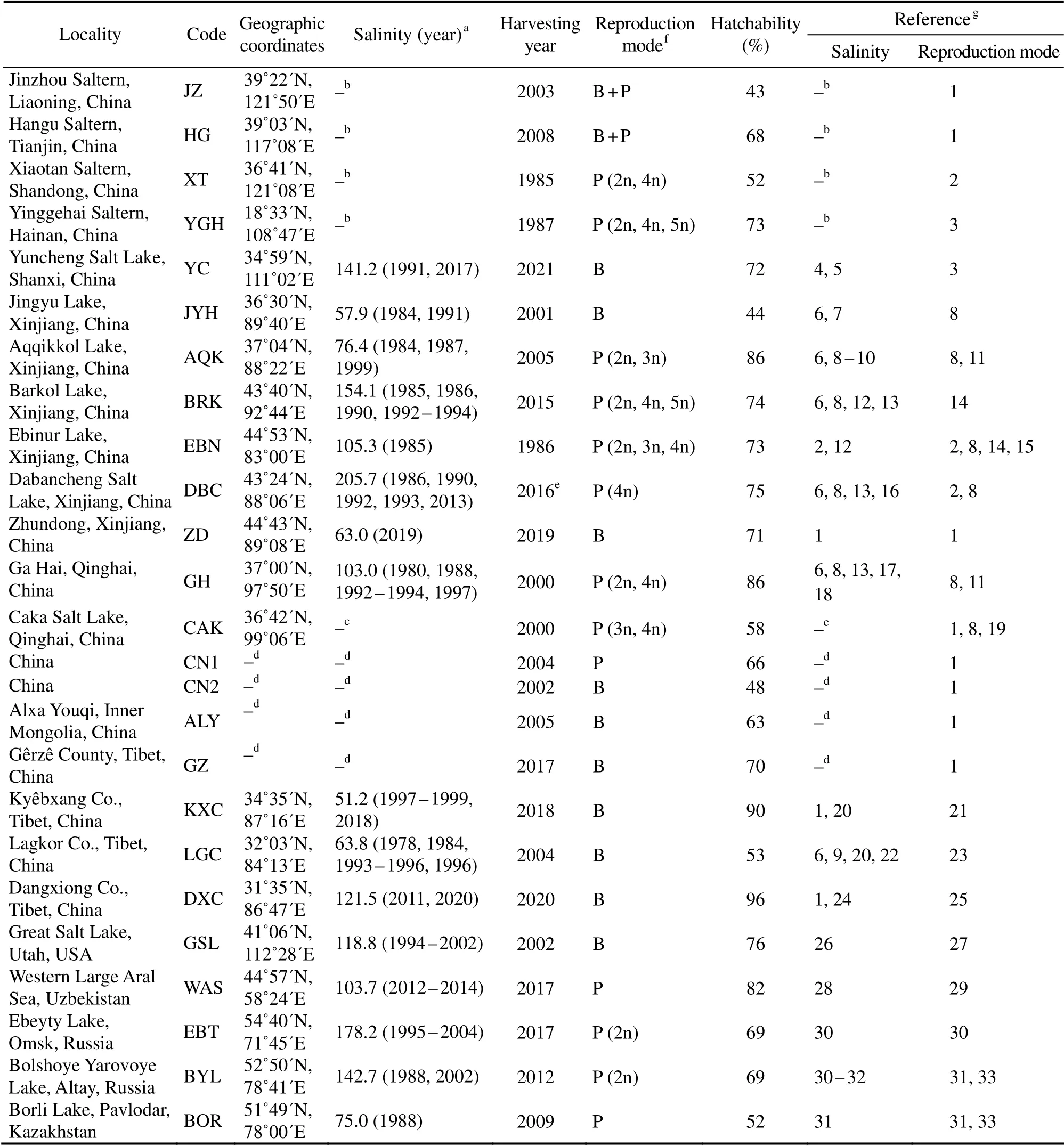
Table 1 Artemia resting eggs used in the present study
FS50: The salinity that 50% of resting eggs float. It was estimated on the linear regression equation obtained from PF values under several key salinities.
2.3 Determination of the Dry Weight of Intact and Decapsulated Eggs
For each population, a small number of resting eggs were incubated in distilled water at 0℃ for 12 h to make sure the resting eggs fully hydrated. Then eggs were divided into two groups. One group of eggs were decapsulated using 8% NaClO solution for 2 – 3 min, washed with distilled water until there were no impurities; the other group were untreated. Then 50 – 150 intact or decapsulated eggs were counted, and put into a 0.2-mL centrifuge tube (pre-heated at 65℃ for 24 h), dried at 65℃ for 24 h, and then weighed using a MYA 5.4Y (Radwag Wagi Electroniczne)electronic balance. The average weight of individual egg was calculated. Three replicates were performed for each population.The following parameter was calculated for each population:

2.4 Determination of the Diameter and Volume of Intact and Decapsulated Eggs
Resting eggs were incubated in seawater with a salinity of 30 at 0℃ for 12 h. Then eggs were divided into two groups for determining diameters of intact and decapsulated eggs(method of decapsulation, see above), respectively. Diameters were measured using an Olympus SZX16 stereomicroscope equipped with an Olympus DP74 camera. For each population, 100 intact and 100 decapsulated eggs were measured, respectively. The volume of each egg was calculated using the volume formula for a solid sphere:
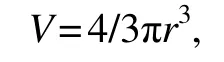
whereVandrare the volume and radius of the egg, respectively. The following parameters are calculated:

2.5 Statistic Analysis
To analyze factors that potentially influence the buoyancy of resting eggs, correlation and regression analyses were conducted between 1)theFS50and the dry weight of intact eggs; 2)theFS50and the weight percentage of the chorion in intact eggs; 3)theFS50and the volume of intact eggs; 4)theFS50and the chorion thickness; 5)theFS50and the volume percentage of the chorion in intact eggs; 6)theFS50and the habitat salinity; 7)the habitat salinity and the dry weight of resting eggs. We suppose that the selection effect of salinity on the buoyancy of resting eggs (if any)may become observable only after an accumulation of enough generations. A directional evolution of resting egg buoyancy was observed after 20 years’ selective catching of floating eggs (Sura and Belovsky, 2016).Considering the data availability and the salinity fluctuation between different years/seasons, the mean of salinities recorded during the 30 years until the sampling time was adopted as the habitat salinity of each population. Original salinity data were obtained from literature or measured on site. When salinities of different stations were provided in a literature, their mean was used as the salinity value of the lake. In case salinities of different months/dates were provided in a literature, they were averaged first, and then the mean of different years was calculated. Due to the high variation of salinities among different ponds in an artificial salt production system (coastal solar salterns), and the unavailability of salinities for some inland populations (see Notes of Table 1), 9 populations were excluded from the last two analyses. Statistical analyses were performed using OriginPro 2018C, andPvalues calculated by Graph-Pad Prism Version 5.01. The difference was considered significance whenP< 0.05.
3 Results
3.1 Buoyancy of Resting Eggs
The left column of Fig.1 shows PF values at different salinities. For JZ, XT, JYH, BRK, GH, CN1 and WAS populations (The full names of the abbreviations can be referred in Table 1), the minimum salinity with some resting eggs floating was 20 (Figs.1A, C, F, I, L, N and V). It was 40 for CN2 population (Fig.1O), and 180 for DBC population (Fig.1J). For the remaining 16 populations, some eggs were floating in distilled water (Figs.1B, D, E, G, H,K, M, P – U, W – Y). The salinity at which all resting eggs were floating (i.e., PF = 100%)varied from 80 to 320, with the lowest value being recorded in YGH and ZD populations (Figs.1D, K), and the highest value in DBC population (Fig.1J). The change of PF with the increase of salinity varied among populations, and could be roughly classified into three patterns. 1)The PF increased at a constant speed (a linear increase), represented by JZ, DBC, GH and DXC populations (Figs.1A, J, L, T); 2)PF increased slowly under lower and higher salinities, while increased rapidly under median salinities (a S-curve increase), represented by XT, EBN, BRK, CN1, CN2, ALY, and WAS populations (Figs.1C, H, I, N – P, V); 3)PF increased rapidly under lower salinities, represented by HG, YGH, YC, JYH,AQK, ZD, CAK, GZ, KXC, LGC, GSL, EBT, BYL and BOR populations (Figs.1B, D – G, K, M, Q – S, W – Y).
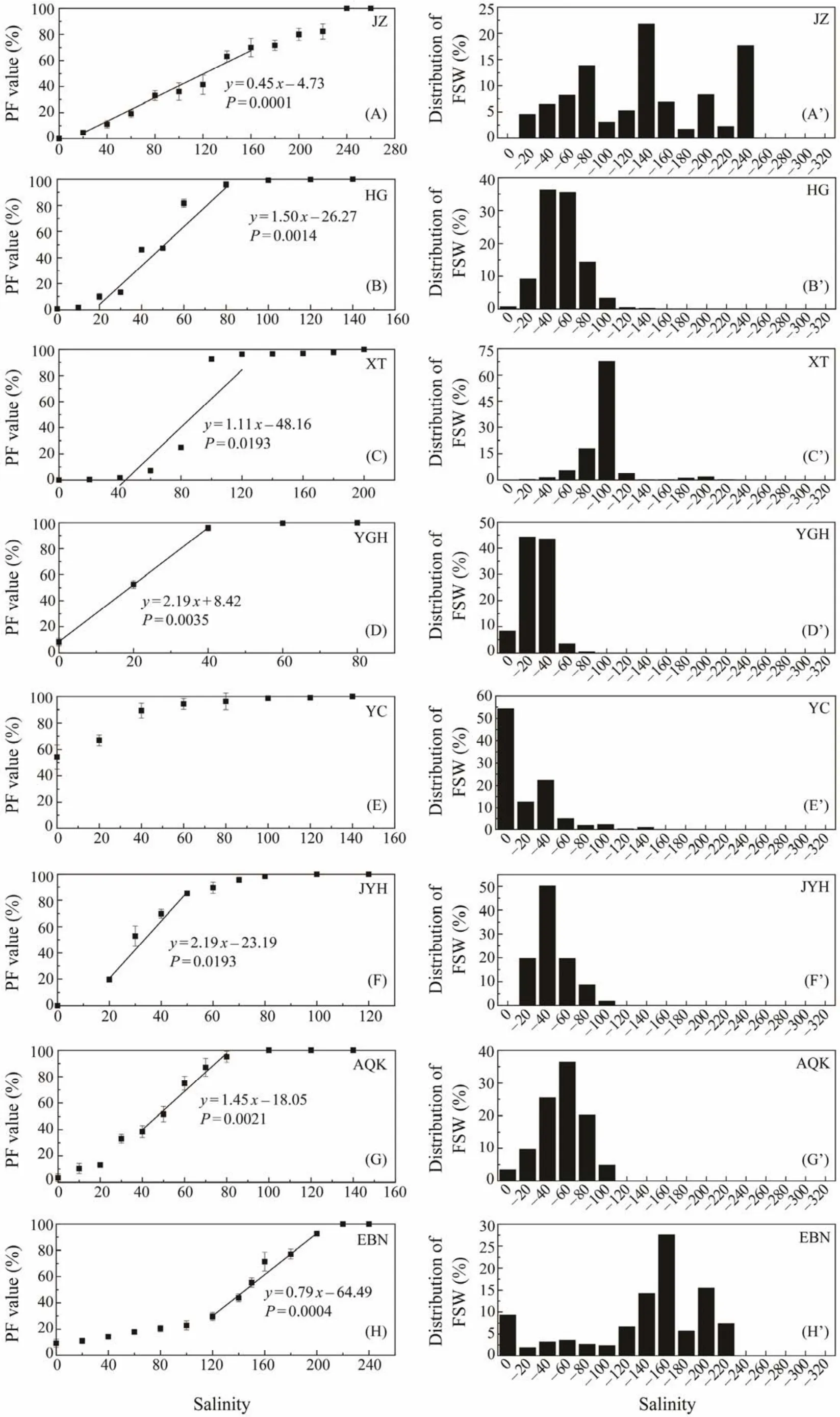
Fig.1 A comparison of the buoyancy of resting eggs from 25 different Artemia populations. Left column, percentage of floating resting eggs (PF)under different salinities (mean ± SD). Right column, distribution of the fraction of ‘apparent specific weight’ (FSW). ‘–x’ means a salinity interval between a neighboring lower salinity and salinity x, e.g., ‘–20’ means the salinity of 0 – 20 (to be continued).
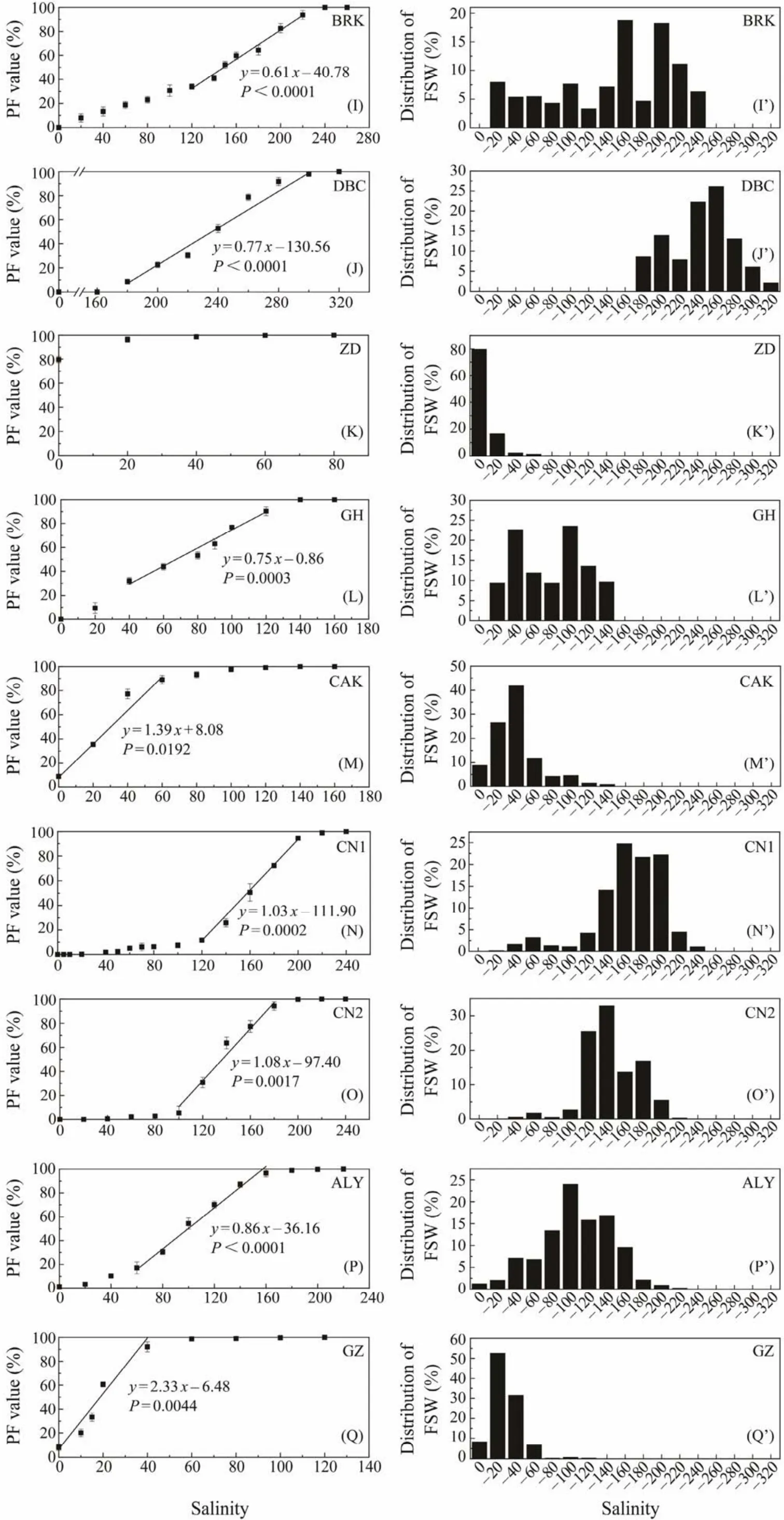
Fig.1 Continued 2 (to be continued).
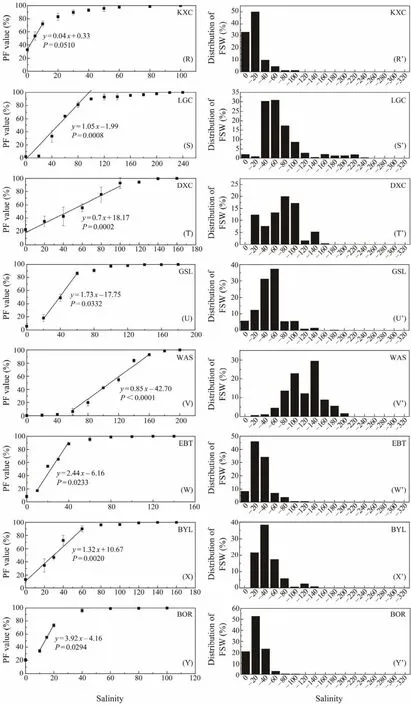
Fig.1 Continued 3.
In terms ofFS50(Table 2), the lightest resting eggs were represented by the populations ZD, YC (FS50not determined because over 50% resting eggs were floating in distilled water)and KXC (FS50= 4.3), while the DBC population had the heaviest resting eggs (FS50= 234.5). There were 14 populations (HG, XT, YGH, JYH, AQK, GH, CAK, GZ,LGC, DXC, GSL, EBT, BYL, BOR)with theirFS50values between 10 and 100, and 7 populations (JZ, EBN, BRK,CN2, ALY, CN1, WAS)with theirFS50values between 100 and 200.
As shown by the FSW values (Fig.1), the composition of the ‘apparent specific weight’ of resting eggs varied greatly among the studied populations. Eighteen populations (HG,XT, YGH, YC, JYH, AQK, ZD, CAK, CN1, CN2, ALY, GZ,KXC, LGC, GSL, EBT, BYL and BOR)exhibited a singlepeak distribution pattern (Figs.1B’ – G’, K’, M’ – S’, U’, W’– Y’). The other seven populations (JZ, EBN, BRK, DBC,GH, DXC, WAS)showed a multiple-peak or a non-peaked distribution pattern (Figs.1A’, H’ – J’, L’, T’, V’), and resting eggs of these populations usually had wider ranges in the distribution of ‘apparent specific weight’.
3.2 Biometrical Parameters of Resting Eggs
The largest diameter of intact eggs was recorded for GZ(356.8 µm ± 16.2 µm), followed by LGC (347.4 µm ± 22.7µm), AQK (325.4 µm ± 13.7 μm)and JYH (322.2 µm ±16.6 μm), while the smallest value was in YC (233.8 µm ±10.2 µm). For decapsulated eggs, LGC possessed the largest diameter (336.8 µm ± 21.7 µm), while the smallest diameters were found in YC (216.6 µm ± 12.8 µm)population(Table 2). Values of chorion thickness ranged from 1.5 µm(JZ)to 13.4 µm (JYH)(Table 2).
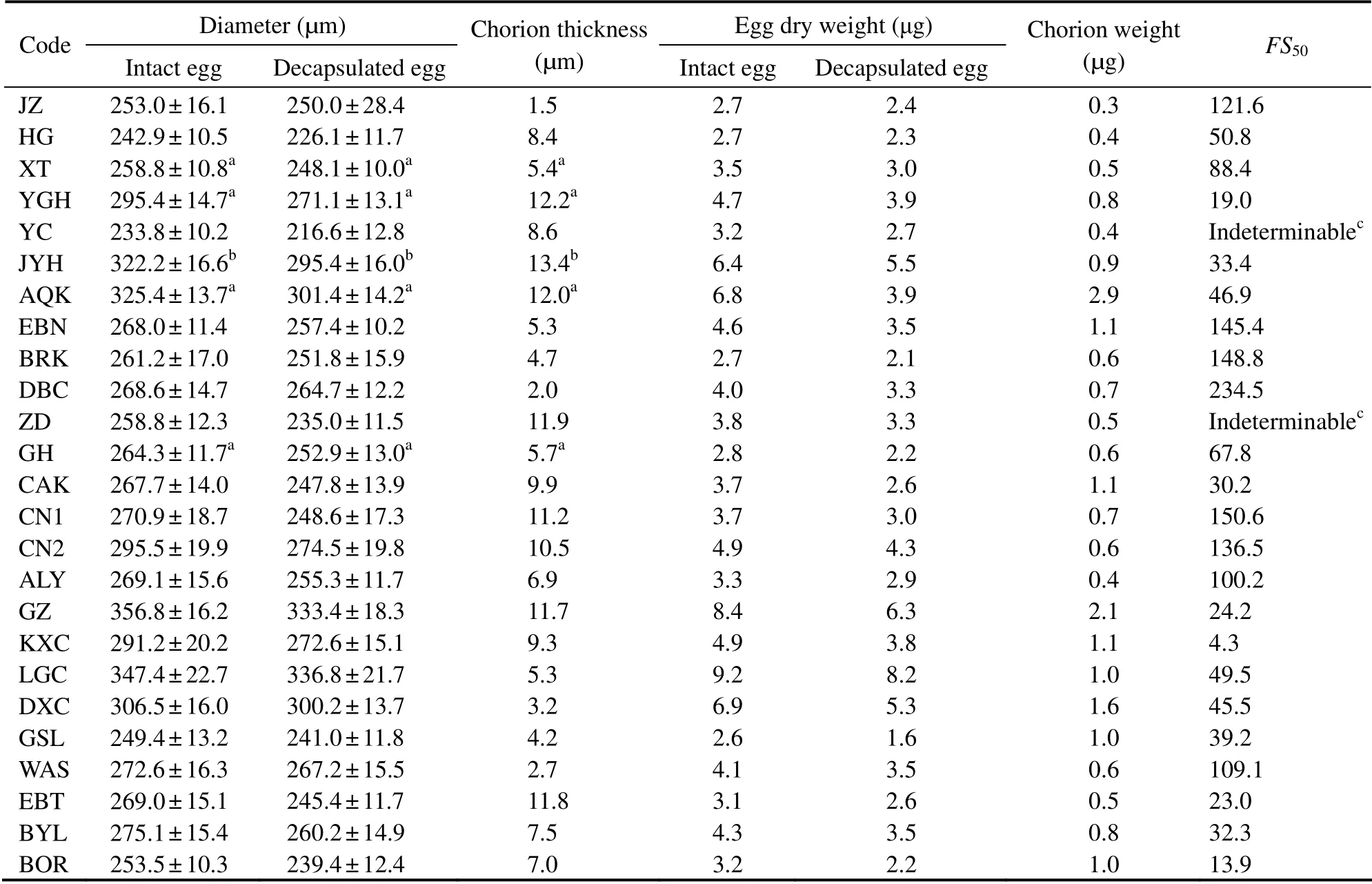
Table 2 Biometric values and FS50 for Artemia resting eggs from 25 localities
The average dry weight of intact eggs ranged from 2.6 μg (GSL)to 9.2 μg (LGC), that of decapsulated eggs varied from 1.6 μg (GSL)to 8.2 μg (LGC). The lightest chorion was recorded for JZ (0.3 μg), while the heaviest chorion was seen in AQK (2.9 μg)(Table 2).
3.3 Correlation Analysis
Correlation analyses did not show any significant correlation (P> 0.05)betweenFS50and the dry weight of intact eggs (Fig.2A), betweenFS50and the weight percentage of chorion in intact eggs (Fig.2B), as well as betweenFS50and the volume of resting eggs (Fig.2C). However,significant negative correlations were detected betweenFS50and chorion thickness (R2= 0.2486,P= 0.0112)(Fig.2D),and betweenFS50and the volume percentage of chorion in intact eggs (R2= 0.2660,P= 0.0083)(Fig.2E). Their relationship could be expressed byy= -8.40x+ 133.39 (Fig.2D)andy= -472.34x+ 141.47 (Fig.2E), respectively.
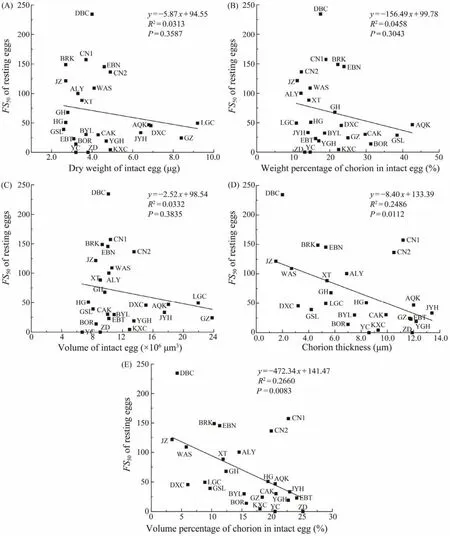
Fig.2 Relationships between FS50 and selected biometric parameters of Artemia resting eggs. The indeterminable FS50 of ZD and YC populations (see Table 2)are set to be 0.
A positive correlation was found betweenFS50and habitat salinity (R2= 0.2920,P= 0.0307). Their relationship could be expressed asy= 0.78x- 23.80 (Fig.3A). There seemed to be a negative relationship between habitat salinity and the dry weight of intact eggs, though the relationship is not statistically significant (P= 0.1121; Fig.3B).
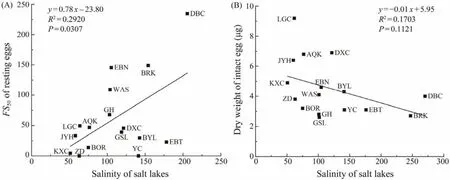
Fig.3 Relationships between habitat salinity and the FS50 of Artemia resting eggs (A), and between habitat salinity and the dry weight of intact eggs (B). The indeterminable FS50 of ZD and YC populations (see Table 2)are set to be 0.
4 Discussion
Present results show that the buoyancy or the ‘apparent specific weight’ ofArtemiaresting eggs varies greatly among different populations (Fig.1). TheFS50of ZD and YC population is not detectable because over 50% of resting eggs float in distilled water, while it is as high as 234.5 for DBC population (Table 2). Eimanifaret al. (2006)documented that floating eggs ofA. urmianahad higher haplotype diversity and nucleotide diversity than sinking eggs.Sura and Belovsky (2016)found that the percentage of floating resting eggs produced by floater parents (hatched from floating eggs)was 2.1 times higher than that of the eggs produced by sinker parents (hatched from sinking eggs).Both studies suggested that the buoyancy of resting eggs might have some kind of genetic component. Hence, the remarkable difference in the resting egg buoyancy among populations may be (at least partly)explained by the difference of their genetic basis. Moreover, among the 10 bisexual populations, only DXC (10.0%)shows a multi-peak distribution in the ‘apparent specific weight’ (Fig.1T’), while in the 13 parthenogenetic populations, 38.5% (EBN, BRK,DBC, GH and WAS)have a multi-peak distribution (Figs.1H’ – J’, L’, V’), suggesting that parthenogenetic populations seem to have higher diversity in the ‘apparent specific weight’ of resting eggs. This may be related to the different ploidies and different clones that each parthenogenetic populationhas. However, non-genetic factors may also affect the floating capacity of resting eggs,e.g., it can be affected by the food condition of parents (Sura and Belovsky, 2016; see Introduction).
For most (of the 16)populations with habitat salinity available (Table 1), there are some resting eggs that cannot float in their habitat salinity. For instance, percentages of resting eggs floating in their habitat salinity are 89.5%,95.1%, 23.0%, 52.1%, 22.5%, 76.8%, 95.7%, 64.0%, 94.4%,42.5% for JYH, AQK, EBN, BRK, DBC, GH, KXC, LGC,DXC and WAS populations, respectively (Figs.1F – J, L, R T, V). This is perhaps because these samples (except for BRK)were collected from coastal waters, where sinking resting eggs deposited in the sediment might have been suspended by waves and catching activities (DBC sample containing resting eggs that were forced to the shore by wind and waves). Resting eggs of BRK were collected from open waters with a boat, but might also contain eggs suspended from the sediment, because this lake is only 0.3 – 0.8 m in depth and is prevailing with northwest wind (Renet al.,1996). Resting eggs of YC, ZD, GSL, EBT, BYL and BOR were almost completely floating at their respective habitat salinity (Figs.1E, K, U, W – Y). Eggs of ZD, BOR, EBT and YC were collected along the shoreline. For these lakes,more than 99% eggs float at a lower-than-habitat salinity(60, 60, 140 and 140, respectively; Fig.1), suggesting that few resting eggs are sinking in their natural habitat. For GSL and BYL populations, unfortunately, we do not have the information about the collecting method of the samples.
No significant correlation is found between theFS50and the dry weight of resting eggs (Fig.2A), between theFS50and the volume of intact resting eggs (Fig.2B), and betweenFS50and the weight percentage of chorion in intact eggs(Fig.2C). On the contrary, a significant negative correlation is found betweenFS50and the thickness of chorion (Fig.2D),and betweenFS50and the volume percentage of chorion in intact eggs (Fig.2E). These results provide a statistical support to a previous conclusion that the buoyancy of resting eggs is largely determined by the chorion thickness. For instance, the absence of floating capacity in resting eggs ofA. monicais thought to be owing to their thin shell (Drinkwater and Crowe, 1991). Resting eggs ofA. franciscanahave a greater floating capacity than eggs ofA. urmiana, because the egg shell ofA. urmianahas a thinner alveolar layer than that ofA. franciscana(Sorgeloos, 1997; Abatzopouloset al., 2006).
Baitchorov and Nagorskaja (1999)reported a positive correlation between the dry weight of resting eggs and habitat salinity, but this is not supported by the present study(Fig.3B). The difference in results of these two studies is likely because different populations are studied. Samples of Baitchorov and Nagorskaja (1999)were collected from Crimea, Kazakhstan and Altai area, while the present study included some Qinghai-Tibet Plateau populations with altitudes over 4200 m above sea level. These high altitude populations (e.g., LGC, JYH, AQK)have remarkably bigger size and weight (Table 2). Since the habitat salinities of these populations are not higher than most otherArtemiahabitats, their large egg size is apparently not induced by high salinity. It is perhaps relative to their cold habitat climate (Van Stappenet al., 2003).
A recent study demonstrated that the selective harvesting against floating resting eggs in the Great Salt Lake had caused a decrease of resting egg buoyancy during 1991 to 2011 (Sura and Belovsky, 2016), suggesting that selective evolution of resting egg buoyancy could occur within a relatively short period. Present analysis shows a positive relationship betweenFS50and habitat salinity (Fig.3A). This seems to support the hypothesis that lower salinity may induce a selective evolution toward producing resting eggs with better floating capacity, while higher salinity induces an evolution toward producing heavier resting eggs. However, this hypothesis need to be further tested because of the following reasons. 1)The salinity of some lakes has considerable fluctuation among years/seasons, and for some lakes the existing salinity records are few and may be bias towards certain seasons/years. 2)In addition to genetic selection, phenotypic plasticity induced by the site salinity as well as other factors may also influence the buoyancy of resting eggs. 3)The resting egg bank of a population may be composed of both floating and sinking eggs, to what extent a resting egg sample can represent the natural egg bank may be dependent on the collecting method (see above).For DBC population, Renet al. (1996)reported that the salinity to float resting eggs was 140 – 175, while it was 180– 320 in the present study (Fig.1). Sampling methods may be a reason for such big difference in results of the two studies, though there may be also other reasons such as the change of salinity between sampling years. 4)A higher fraction of heavier resting eggs may be collected in a higher salinity season than that in a lower salinity season. For instance, Litvinenkoet al. (2016)found that at a salinity of more than 180 g L-1, the number of benthic resting eggs decreased and the number of planktonic resting eggs increased in some Siberian populations, which was probably due to the improvement of their buoyancy at high salinity. Unfortunately, it is almost impossible to collect samples from water body and/or upper sediment properly representing the resting egg bank of a lake, not only because the sediment may contain eggs deposited in years, but also because the distribution of floating resting eggs is very uneven at the water surface.
In conclusion, the floating capacity and the distribution pattern of the ‘apparent specific weight’ of resting eggs vary greatly among differentArtemiapopulations. The chorion of the resting egg is a major structure that provides the buoyancy of resting eggs. Habitat salinity seems to be an environmental factor inducing directional selection on the buoyancy of resting eggs, withArtemialiving in lower salinity waters tending to produce lighter resting eggs and vice versa. However, this hypothesis needs to be further tested by more intensive and ingenious samplings.
Acknowledgements
This study is supported by the Science and Technology Project of Tibet Autonomous Region (Nos. XZ201703-GB-04, XZ202102YD0022C), and the Science and Technology Project of Zhejiang Province (No. 2020C26008).We are grateful to Ms. Bakytgul Ospan, Mr. Jianbao Liu,Mr. Hongjun Zhang, Mr. Liwei Luan, Mr. Dian’an Zhang,Mr. Rongchang Tian and Mr. Pengfei Wang for providing samples of resting eggs and the collecting information of some samples. We thank Miss Ran Gao for her help in the experiment. We also thank Prof. Zengjun Du for clarifying the determination time of salinities reported by Sang (2018).
杂志排行

Journal of Ocean University of China的其它文章
- A Theoretical Model for the Microwave Emissivity of Rough Sea Surfaces
- Mechanism of Regional Subseasonal Precipitation in the Strongest and Weakest East Asian Summer Monsoon Subseasonal Variation Years
- Analytical and Experimental Studies on Wave Scattering by a Horizontal Perforated Plate at the Still Water Level
- Theoretical Prediction of the Bending Stiffness of Reinforced Thermoplastic Pipes Using a Homogenization Method
- Penetration Resistance of Composite Bucket Foundation with Eccentric Load for Offshore Wind Turbines
- Application of Converted Displacement for Modal Parameter Identification of Offshore Wind Turbines with High-Pile Foundation