金枝黄花柳回交子代枝条颜色与转录组研究
2022-12-24葛卉宋文龙刘兴菊梁海永
葛卉,宋文龙,刘兴菊,,梁海永
(1 河北农业大学 林学院,河北 保定 071000;2 河北省种质资源与森林保护重点实验室,河北 保定 071000)
柳属(Salix)植物种类多样,在世界各地都有广泛分布。全世界共有520余种,其中中国共有 257种[1]。柳属植物多为灌木或乔木,不仅用途广泛,而且具有长远开发前景,在环境保护方面具有净化空气、防风固沙、保持水土、修复被污染的土壤和修复水域的作用,在生活中可应用于城市园林绿化、营造木材林,作为生物质能源、编织品原材料、动物饲料等[2]。枝条是木本植物的重要观赏部位之一,尤其在冬季落叶后。目前,园林绿化常用的彩枝树种枝条分为红枝型和黄枝型,红枝以红瑞木、柽柳为主,黄枝型以金枝国槐、金丝垂柳为主[3]。柳树枝条颜色的变异极为丰富,最常见的有绿色、黄绿色和灰色,其他常见的还有黄色、红色、褐色等[4]。金枝黄花柳(Salixcapreavar.aurea)为自然变种,其枝条为金黄色,颜色亮丽,株型丰满,经引种驯化后,可应用于绿化观赏[2]。但是金枝黄花柳为雌株,种子飞散时会造成飞絮污染,金枝黄花柳半同胞家系存在广泛的形态变异[5]。枝条颜色由隐形基因控制,通过亲本杂交产生的子代回交会产生2种表现型,分别为金枝型与普通型,并产生雌雄株。
目前,对选育速生、耐盐、抗寒和抗旱的柳树已经有大量研究,但柳树作为园林绿化重要树种,在优良观赏性状方面的研究还很少[6-8]。本研究通过对金枝黄花柳回交产生的金枝型子代与普通型子代枝条颜色进行对比,并与亲本进行比较,研究其遗传特点,通过高通量测序、通路富集和功能注释,挖掘2种枝条的差异基因以及探讨色素合成的过程,为柳属育种亲本选配及早期选择指标筛选奠定理论基础,并为探索柳属回交子代苗木生长、枝条性状遗传变异规律及杂种优势提供了重要线索。尤其是北方冬季彩色树种较少,无絮彩色柳树的培育在一定程度上将改善北方园林绿化植物品种应用匮乏的现状,具有非常广阔的市场空间。
1 材料与方法
1.1 试验材料
试验材料取自于河北农业大学林学苗圃,位于E 115°26′47″,N 38°49′37″。该地形为平地,土壤类型为沙土,株行距为40 cm×50 cm。2020年3月对雄株(金枝黄花柳为母本、白皮柳为父本杂交产生的子代)与金枝黄花柳进行回交,回交子代枝条产生2种极端表现型,即金枝型与普通型。
1.2 试验方法
1.2.1 枝色测定(2021年5月) 选取2种回交子代枝条与亲本枝条测定枝条颜色,为了减少误差,均选择枝条的向光面。在统一自然光下,将白纸作为背景进行拍摄,用Photoshop调节白平衡,利用Photoshop中的取色器查看并记录调节白平衡后的枝条Lab值。L表示亮度,其值域为0~100,即从黑色到白色;a表示红绿程度,其值域为-128~127,即洋红到绿色的变化;b表示蓝黄程度,其值域为-128~127,即从蓝色到黄色[9]。
1.2.2 转录组测序(2020年8月) 取极端金枝型和极端普通型2种颜色枝条各30根,构建2个极端混池,将所选材料放入液氮罐内带回林木遗传育种实验室转移到-80 ℃超低温冰箱内,等待送样进行转录组测序。将各混样组送至北京百迈客生物科技有限公司进行DNA文库构建以及高通量测序。
1.2.3 数据分析 运用 Excel 2016、SPSS 23和Photoshop 2020软件对测得的试验结果进行数据分析并绘制出图形,分析金枝与普通枝条间的差异情况。利用百迈克云平台进行转录组数据及相关生物信息学分析。
2 结果与分析
2.1 金枝黄花柳杂交子代枝色比较
对枝条颜色进行定量分析,并做出色差图,金枝型及普通型子代枝条色差图,见图1;母本及父本枝条色差分布,见图2。

(a)金枝型
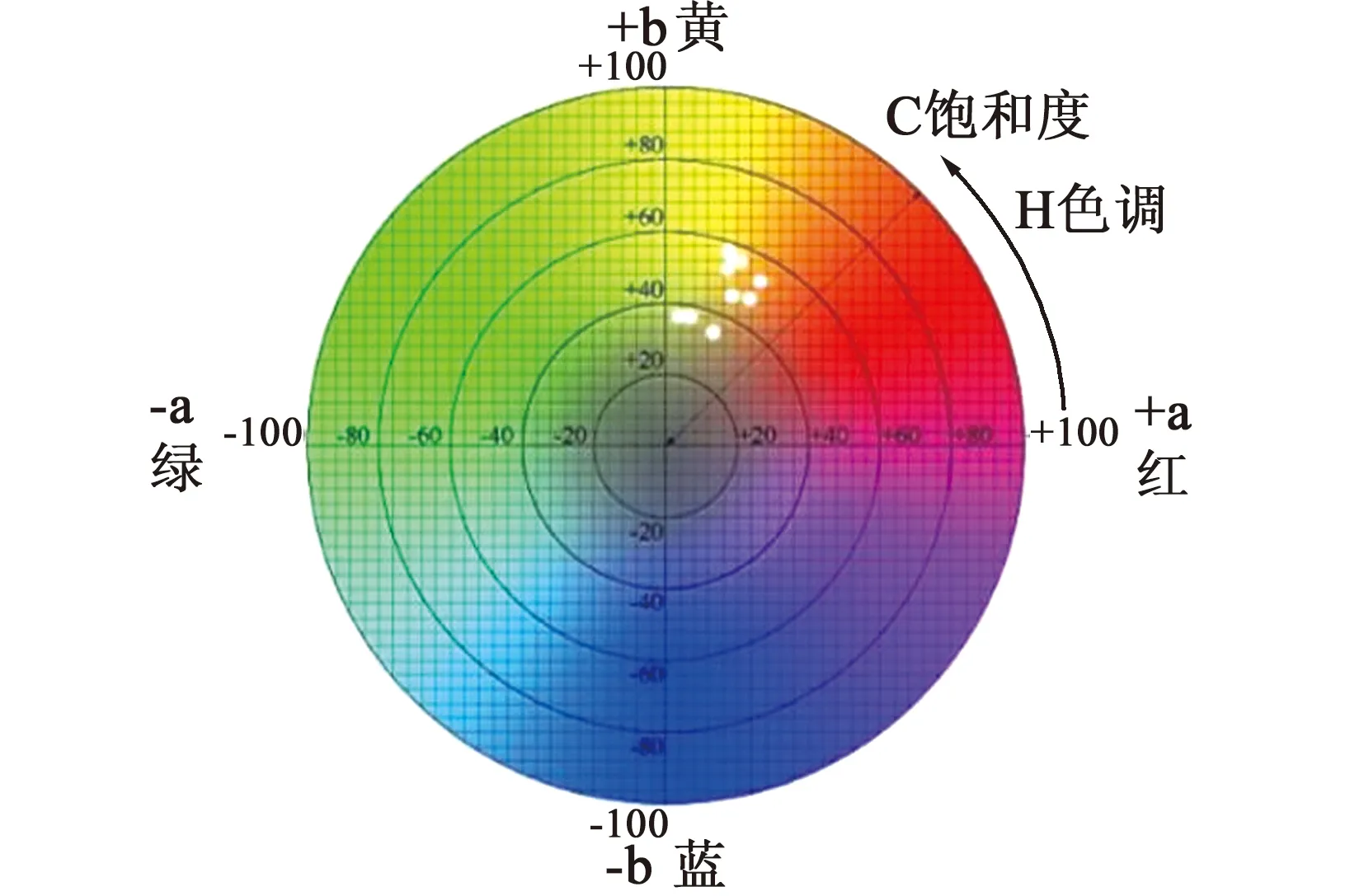
(a)母本
由图1可知,在回交子代植株枝条中,金枝型金枝黄花柳回交子代枝条表现为黄色到橙色,不同单株之间存在枝条颜色差异;普通型金枝黄花柳回交子代枝条表现为墨绿色至棕色,不同单株之间存在明显的枝条颜色差异。
由图2可知,金枝黄花柳回交子代群体与其父母本相比,金枝型子代枝条颜色与母本相差不多,普通型相较于父本来说更偏向于棕色。颜色性状属于数量性状遗传,一般具有连续变异的特点,金枝黄花柳经过杂交产生的子代并不表现为金枝,而子代回交后,出现金枝型与普通型2种性状,并且产生雌雄株[10-11]。
2.2 转录组测序分析
2.2.1 测序结果产出统计 基于高通量转录测序结果,对干净数据、总碱基数、GC含量和质量值≥30的碱基所占的百分比进行统计。测序数据统计表,见表1。

表1 测序数据统计表
由表1可知,运用高通量转录组测序技术,分别获得6.30 Gb和5.95 Gb有效数据(Clean data),试验样品中GC含量均超过45.12%,Q30碱基百分比均超过94.62%,可以用于进一步的转录组分析。
2.2.2 差异表达基因筛选 金枝与普通型枝条基因差异表达火山图,见图3。
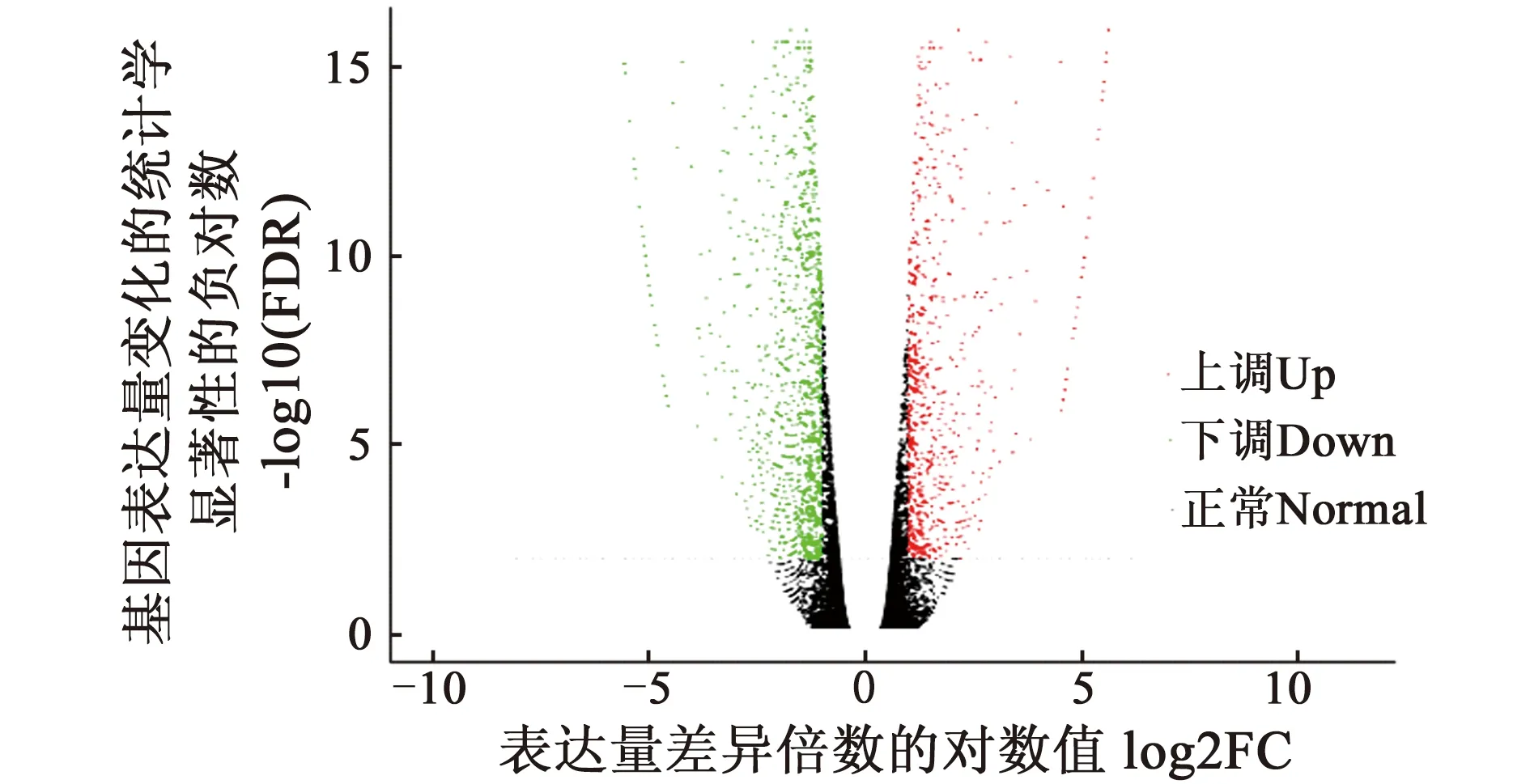
图3 金枝型与普通型枝条基因差异表达火山图
由图3可知,金枝黄花柳回交子代群体中金枝型枝条与普通型枝条相比,存在2 499个显著差异表达基因,其中上调基因数量984个,占差异表达基因总数的39.38%;下调基因数量1 515个,占差异表达基因总数的60.62%,上调表达基因数量明显少于下调表达基因数量。
2.2.3 差异表达基因 GO 功能富集分析 差异表达基因GO注释分类统计图,见图4。
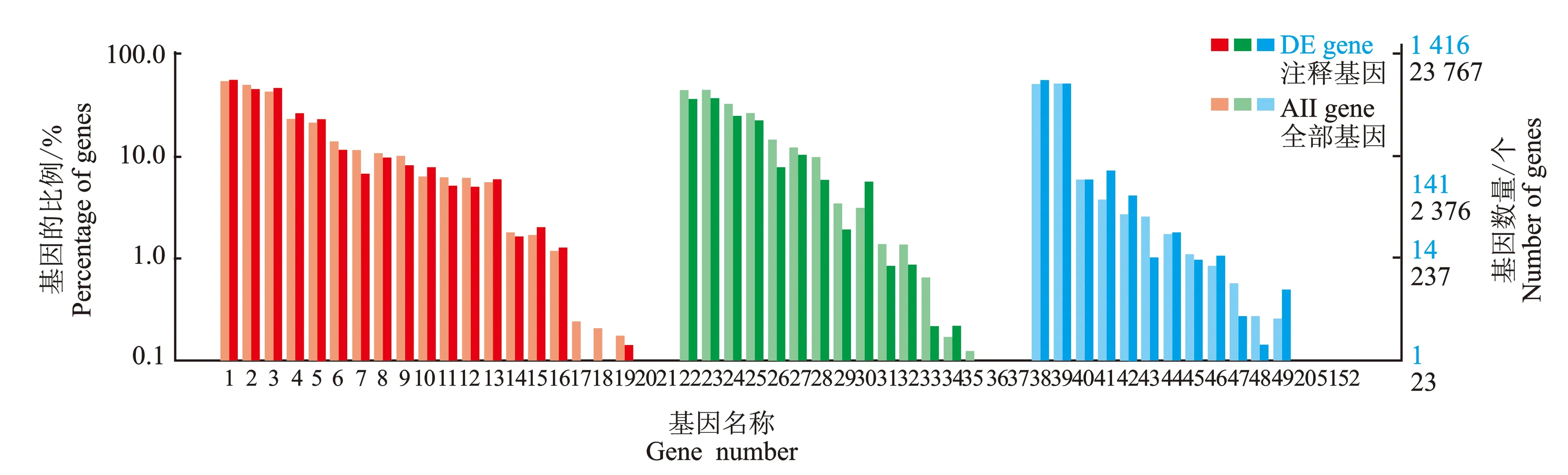
图4 差异表达基因GO注释分类统计图
由图4可知,通过GO数据库注释,共获得1 416个差异表达基因,其中上调540个,下调876个,分别包括GO系统3个主要分支,即:生物学过程(1-21),细胞组分(22-37)和分子功能(38-52)。回交子代金枝型与普通型枝条在生物学过程的21个亚分组(代谢等)中,在代谢过程、细胞过程、单一生物过程中得到注释的序列最多,仅有极少数注释到节律过程。在构成细胞组分的16个亚分组(细胞器部分等)中,细胞、细胞部分注释的差异表达基因最多,超分子复合物和细胞外区部分注释到的序列最少。在分子功能的15个亚分组(催化活性等)中,回交子代金枝枝条与普通型枝条对催化活性、结合活性注释到的基因序列多,在转录因子活性和蛋白质结合活性中序列少。
2.2.4 差异表达基因COG分类分析 利用COG数据库对基因产物进行直系同源分类,差异表达基因 COG 功能富集分析见图5。
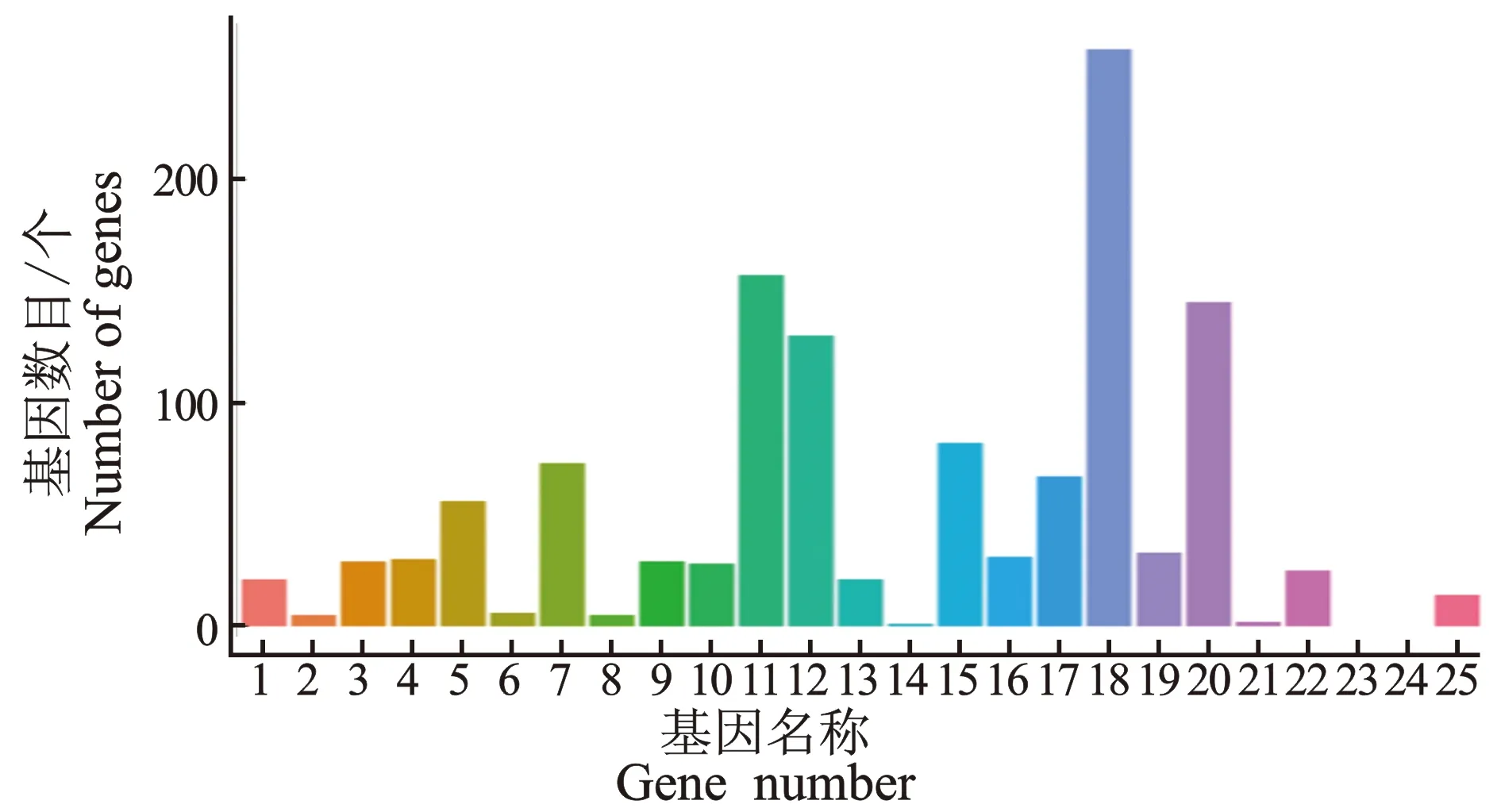
图5 差异表达基因 COG 功能富集分析
由图5可知,样品组之间差异表达基因COG分类统计结果显示,通过COG功能富集分析,共注释到750个差异基因在不同功能类中,其中上调277个,下调473个。在一般功能预测基因中,存在256个差异表达基因,其次是转录通路157个,信号转导机制通路145个,复制、重组和修复通路130个,在翻译后修饰、蛋白质周转和伴侣,糖转运和代谢,次级代谢物的生物合成、运输和分解代谢,氨基酸转运和代谢通路中也有较多富集。
2.2.5 差异表达基因KEGG通路富集分析 差异表达基因KEGG通路富集散点图,图中呈现了显著性Q值最小的前20个通路,见图6。
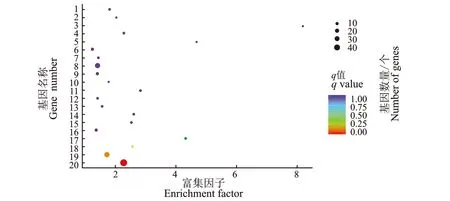
图6 差异表达基因KEGG通路富集散点图
由图6可知,利用KEGG分析差异表达基因在某一通路上是否存在显著差异,由KEGG 注释到的差异基因数目为753个,其中上调323个,下调430个。回交子代2类枝条样本在植物-病原体相互作用通路富集的基因数最多且显著性明显,在内质网蛋白质加工和酪氨酸代谢通路富集的基因数较多,在牛磺酸和次牛磺酸代谢中也有一些差异表达基因。
基因的KEGG富集结果,见表2。
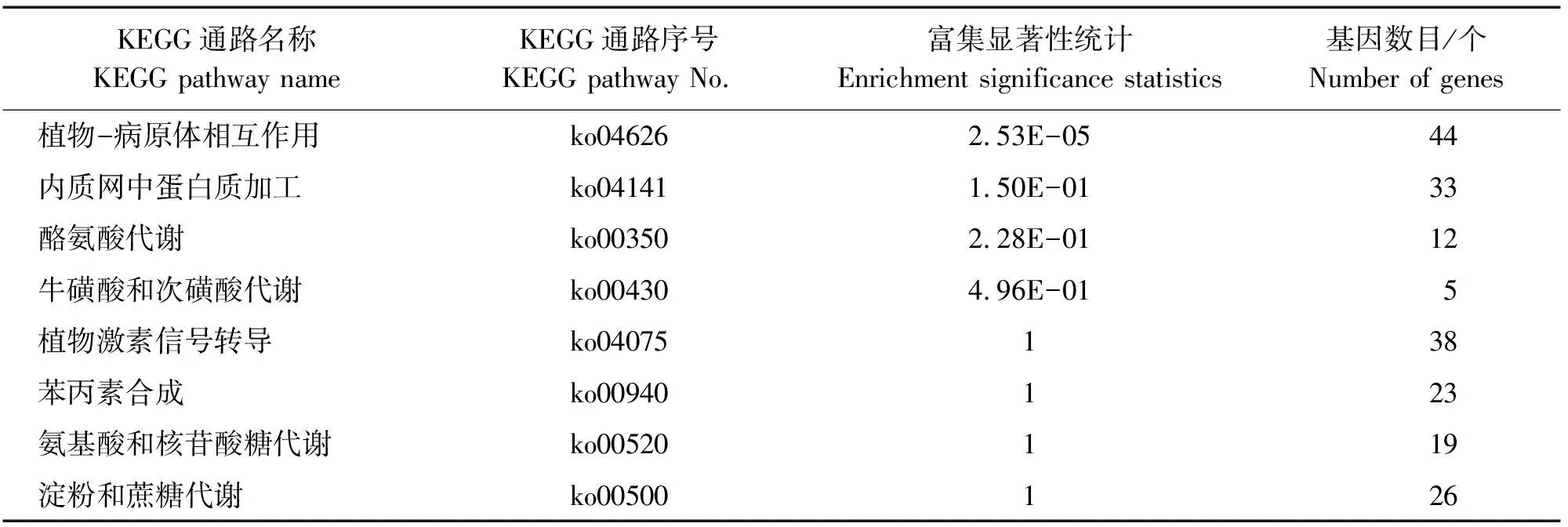
表2 基因的KEGG富集结果
由表2可知,回交子代2类枝条样本在植物-病原体相互作用通路富集的基因数最多且显著性明显,为44个差异表达基因,占KEGG通路富集的10.38%,在内质网蛋白质加工和酪氨酸代谢通路富集的基因数较多,分别为33个和12个差异基因,在牛磺酸和次牛磺酸代谢中也有一定显著性,注释到5个差异表达基因,在植物激素信号传导、淀粉和蔗糖代谢、苯丙素合成、氨基酸和核苷酸糖代谢通路中也有较多基因富集,但差异不显著。其余富集通路的差异基因比例为 0.94%~3.77%。
2.2.6 叶绿素合成与代谢途径中相关差异表达基因 由于枝条中色素表达产生差异,金枝黄花柳回交子代群体枝条产生不同颜色,因此,对叶绿素合成与代谢途径相关差异表达基因进行筛选。叶绿素合成与代谢相关通路注释结果,见表3。
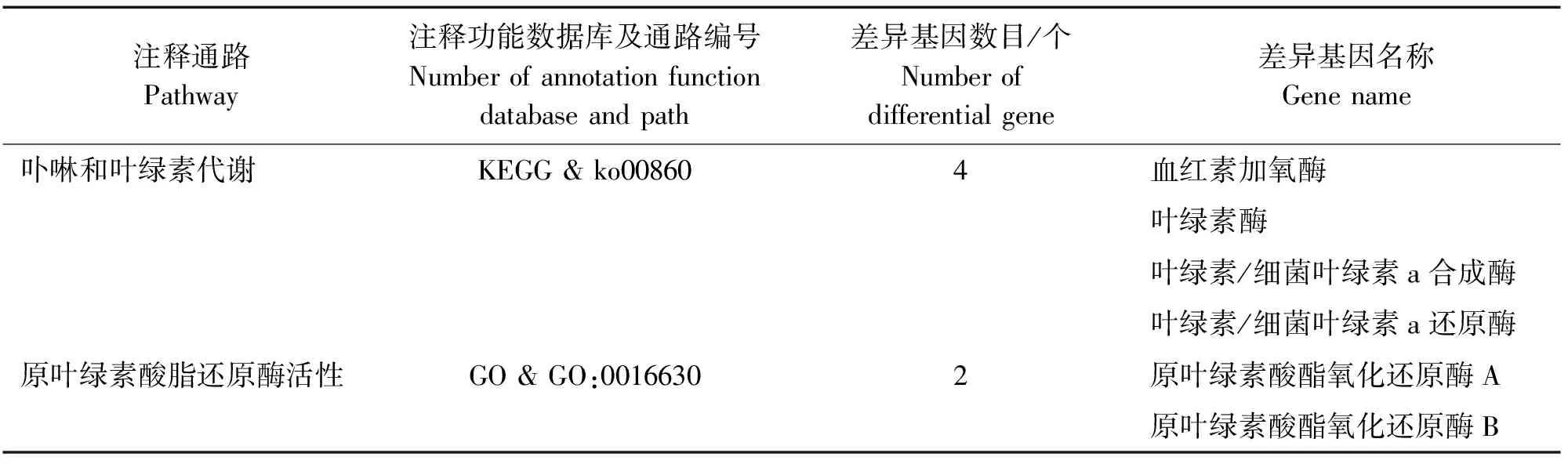
表3 叶绿素合成与代谢相关通路注释结果
由表3可知,通过KEGG注释分析通路,筛选出参与卟啉和叶绿素代谢通路(ko00860)的差异基因共有4个,分别是叶绿素/细菌叶绿素a合成酶、叶绿素酶、叶绿素/细菌叶绿素a还原酶和血红素加氧酶。通过GO注释分析,发现参与分子功能中原叶绿素内酯还原酶活性通路(GO:0016630)的差异基因有2个,分别是原叶绿素酸酯氧化还原酶B和原叶绿素酸酯氧化还原酶A。叶绿素合成与代谢相关通路差异基因表达情况,见表4。
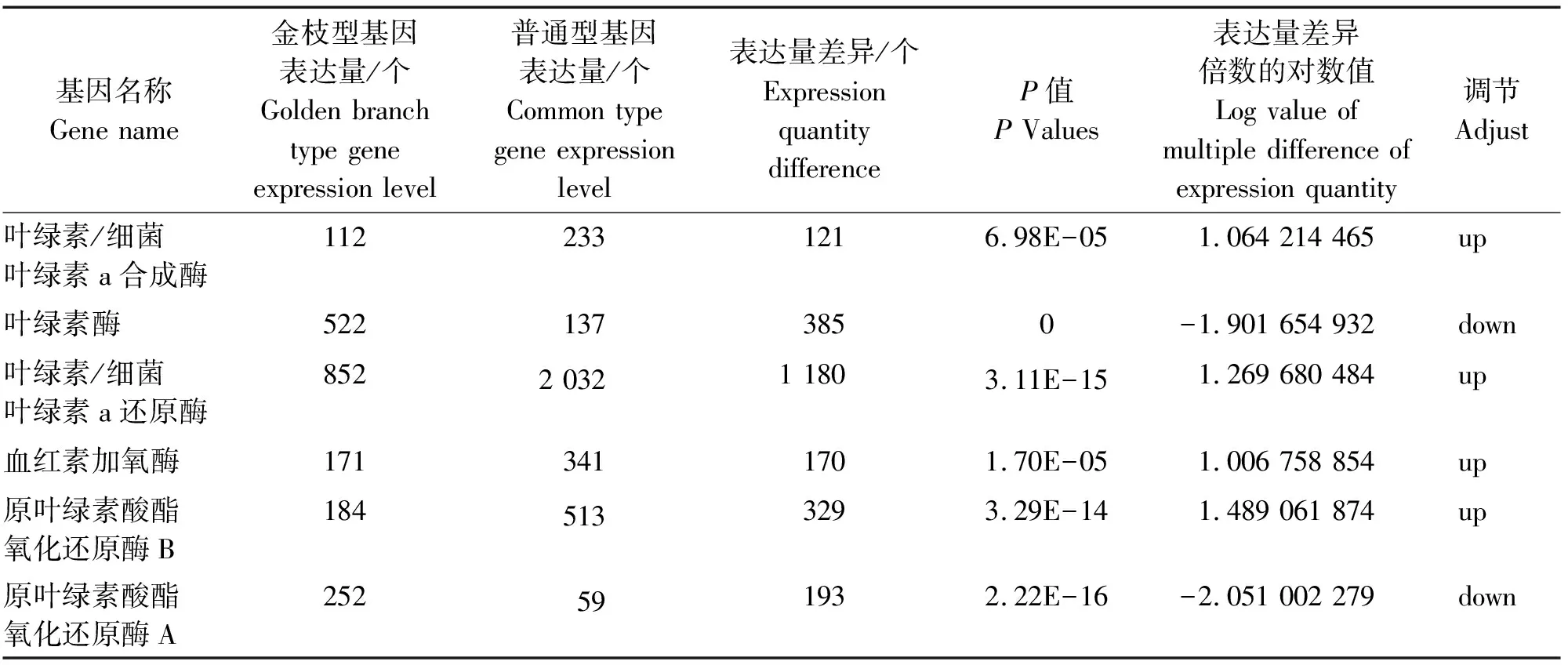
表4 叶绿素合成与代谢相关通路差异基因表达情况
由表4可知,6种差异基因中有4个上调基因,2个下调基因。上调基因中,叶绿素/细菌叶绿素a还原酶差异基因表达量最多,有1 180个;其次是原叶绿素酸酯氧化还原酶B差异基因,有329个;血红素加氧酶差异基因,有170个;叶绿素/细菌叶绿素a合成酶差异基因数量最少,有121个。下调基因中,叶绿素酶差异基因较多,有385个;原叶绿素酸酯氧化还原酶A差异基因数量较少,有193个。
2.2.7 卟啉与叶绿素代谢通路分析 绿色植物的叶绿素合成需要多种酶的参与,是一个非常复杂的过程。从谷氨酰-t RNA(Glu-t RNA)到合成叶绿素,由16种酶20多个基因编码的完成[12]。此过程中任何一种基因表达发生差异,都有可能对叶绿素的合成产生影响。卟啉与叶绿素代谢通路,见图7。
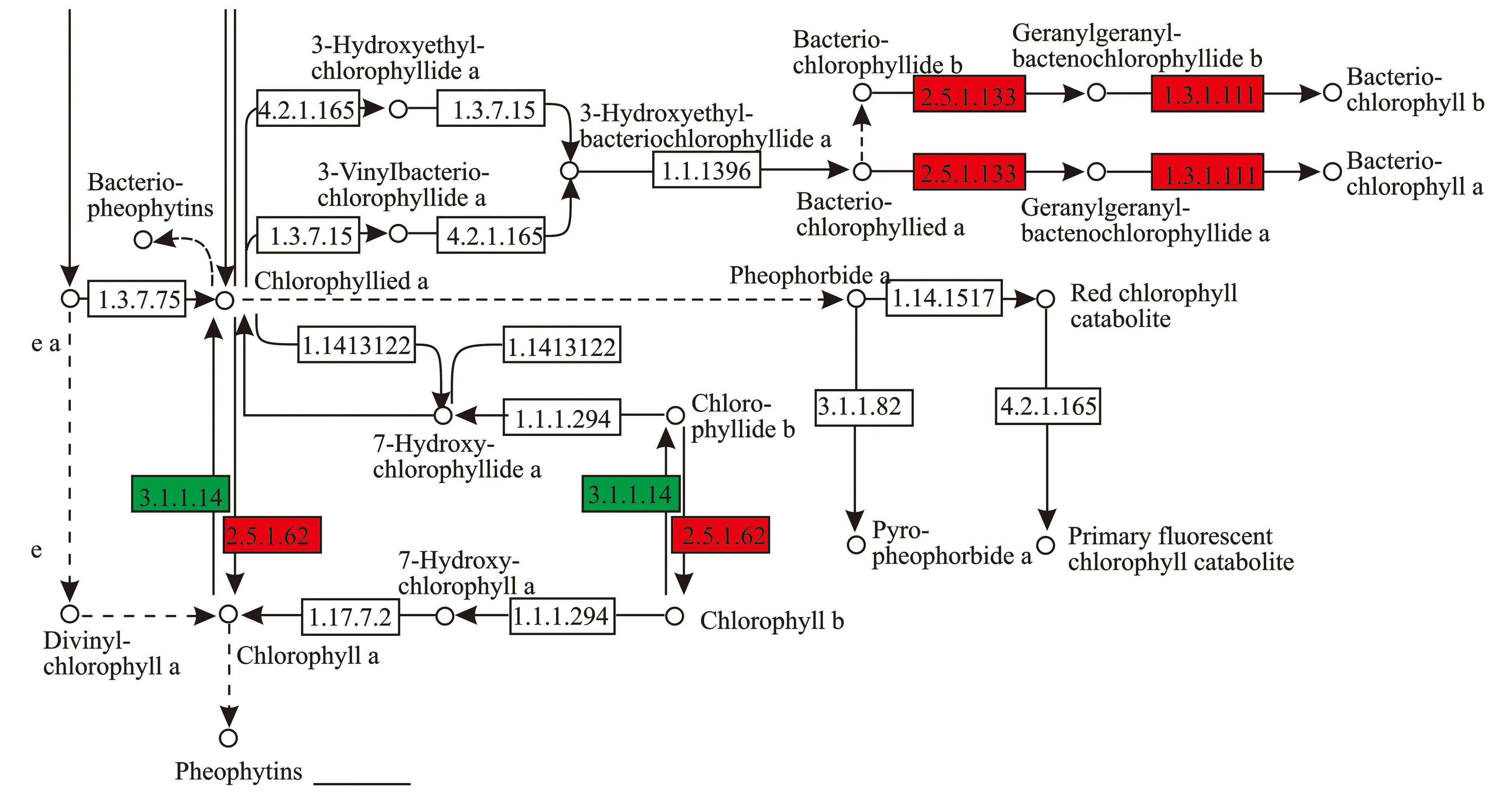
图7 卟啉与叶绿素代谢通路
由图7可知,与金枝型枝条相比,普通型枝条的叶绿素/细菌叶绿素a合成酶(2.5.1.133)与叶绿素/细菌叶绿素a还原酶(1.3.1.111)基因表达上调,因此,普通型细菌叶绿素a和细菌叶绿素b含量比金枝型多。叶绿素/细菌叶绿素a合成酶(2.5.1.62)基因表达上调,叶绿素酶(3.1.1.14)基因表达下调,二者共同通过反馈作用共同调节叶绿素a与叶绿素b的表达量。
3 讨论与结论
3.1 讨论
高通量转录组测序技术可以发掘植物潜在的功能基因、探索植物体内的生理调控机制、形态差异产生机制,发掘参与植物生长发育相关基因,以及抗病、抗逆等优良性状相关的基因,分析杂种优势的分子机理等,为进一步了解基因的作用机理打下基础[13-18]。在关于柳属树种转录组测序技术应用中,郑纪伟以垂柳和簸箕柳为研究材料,比较乔木柳与灌木柳转录本之间的差异,结果发现,利用乔木柳开发的引物在乔木柳不同种及种间的通用性更高,而利用灌木柳开发的引物在灌木柳不同种及杂种间的通用性更高[19];刘菁菁以簸箕柳雌、雄花序为材料,通过高通量测序与生物信息分析,确定这2种花序中的差异基因和表达差异基因[13]。通过对金枝型与普通型2个样品的转录组分析,发掘新基因1 705个,其中1 430个得到功能注释。基于比对结果,进行基因表达量分析。根据基因在不同样品中的表达量识别差异表达基因,并对其进行功能注释和富集分析。
木本植物枝条的树皮、射线薄壁组织、韧皮部、木质部甚至髓中都可发现叶绿体,尤其是幼嫩枝条中[20]。本研究发现,与叶绿素合成有关的卟啉,叶绿素代谢通路和原叶绿素酸脂还原酶活性通路中基因表达存在差异。卟啉环是叶绿素核心部分,叶绿素合成受多种酶催化和某些转录因子调控[21]。通过KEGG注释通路分析,筛选出参与卟啉和叶绿素代谢通路(ko00860)的差异基因共有4个,其中叶绿素/细菌叶绿素a合成酶、叶绿素/细菌叶绿素a还原酶和血红素加氧酶基因表达上调,叶绿素酶基因表达下调。在有光条件下,原叶绿素酸酯还原酶在植物显绿过程中起重要作用[22]。叶绿素酸酯氧化还原酶的作用是在光照条件下,催化原叶绿素酸酯形成叶绿素酸酯。原叶绿素酸酯氧化还原酶A基因易受光照影响,在黄化组织中特异性表达,参与构成黄化质体中原片层体,而原叶绿素酸酯氧化还原酶B组成型表达在2种类型植物中并不受光照条件影响[23]。通过GO注释通路分析,发现参与分子功能中原叶绿素内酯还原酶活性通路(GO:0016630)的差异基因有2个,其中原叶绿素酸酯氧化还原酶B基因表达上调,原叶绿素酸酯氧化还原酶A基因表达下调。
3.2 结论
本研究以金枝黄花柳回交子代及其亲本为材料,发现子代枝条主要表现金枝和普通型2种类型。金枝型子代枝条表现为黄色到橙色,普通型回交子代枝条表现为墨绿色至棕色。通过回交,子代重新获得具有与母本相似的金枝性状,且产生雄株。未来可在园林绿化进行应用。
通过RNA转录组测定及分子生物学分析,对比不同表型金枝黄花柳回交子代相关基因表达谱的不同。结果显示,枝条转录组共检测到差异表达基因2 499个,其中上调基因984个,下调基因1 515个。功能注释和富集分析发现,与色素合成有关的基因表达量存在差异,部分基因会影响代谢通路,进而影响枝条中叶绿素的含量,导致枝条颜色存在差异。通过差异表达基因通路发现,金枝型枝条细菌叶绿素含量低于普通型枝条,枝条呈现金黄色,与原叶绿素酸酯氧化还原酶A基因上调表达有关,对光照更敏感。叶绿素酸酯氧化还原酶通路中的叶绿素酸酯氧化还原酶A(porA)基因,在金枝型中发生特异性表达。
