大盈江捕食线虫真菌空间分布格局
2022-12-21朱娅佼杨晓燕
张 欣,邓 巍,朱娅佼,杨晓燕,肖 文,李 娜*
(1.大理大学东喜玛拉雅研究院,云南大理 671003;2.云南省高校洱海流域保护与可持续发展研究重点实验室,云南大理 671003)
微生物多样性分布与生态系统的调节过程、生态系统功能、生态系统恢复关系密切,因此微生物分布格局及其维持机制一直是生态学研究的热点〔1-3〕。早期的微生物学家认为微生物的分布与地理隔离无关,微生物在环境中呈随机分布模式〔4-5〕。近年来,随着生态学研究的不断发展,越来越多的证据表明微生物与地理历史关系密切〔6〕。在多种影响因素的共同作用下,微生物存在着复杂的空间分布格局〔7-8〕。如季节变化、气候条件、海拔、微生物特性以及人为干扰等因素均会影响微生物的空间分布〔9-11〕。自然界中的微生物不仅与非生物环境相互作用〔12〕,还受动植物分布〔13〕、研究区域环境异质性〔14〕的影响,因此微生物多样性分布格局的研究相较动植物更为困难,而选择一个合适的研究区域开展相关研究也尤为重要。
流域是一个具有清晰边界和相对独立的生态系统〔15〕,在分隔陆地系统的同时也驱动了物质和能量在水陆生境和上下游的分布。有学者提出微生物多样性在流域视角下是不均匀分布的〔16〕,随机过程在河流微生物群落形成中起着更重要的作用〔17〕。河流的地理阻隔和连通性也会塑造微生物空间分布格局〔18〕。在流域视角下可更好地预测微生物多样性〔19〕。目前关于微生物多样性分布格局等研究多集中在海洋和湖泊环境〔20-21〕,在流域生态系统的研究相对较少。基于流域开展微生物空间分布格局的研究,必须关注空间尺度,如流域内的上下游、水陆生境、干支流等各个流域单元内微生物的空间分布差异。研究〔16〕表明,微生物在大的流域尺度下具有明显的空间地理分布特征,也有研究〔22-23〕证明了微生物在流域的水陆生境间具有异质性分布格局,且同一条流域干支流上的微生物具有相似的水陆分布格局。因此,选择在一条小型河流的上下游初步探究微生物的空间地理分布是逐步明晰流域视角下微生物空间分布格局的关键。然而微生物种类众多,且大多为人类未知,尽管高通量测序技术的不断发展能够帮助人们获得更多的微生物群落信息,但也仍然没有办法理解整个微生物类群的空间异质性分布〔24〕。而结合纯培养手段,选择一类物种和分类信息相对完善、生态功能明确的微生物类群来开展相关研究也为微生物生态学研究提供了一种新的思路。
捕食线虫真菌(nematode-trapping fungi,NTF)是一类水陆两栖并广泛分布的真菌类群。该类群既可以营腐生生活又可以通过营养菌丝特化形成捕食器官来捕捉线虫,目前仅有116种〔25-31〕,容易鉴别,是利用纯培养手段开展微生物生态学研究的理想类群。因此,该类群是研究微生物分布格局的理想对象〔27〕。本研究选择生物多样性丰富、环境异质性强的大盈江作为研究区域,以NTF为研究对象,探究NTF在流域上下游的空间分布格局,同时验证水流对微生物分布具有汇聚作用的假说。研究结果将为在流域视角下开展微生物空间分布格局研究提供数据支持。
1 材料与方法
1.1 研究区域 大盈江又称大车江,全长约204.5 km,流域面积约5 636 km2,是云南六大水系之一的伊洛瓦底江一级支流,具有云南重要的河流湿地类型,地处热带和南亚热带的过渡地带〔32〕。其特殊的地理位置和气候条件以及复杂的地形地貌,孕育了大盈江的生物多样性。本研究选择大盈江为研究区域,沿着其河流流向进行样区布设,根据河流长度将其划分为上、中、下游。
1.2 样品采集 研究区域内,共布设3个样区,每个样区处以河流交汇点为支点,在支点及支点上、下游10、20 m处设置样点,每个样点处对称采集水体底泥和河岸陆地土壤样品,共获得3个样区、30个样点处的30份样品。陆地土壤样品采集方法:按5点采样法采集土壤表层以下0~10 cm的土壤约50 g,将得到的样品充分混匀;水中底泥采集方法:选用自制取样设备采集立下0~10 cm的底泥约50 g,每份样品通过5点采样法混匀得到。将采集到的样品分别置于一次性无菌自封袋中,做好样品信息标记,带回实验室,1周内完成所有样品的处理。
1.3 培养基配制 参照文献〔27〕,配制玉米粉琼脂培养基(corn meal agar medium,CMA),用于分离纯化NTF;配制马铃薯葡萄糖琼脂培养基(potato dextrose agar medium,PDA),用于NTF的菌丝富集;配制燕麦培养基,用于诱饵线虫的培养。
1.4 诱饵线虫的培养和悬浮液的制备 将全齿复活线虫接入燕麦培养基放恒温培养箱(26.5±1.0)℃培养1周,观察到线虫爬壁,取出备用或放4℃冰箱保存。采用贝尔曼漏斗法〔27〕制备线虫悬浮液。
1.5 NTF的分离纯化 分离纯化采用诱饵平板法〔27〕:用5点撒样法在CMA上撒入1.5 g土样,加入线虫悬浮液2 mL(约5 000条)以诱导NTF生长,每份样品设置3个平行,将平板放置于常温下培养4周,用体视显微镜镜检。观察到NTF后,用无菌牙签将NTF单个分生孢子转接到CMA上,将接种好的CMA置于恒温培养箱中26.5℃培养,培养出的NTF纯化3~5次。
1.6 NFT物种鉴定NFT形态特征观察:用无菌刀在CMA上挖取1个2 cm×2 cm的观察室,在观察室边缘45°角斜插入洁净的盖玻片,待分生孢子和菌丝铺满盖玻片,用小镊子夹出盖玻片,在一片洁净的载玻片上(靠正中间位置)滴加1滴无菌水,覆上盖玻片,制成临时装片,在显微镜(40×)下观察〔33〕,用奥林巴斯BX53微分干涉显微成像系统拍取分生孢子、孢子梗、厚垣孢子等形态特征。
NFT捕食器官诱导:待菌丝长满观察室,即可滴加诱饵线虫悬液(约200条线虫),12~24 h后,用体式显微镜(10×或40×)观察培养皿上形成的各种捕食器官类型;结合菌株形态特征及捕食器官类型进行形态学鉴定〔27〕。
NFT的分子鉴定:在PDA上进行NFT菌丝富集培养后,参照文献〔34〕进行DNA的提取,以真菌通用引物ITS1/ITS4〔35〕和RPB1-DF2asc〔36〕/RPB1-G2R〔37〕分别扩增核糖体RNA上的非转录间隔区和RNA聚合酶Ⅱ核心亚基编码基因序列。获得测序结果后,在美国国家生物技术信息中心NCBI(https://www.ncbi.nlm.nih.gov/)中通过上述序列同源性比对进行分子生物学鉴定,结合菌株形态特征进行最终的物种鉴定。
1.7 数据处理 计算物种的检出率,检出率(occurrence frequency,OF)=(物种出现的样品数/总的样品数)×100%〔33〕。
采用香农-维纳多样性指数(Shannon Wiener's diversity index,H′)计算上、中、下游各样区的物种多样性:,式中,S为物种数目,N为物种的个体数之和,Ni为第i个物种个体数量。
采用Jaccard指数计算各样区间的群落相似性,Jaccard指数=a/(a+b+c)。式中,a表示2个样点均出现的物种数,c表示仅在样点1出现的物种数,b表示仅在样点2出现的物种数。Jaccard指数值越大,表示物种相似度越高。
2 结果
2.1 大盈江NTF物种分布 大盈江30份样品中共分离鉴定出1属9种19株NTF,检出率为43.33%,物种多样性指数为1.91。其中上游1属3种,中游1属4种,下游1属6种。弯孢节丛孢Arthrobotrys.musiformis在上、中、下游均有分布;秀丽单顶孢A.oudemansii分布在上、中游;爪哇节丛孢A.javanica分布在中、下游;而少孢节丛孢A.oligospora仅在上游有分布;厚皮单顶孢A.eudermata仅分布在中游;奇妙单顶孢A.thaumasium、中华单顶孢A.sinensis、狂带单顶孢A.rutgeriense和圆锥节丛孢A.conoides仅在下游有分布。见图1。
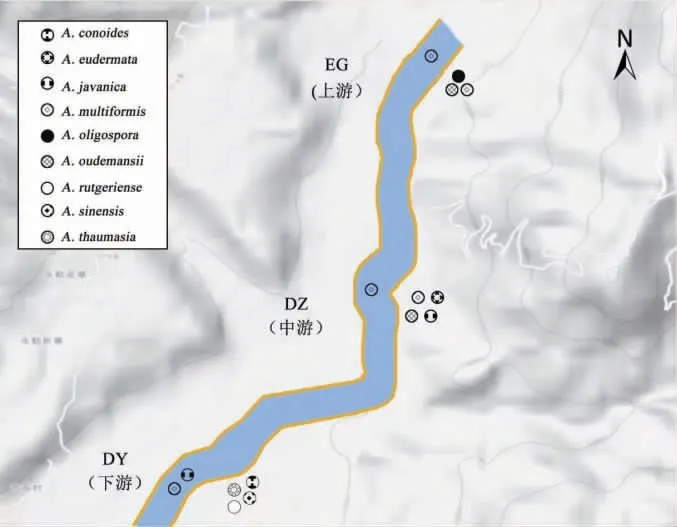
图1 大盈江上、中、下游样区NTF物种分布图
2.2 大盈江上、中、下游NTF物种检出率和多样性对大盈江上、中、下游的NTF检出率和香农-维纳多样性指数进行计算。结果显示,下游NTF的检出率最高,中、上游检出率相同;香农-维纳多样性指数为下游最高,中游次之,上游最低。见表1。
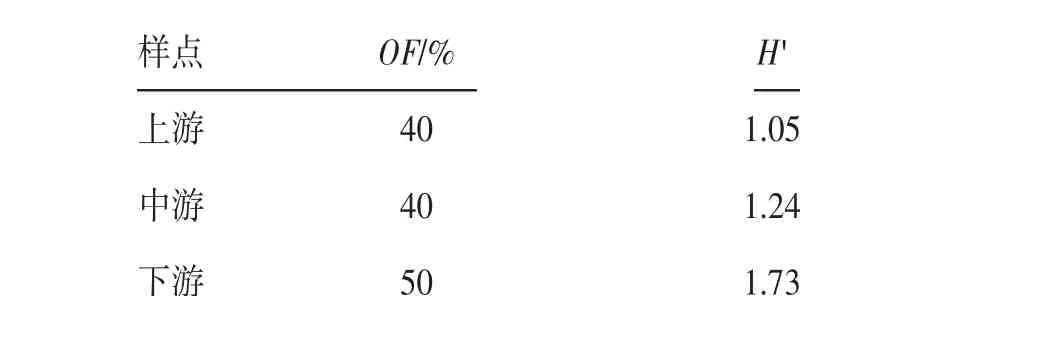
表1 大盈江上、中、下游样区NTF检出率和香农-维纳多样性指数
2.3 大盈江各样区间的NTF群落相似性 分别对大盈江上中游、中下游、上下游的水体底泥和陆地土壤的Jaccard指数进行计算,结果发现:NTF在上中游、中下游、上下游的水体中物种相似性均高于陆地。见表2。

表2 大盈江各样区间NTF Jaccard指数
3 讨论
在一个通过水流进行联通的流域生态系统中,微生物在流域内的分布在理论上应该是均质的。但本研究结果显示,NTF在大盈江流域内存在上下游异质性空间分布格局。在大盈江分离到的NTF中,只有弯孢节丛孢A.musiformis在各个样区均有分布,其余物种仅在局部样区分布,且沿着河流的方向,区域特有物种逐渐增多,NTF物种多样性从上游至下游依次增加。说明NTF在大盈江流域内是非均质分布的。在生态学研究中,多样性是衡量群落稳定性的一个重要指标,常用于反映群落结构的复杂程度和稳定性〔38〕。王佳等〔39〕发现浑河不同河段底泥中微生物多样性指数随水期的变化存在差异,但不仅水期会影响微生物分布,水流、气候等环境条件也都可能会导致微生物分布出现差异〔38〕。本研究选择在一年中微生物群落结构最稳定的时期(3~4月)进行样品的采集,结果仍发现大盈江上、中、下游的物种多样性存在差异,这可能是因为在水流的汇聚作用下,上游的有机物、营养物质等汇入下游,而下游海拔较低、水流速度较慢、底泥更容易沉积,所以下游的NTF检出率和多样性高于上游。
整体上,大盈江陆地土壤中检出的NTF物种数比水体底泥中更多,这与之前的研究结果一致〔22-23〕,说明在流域生态系统内,陆地土壤是微生物栖息、繁殖、演化的主要场所,而流域内微生物的物种多样性可能主要由其临近的陆地土壤贡献。在大盈江的陆地土壤中,仅只有上游和中游间的NTF物种组成存在一定的相似性,中游与下游、上游与下游间的物种组成完全不同。有研究〔40〕表明,土壤样品之间的地理位置相隔越远,微生物群落之间的差异就越明显。在本研究中,由于各样区的陆地土壤间距离较远,且有山脉阻隔,群落间的差异性也就更大;在水体底泥中,整个研究区域内水体底泥间的NTF均存在一定的相似性,并且各样区水体底泥间物种相似性均高于陆地土壤,说明水流的均一性在整条河流的物种分布中具有重要的作用,水流对于NTF在流域内的传播非常关键。尽管水流的联通和均一性使得各样区的水体底泥间NTF物种组成存在一定的相似性,但也具有较大的差异性,这表明流域的上下游间仍然具有较大的环境异质性而使微生物在流域内形成差异性分布。
研究区域内,NTF在上下游存在空间分布格局,水流的汇聚作用是流域内微生物传播的主要驱动力之一。在流域生态系统中,除水流对微生物的传播作用以外,还存在河流自身产生的屏障对微生物形成扩散限制,河流沿岸的陆地土壤间的物种组成还受山脉阻隔的影响。因此,在流域视角下开展微生物空间分布格局研究时,应该兼顾水陆和上、下游等流域特征,将更有助于探索流域内水流的扩散作用和环境异质性在微生物群落演化过程中的相对贡献。
