福建龟山伞花木所在群落乔灌层优势种的种间联结性
2022-12-16罗敏贤林碧华陈绪辉肖丽芳潘标志叶宝鉴郑世群
罗敏贤, 林碧华, 陈绪辉, 肖丽芳, 潘标志, 叶宝鉴, 刘 宝, 郑世群,①
(1. 福建农林大学: a. 林学院, b. 海峡自然保护区研究中心, c. 生态与资源统计福建省高校重点实验室, 福建 福州 350002;2. 福建省林业调查规划院, 福建 福州 350003; 3. 福建工程学院建筑与城乡规划学院, 福建 福州 350118)
伞花木〔Eurycorymbuscavaleriei(Lévl.) Rehd. et Hand.-Mazz.〕为无患子科(Sapindaceae)伞花木属(EurycorymbusHand.-Mazz.)高大落叶乔木,是第三纪孑遗植物和中国特有单种属植物[1],喜光、耐旱,主要分布于云南、贵州、广西、湖南、江西、广东、福建和台湾,湖北神农架亦有分布,且多生长在海拔300~1 400 m的阔叶林中。由于伞花木分布地的森林遭到严重的人为破坏,其野生资源日趋减少,分布范围逐渐缩小,濒临灭绝风险,已被列为国家Ⅱ级重点保护野生植物[2,3]。
种间联结性(interspecific association)指不同物种在空间分布上的相互关联性,是对某个时间范围内物种关系的静态描述,通常受群落内生物因子和非生物因子的影响[4]。种间联结性能够量化群落物种间的作用关系,进而反映物种与生境因子间的耦合关系,有助于正确认识群落的结构与功能,为植被的经营管理、自然植被恢复和生物多样性保护提供理论依据[5-7]。
开展濒危植物群落种间联结性研究能够使人们更准确地了解濒危植物群落的总体联结性和群落稳定性,分析与濒危植物存在竞争或协同共生关系的物种,据此提出针对性强且行之有效的濒危植物保护策略。近年来,国内许多学者对珍稀濒危植物群落的种间联结性展开了相关研究,发现赣北九江市永修县的狭果秤锤树(SinojackiarehderianaHu)与群落内多数乔木种类的种间关系较为松散,群落总体上呈不显著正关联,尚未达到稳定阶段[8];在吉林长白山的对开蕨(AspleniumkomaroviiAkasawa)群落中,对开蕨与多数植物的联结性不显著,各种类趋于相互独立,群落总体上不稳定[9];贵州雷公山的台湾杉(TaiwaniacryptomerioidesHayata)群落的主要乔木种类间以负联结为主,群落总体处于不稳定阶段[10];贵州茂兰国家级自然保护区的四药门花〔Loropetalumsubcordatum(Benth.) Oliv.〕群落内各层次优势种间的联结性较弱,该群落中的四药门花独立性较强,仅与少数优势种存在竞争关系[11]。
经过文献查阅,笔者发现目前关于伞花木的研究主要涉及基因组学和保护遗传学[12-14]、染色体核型[15]、营养成分[16,17]、繁殖培育[18]和群落特征[19]等方面,尚未见关于伞花木群落种间联结性的研究报道,因此,人们尚未了解伞花木与其他物种的种间关系,无法制定有效的就地保护措施。中国东南部地区的伞花木多零星分布于沟谷溪旁的常绿阔叶林中,很少形成群落。2015年,肖书平[20]在福建省明溪县龟山自然保护区内发现了近百株伞花木。本研究以该自然保护区内伞花木所在群落为研究对象,在χ2检验的基础上,结合联结系数、共同出现百分率和点相关系数分析群落内乔木层和灌木层优势种(重要值大于等于2.0%)的种间联结性,尤其是伞花木与其他优势种的种间联结性,以探讨伞花木在该群落中的地位及其与优势种间的关系,为预测该群落的结构变化和演替趋势以及探究伞花木的濒危机制和保护措施提供科学依据。
1 研究区概况和研究方法
1.1 研究区概况
供试伞花木所在群落位于福建省明溪县龟山自然保护区内,地理坐标为东经117°29′、北纬26°38′。该群落分布在龟山的中上坡位置,海拔305~420 m,面积约0.012 km2。该区域属亚热带海洋性季风气候,年平均气温18 ℃,年平均降水量1 800 mm,年平均无霜期261 d,气候温和,雨量充沛,冬少严寒、夏无酷暑,光照充足。区域土壤为石灰岩风化成的钙质土,腐殖质厚约3~4 cm。
供试伞花木所在群落为常绿落叶阔叶混交林,林相较为完整。乔木层高约10.0 m,郁闭度约50%,主要树种有木樨〔Osmanthusfragrans(Thunb.) Lour.〕、青冈〔Cyclobalanopsisglauca(Thunb.) Oerst.〕、光皮梾木(CornuswilsonianaWanger.)、伞花木、光叶细刺枸骨(Ilexhylonomavar.glabraS. Y. Hu)等;灌木层高约1.0 m,盖度约35%,主要种类有伞花木、木樨、紫麻〔Oreocnidefrutescens(Thunb.) Miq.〕、肿节少穗竹〔Oligostachyumoedogonatum(Z. P. Wang et G. H. Ye) Q. F. Zhang et K. F. Huang〕、扶芳藤〔Euonymusfortunei(Turcz.) Hand.-Mazz.〕等;草本层高约0.2 m,盖度约50%,该层次植物种类较少,主要有贯众(CyrtomiumfortuneiJ. Sm.)、山麦冬〔Liriopespicata(Thunb.) Lour.〕、红蓼〔Persicariaorientalis(Linn.) Spach.〕、紫金牛〔Ardisiajaponica(Thunb.) Bl.〕、江南星蕨〔Lepisorusfortunei(T. Moore) C. M. Kuo〕等。
1.2 研究方法
1.2.1 实地调查 参考方精云等[21]的植物群落调查方法,于2021年4月对伞花木所在群落进行实地调查。在伞花木集中分布区域内划分5个面积20 m×20 m的样地,每个样地分成4个面积10 m×10 m的乔木层样方,在每个样地的四角和中央各划分1个面积5 m×5 m的灌木层样方,共20个乔木层样方和25个灌木层样方。详细记录每个乔木层样方内所有株高大于等于5.0 m的乔木个体的种名、株数、高度和胸径等信息,并详细记录每个灌木层样方内所有株高在0.3~ 5.0 m的灌木、木质藤本或乔木幼树的种名、株数、高度和冠幅等信息。分别计算乔木层和灌木层各种类的重要值,计算公式分别为乔木种类重要值=(相对密度+相对频度+相对显著度)/3;灌木层种类重要值=(相对密度+相对频度+相对盖度)/3。其中,相对密度=(某个种类的个体数/全部种类的个体数)×100%;相对频度=(某个种类的频度/全部种类的频度)×100%;相对显著度=(某个种类胸高断面积的总和/全部种类胸高断面积的总和)×100%;相对盖度=(某个种类的盖度/全部种类的盖度)×100%。
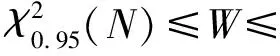
1.2.3 种间联结性分析 根据王伯荪等[23]的方法建立2×2联列表,采用Yates连续矫正公式计算χ2值。χ2<3.841,表示种对联结性不显著(P>0.05);3.841≤χ2<6.635,表示种对联结性显著(0.01 1.2.4 种间联结性检验 虽然χ2检验可定性种间联结性,但无法确定联结程度,为此,本研究通过联结系数(AC)和共同出现百分率(PC)进一步检验χ2结果,确定种对联结程度[22]。AC的值域为[-1,1],-1.0≤AC≤-0.5,表示种对联结性为显著负联结;-0.5 值得注意的是,AC和PC值虽然能够反映种对的联结程度,但对种对联结程度等级的划分缺少统一标准,而且当物种出现次数少,特别是a=0时,AC和PC值均会夸大物种间的联结程度,甚至得出完全相反的结论[7]。点相关系数(PCC)能够降低上述情况对种对联结程度分析结果的影响[22]。-1.0≤PCC≤-0.5,表示种对联结性为显著负联结;-0.5 统计结果(表1)显示:伞花木所在群落乔木层中,重要值大于等于2.0%的优势种有10个,重要值 表1 福建龟山伞花木所在群落乔木层和灌木层优势种的重要值 总和为91.7%。其中,木樨的重要值最高,为21.8%;其次为青冈和光皮梾木,重要值分别为20.5%和17.4%;伞花木的重要值为6.3%,排名第4位。该群落灌木层中,重要值大于等于2.0%的优势种有13个,重要值总和为63.7%。其中,木樨的重要值仍最高(12.8%),但明显低于其在乔木层的重要值;其次为紫麻、肿节少穗竹和青冈,重要值分别为10.5%、6.0% 和5.6%;伞花木的重要值为5.2%,排名第5位,低于其在乔木层的重要值。 乔木层10个优势种的χ2检验结果(图1)显示:在这些优势种组成的45个种对中,正联结种对有20个,负联结种对有25个,分别占种对总数的44.4%和55.6%,正、负联结种对数的比值为0.8∶1.0。值得注意的是,所有种对的χ2值均小于3.841,说明乔木层优势种的种间联结性均不显著。 灌木层13个优势种的χ2检验结果(图2)显示:在这些优势种组成的78个种对中,正联结种对有43个,负联结种对有35个,分别占种对总数的55.1%和44.9%,正、负联结种对数的比值为1.2∶1.0。这些种对中,仅肿节少穗竹-青冈种对呈极显著正联结(χ2≥6.635),扶芳藤-光叶细刺枸骨种对呈显著正联结(3.841≤χ2<6.635),其余76个种对的联结性均不显著。 2.4.1 联结系数 从乔木层优势种间的联结系数(表2)看,正联结种对有20对,负联结种对有24个,无联结性种对有1个,分别占种对总数的44.4%、53.3%和2.2%,正、负联结种对数的比值为0.8∶1.0,与χ2检验结果基本相同。其中,联结性不显著的种对有20个,占种对总数的44.4%;显著正联结种对有10个,占种对总数的22.2%,其中8个种对由于b值为0,导致联结系数为1.000,致使联结系数在0.5及以上的种对偏多,这一研究结果并不符合实际情况;显著负联结种对有14个,占种对总数的31.1%,其中12个种对由于a值为0,导致联结系数为-1.000,致使联结系数在-0.5及以下的种对偏多,这一研究结果也不符合实际情况。值得注意的是,伞花木仅与木樨和山牡荆〔Vitexquinata(Lour.) Will.〕呈显著负联结,与其他优势种的联结性均不显著。 : 不显著负联结Non-significant negative association; : 不显著正联结Non-significant positive association. : 不显著负联结Non-significant negative association; : 不显著正联结Non-significant positive association; : 显著正联结Significant positive association; : 极显著正联结Highly significant positive association. 表2 福建龟山伞花木所在群落乔木层优势种的联结系数1) 从灌木层优势种间的联结系数(表3)看,正联结种对有41个,负联结种对有36个,无联结性种对有1个,分别占种对总数的52.6%、46.2%和1.3%,正、负联结种对数的比值为1.1∶1.0,与χ2检验结果基本一致。其中,联结性不显著的种对有58个,占种对总数的74.4%;显著正联结的种对有10个,占种对总数的12.8%,其中紫麻与香叶树(LinderacommunisHemsl.)和络石〔Trachelospermumjasminoides(Lindl.) Lem.〕的联结系数均为1.000;显著负联结的种对有9个, 占种对总数的11.5%,其中香叶树与扶芳藤、臭常山(OrixajaponicaThunb.)和络石以及小蜡(LigustrumsinenseLour.)与臭常山的联结系数均为-1.000。值得注意的是,伞花木与紫麻和络石呈显著正联结,与小蜡呈显著负联结,与其他优势种的联结性均不显著。 2.4.2 共同出现百分率 从乔木层优势种间的共同出现百分率(表4)看,木樨与青冈、光皮梾木和光叶细刺枸骨的联结性显著,这些种对的共同出现百分率分别为0.833、0.500和0.500;其余42个种对的联结性不显著,占种对总数的93.3%。该层次中,伞花木与其他优势种的联结性均不显著。 表3 福建龟山伞花木所在群落灌木层优势种的联结系数1) 表4 福建龟山伞花木所在群落乔木层优势种的共同出现百分率1) 从灌木层优势种间的共同出现百分率(表5)看,肿节少穗竹与木樨、紫麻、青冈和臭常山的联结性显著,这些种对的共同出现百分率分别为0.500、0.500、0.643和0.533;扶芳藤与光叶细刺枸骨以及伞花木与紫麻的联结性也显著,2个种对的共同出现百分率分别为0.538和0.529;其余72个种对的联结性均不显著,占种对总数的92.3%。该层次中,伞花木与其他优势种(紫麻除外)的联结性基本不显著。 2.4.3 点相关系数 从乔木层优势种间的点相关系数(表6)看,正联结种对有19个,负联结种对有25个,无联结性种对有1个,分别占种对总数的42.2%、55.6%和2.2%,正、负联结种对数的比值为1.0∶1.3。其中,联结性不显著的种对有42个,占种对总数的93.3%;显著正联结种对仅有1个,即木樨-青冈种对,二者的点相关系数为0.577;显著负联结种对也仅有1个,即鸡仔木〔Sinoadinaracemosa(Sieb. et Zucc.) Ridsd.〕-木樨种对,二者的点相关系数为-0.577。 表5 福建龟山伞花木所在群落灌木层优势种的共同出现百分率1) 表6 福建龟山伞花木所在群落乔木层优势种的点相关系数1) 从灌木层优势种间的点相关系数(表7)看,正联结种对有41个,负联结种对有36个,无连接性种对有1个,分别占种对总数的52.6%、46.2%和1.3%,正、负联结种对数的比值为1.1∶1.0。其中,联结性不显著的种对有75个,占种对总数的96.2%;显著正联结种对有2个,分别为青冈-肿节少穗竹种对和光叶细刺枸骨-扶芳藤种对,2个种对的点相关系数分别为0.600和0.510。 表7 福建龟山伞花木所在群落灌木层优势种的点相关系数1) 植物群落的总体种间联结性与群落的稳定性密切相关。研究表明:随着植物群落的正向演替,不同层次和不同生活型的植物种类通过相互竞争和合作逐渐趋于和谐共生的状态,种间关系逐渐趋于正联结,最终发展成稳定的顶极群落[24,25]。总体种间联结性结果与χ2检验、联结系数和点相关系数结果均显示:福建龟山伞花木所在群落乔木层优势种表现为不显著负联结,灌木层优势种表现为不显著正联结,说明福建龟山伞花木所在群落的乔木层和灌木层的优势种尚未形成稳定的种间关系。并且,乔木层作为森林群落的优势层,是群落发展方向的重要影响因子,据此可推断该伞花木所在群落处于相对不稳定的演替阶段[26,27]。 χ2检验侧重于检验种间联结性是否显著,联结系数和共同出现百分率可反映χ2检验判断为种间联结性不显著的种对的联结程度。一般情况下,在χ2检验结果显示联结性不显著的种对中,往往能发现个别种对的联结系数和共同出现百分率显示其种间联结性显著。当a=0时,联结系数和共同出现百分率往往会得出完全相反的结论,使得相关种类的种间联结性被夸大。例如:在伞花木所在群落乔木层的10个优势种中,联结系数为-1.000的种对有12个,其中9个种对的共同出现百分率为0.000;而在灌木层的13个优势种中,联结系数为-1.000的种对有4个,且其共同出现百分率均为0.000。点相关系数可降低a或b值为0时对种对联结性程度分析结果的影响,通过点相关系数分析,可明显降低联结系数和共同出现百分率分析中种对联结性的显著程度。因此,以χ2检验为基础、以联结系数、共同出现百分率和点相关系数为辅助参数进行综合分析,能够更准确地判定福建龟山伞花木所在群落乔木层和灌木层优势种的种间联结性[28,29]。 相关研究结果表明:对环境需求相似或互利共生的物种往往表现为显著正联结,而对环境需求有差异或在资源竞争中互相排斥的物种往往表现为显著负联结[30-32]。本研究结果也验证了这一规律。例如:在伞花木所在群落乔木层中,共同出现百分率结果显示木樨与青冈、光皮梾木和光叶细刺枸骨呈显著正联结,这是因为这些植物的更新能力强、喜光、对土壤和水分要求不严,适应性广;而点相关系数结果显示木樨与鸡仔木呈显著负联结,这与木樨喜光,而鸡仔木在本地喜生于水边或林内潮湿处有关;联结系数结果显示伞花木与木樨呈显著负联结,原因是木樨为常绿树种,伞花木为落叶树种,二者生长需光性不同,并且木樨枝叶浓密,林下光照度极低,可能会造成伞花木的种子萌发和幼苗生长没有合适的环境条件。在灌木层中,综合χ2检验、联结系数和点相关系数结果,肿节少穗竹分别与青冈、木樨、紫麻、臭常山呈显著正联结,由于肿节少穗竹具有入侵性,易迅速占领群落,完全或部分遮挡其他种类的光照,对林下灌木和乔木的更新小植株易形成强烈的竞争,因此,在福建龟山伞花木所在群落中需要注意疏伐肿节少穗竹,以维护该群落的植物多样性[33];共同出现百分率和联结系数结果显示伞花木与紫麻和络石均呈显著正联结,伞花木在灌木层为幼苗或幼树阶段,平均高度0.69 m,紫麻常生于林下,为小型灌木,二者可能对生境有基本相同的偏好;络石常生于山野岩石上和攀伏在树体上,适应性和抗逆性都很强。联结系数结果显示伞花木与小蜡呈显著负联结,相比之下,伞花木在幼苗和幼树阶段需要充足的光照,要求乔木层郁闭度小,而小蜡对光的适应范围广,乔木层郁闭度可以有较大波动。伞花木与其余大部分物种的相关性均不太明显,种间关系较为松散,独立性较强。 值得注意的是,笔者在调查过程中共发现伞花木幼苗(株高低于0.3 m)34株,幼树(株高低于5.0 m)73株,成年植株(株高在5.0 m及以上)8株,表明在调查群落中,虽然伞花木幼苗和幼树数量较多,但成年植株偏少,可见福建龟山伞花木所在群落中伞花木幼苗和幼树的生存和生长面临巨大挑战,生境条件不利于其生长发育。笔者还发现,该群落中的伞花木幼苗大多生长在裸露的路边,易受到人为活动、动物采食和自然灾害等干扰,这也在一定程度上影响了其幼苗的存活率。因此,为了促进伞花木幼苗和幼树的生长和发育,应积极实施人工辅助措施,确保其种群正常更新。建议在伞花木的生长地划定特定区域、设置指示牌和宣传栏,降低人为干扰。另外,建议适当疏伐伞花木所在群落中具有入侵性的肿节少穗竹以及与伞花木存在显著负联结的种类(如木樨),定期适度清理病腐木,为伞花木种群发展创造合适的生存条件。 影响树种种间关系的因子除生态适应性外,还包括个体发育阶段和群落演替阶段[34]。本研究仅采用种间联结方法初步分析了现阶段福建龟山伞花木所在群落乔木层和灌木层优势种的种间关系,关于群落内不同种间的生态关系还需进一步深入研究。例如:运用点格局分析法,分析群落内伞花木在不同空间尺度和不同年龄结构的分布格局及种内关联性,进一步探究伞花木所在群落的种内和种间关系;建立固定样地,通过监测生态因子计算生态位,并分析生态位与种间联结的关系,从而揭示伞花木所在群落种间联结的内在机制;还可以从植物生理学和分子遗传学角度研究伞花木所在群落的种间联结,探讨种间联结产生的机制[22,35]。 根据上述研究结果,福建龟山伞花木所在群落乔木层和灌木层的优势种并未形成紧密的关系,种间联结性较低,各优势种均相对独立,群落尚处于相对不稳定的演替前期,宜对该伞花木所在群落采取就地保护措施,积极实施人工辅助措施,确保伞花木种群正常更新。2 结果和分析
2.1 伞花木所在群落优势种分析

2.2 伞花木所在群落优势种的总体种间联结性分析


2.3 伞花木所在群落优势种的种间联结性分析
2.4 伞花木所在群落优势种的种间联结性检验








3 讨论和结论
