亚热带不同纬度森林群落物种空间分布格局
2022-12-16温智峰魏识广叶万辉练琚愉
温智峰,魏识广,李 林,*,叶万辉,练琚愉
1 桂林电子科技大学,桂林 541004
2 广西师范大学珍稀濒危动植物生态与环境保护教育部重点实验室,桂林 541006
3 华南植物园中国科学院退化生态系统植被恢复与管理重点实验室,广州 510650
4 中国科学院核心植物园植物生态中心,广州 510650
物种个体的空间分布是生物多样性监测和研究的核心,个体在群落中聚集或分散的程度,体现了物种对生境的适应情况[1]。植物个体的空间分布格局是种群自身特性、种间关系和生境因素综合作用的结果[2],是研究种群特征、种间关系、种群与生境因子相互影响的重要手段[3],因此群落中物种的空间分布格局及其形成机制一直是植物生态学研究的热点问题[4]。揭示植物群落中物种的空间分布格局及其成因,可以为群落的更新、演替及多样性维持机制提供重要线索[5]。
森林群落中物种个体呈现的空间分布格局是由多种生态过程共同作用的结果[6],也是森林群落中多种多样性维持机制综合作用的最直接体现[4]。因此,通过了解物种个体在群落中的分布可以获取种群的空间分布特征,能够进一步揭示格局产生的生态学过程,例如:研究生境异质性、扩散限制、密度制约、随机作用等过程对群落中物种空间分布格局形成的作用[7],可以解释不同群落物种共存机制和多样性的维持状况[8]。
物种空间分布格局的研究主要集中在群落中的优势种。优势种既作为群落“骨架”,又影响群落类型[9],对群落的结构具有决定作用[10]。分析群落中优势种群的空间分布格局,可以阐明群落生境的形成和维持机制,群落的稳定性以及更新和演替的规律[11—12]。此外,由于优势种在群落中具有数量优势,满足空间点格局统计模型的样本精度要求,所以通常被选作代表群落多样性格局分布状况的研究对象。相反,群落中低多度物种的空间格局分析鲜有报道,现状为:一方面由于低多度物种个体数量少,传统意义上以物种为单位统计其空间分布格局时受限于模型统计的样本量不足,为确保统计精度而舍弃低多度物种;另一方面,研究发现群落中物种的丰富度格局主要由大量的低多度物种决定,在物种多样性丰富的群落中低多度物种所占的比例不容忽视[13]。低多度物种是群落生物多样性中非常重要的组分,研究其空间分布格局及成因,可以为生物多样性保护提供可靠依据,特别是为濒危物种保护提供理论基础,所以对群落的空间分布格局相关研究需要把优势物种和低多度物种同时纳入研究范围。
目前,关于高丰富度群落中物种空间格局分布的研究大多都来自于热带地区[14],美国Smithsonian研究院CTFS(The Center for Tropical Forest Science)已经建成热带森林生物多样性监测网络,进行了森林生态系统群落分布、结构组成和物种多样性等方面的研究[15]。为了揭示其他气候带生境中的物种分布和物种共存机制,我国的生态学家借鉴CTFS大样地建设方法,建成了中国森林生物多样性监测网络,取得了一系列研究成果,如:在温带,对长白山次生杨桦林种群的空间点格局分析结果证明了密度制约是调节温带树木空间分布格局的主要机制[16];北京东灵山落叶阔叶林群落中物种的空间格局分布研究发现了树木径级大小在空间格局分布上互补且不同径级个体占据不同生态位[17]。在亚热带,对古田山大样地群落的研究证实了种子性状是影响物种空间分布格局的重要因素[18];对鼎湖山常绿阔叶林群落的分析证明了聚集分布是其主要空间分布格局类型[14],大径级物种个体的空间分布趋向于随机分布[15]。在热带,尖峰岭热带山地雨林物种多样性空间分布格局具有明显的尺度效应[19],小径级植株比大径级植株对生境变化更加敏感,物种空间格局受到多种因素影响[20]。以上研究大多是分散在各个纬度带上的独立讨论,所得结果虽然较好地解释了对应地带群落的研究问题,但是由于缺乏研究尺度上的连续性,结论有待基于不同纬度群落的连续研究进行验证和拓展。
1 研究区域
1.1 研究区域概况
本研究选取中国亚热带季风气候区域4个纬度带上6个1 hm2典型森林群落,物种数据来源于中国科学院华南植物园生态研究中心的数据共享,调查时间跨度为2010年到2012年[21]。各群落基本信息见表1。
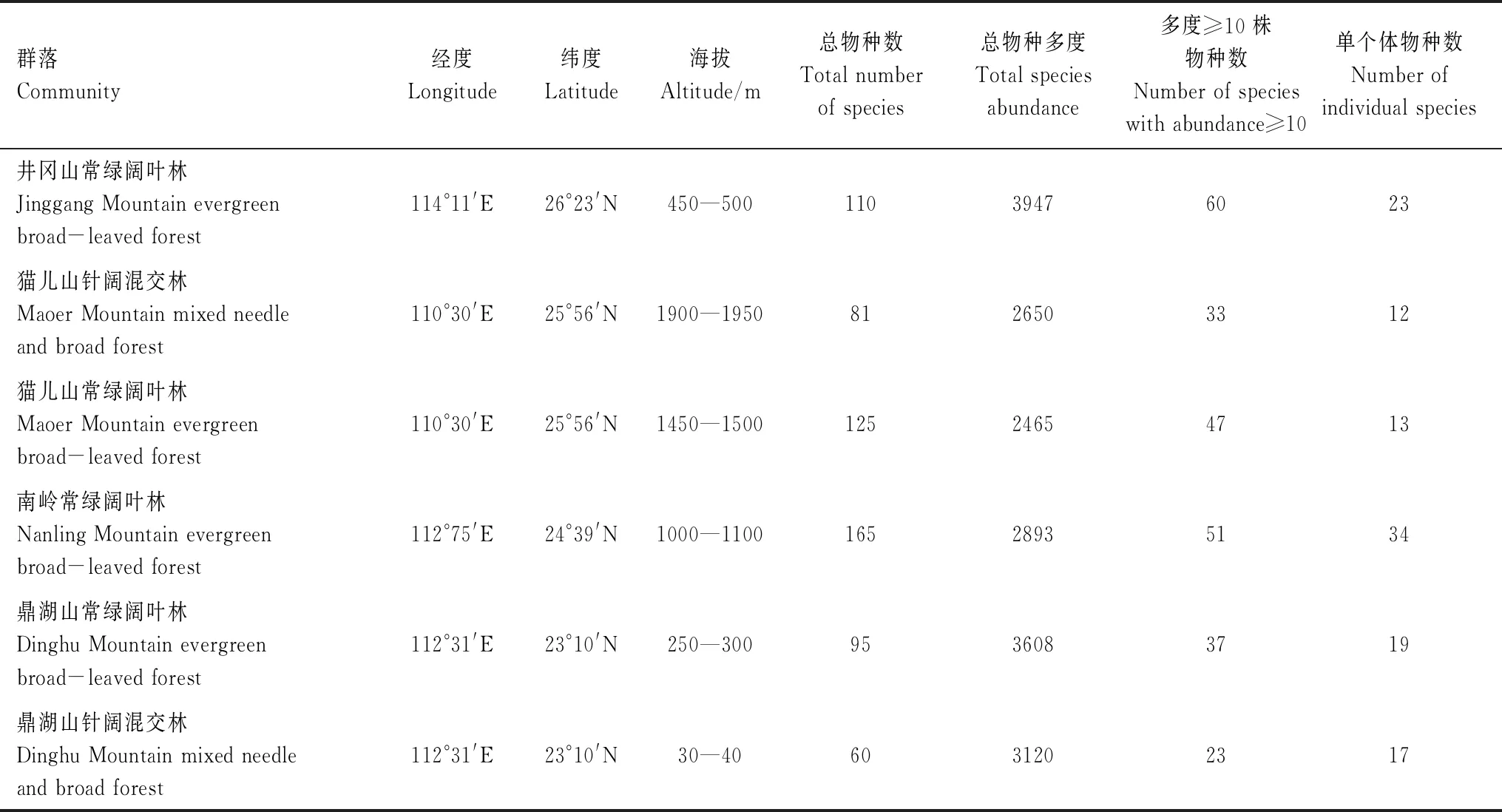
表1 各群落基本信息
1.2 样地设置与调查
各样地的建设和野外调查均按照CTFS国际标准进行。6个面积1 hm2的样地均采用相邻格子法设置25个20 m×20 m的样方,每个样方再细分为16个5 m×5 m的小样方。以5 m×5 m小样方为基本单元,对样方内植株进行每木标尺测量,标记每个胸径(DBH)≥1 cm的个体及其分枝,记录物种的名称、胸径、坐标、生长状况等信息,并建立专项数据库。
2 研究方法
2.1 空间点格局分析方法
采用环形取样方式,研究种群空间分布格局采用Ripley′sK(t)方程的变换L(t)方程进行分析[22],方程的公式表达如下:
(1)
(2)
式中,A为样地面积;n为样地内物种个数;t为距离尺度;Uij为植物两点i和j之间的距离;当Uij≤t时,Ir(Uij)=1,反之为0;Wij为以i为圆心Uij为半径的圆在A中弧长与整个圆周长的比值,表示一个植物被观测到的概率[23]。
Ripley′sK(t)方程可以用来解释以t为半径圆中的物种分布模式,而在实际中为了稳定方差,简化解释,经常使用L(t)方程进行分析[24]。L(t)方程以植物个体在样地中的实际坐标为基本数据,在分析过程中能最大限度地利用个体在二维空间上的坐标,其最大优势在于多尺度的空间格局分析。在均质泊松分布假设下,L(t)的期望值等于0,如果L(t)>0,则物种呈聚集分布,如果L(t)<0,物种则呈均匀分布,若L(t)=0,则为随机分布[23]。本文选用均质泊松分布零模型(CSR),通过Monte-Carlo随机排列999次得到99%的置信区间计算分布的上下包迹线,若计算出的L(t)值曲线在上包迹线以上时,为显著聚集分布格局,若在上下包迹线之间时,为显著随机分布格局,若在下包迹线以下时,为显著均匀分布格局。由于L(t)方程采用环形取样法,样地最大边长为100 m,为消除边缘效应,取样的最大尺度设置为50 m。
2.2 低多度与单个体物种的空间点格局分析
为了满足L(t)方程的统计精度,将各群落中多度≥10的植物以物种为单位统计,进行空间点格局统计分析;多度<10的物种则划分为单个体种组(多度=1)和低多度种组(多度=2—9),满足统计精度后分别对其进行空间点格局统计分析。
空间点格局分析使用R 4.0.0 软件及其下的“splancs”包来编程计算(R Core Team 2020. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL https://www.R-project.org/.)。
另外,为了防止鼻子呛水,女选手们都用一只肉色的夹子夹住两个鼻孔,因此,从远处望去,她们表演时鼻子特别尖长。表演一结束,她们赶紧将夹子取下,这不光是为了好看,恐怕夹得也难受了吧。
2.3 多度和胸径与空间聚集程度的关系分析
因为相邻尺度的L(t)值之间高度相关,所以本研究选取L10(10 m取样尺度上的统计值)作为表征各群落物种空间聚集程度的指标[1]。物种多度与空间聚集程度指标L10的关系采用双变量Pearson相关性分析方法。对各群落的物种多度进行自然对数转换,并用多度的对数值与空间聚集程度指标L10进行曲线拟合。物种胸径(DBH)与空间聚集程度的关系采用最大DBH、最小DBH和平均DBH与空间聚集程度指标L10进行多元线性回归分析。
使用SPSS 26进行相关性分析和多元线性回归分析,使用Origin 2018软件绘图。
3 结果
3.1 各群落多度≥10株物种空间分布格局及其变化规律
各群落中物种显著聚集分布比例具有相同的尺度变化趋势,均为随尺度先增大后降低(图1)。其中,猫儿山和鼎湖山2个针阔混交林群落在20—25 m尺度时显著聚集分布比例达到峰值,4个常绿阔叶林群落均在10 m尺度时达到峰值。显著聚集分布比例变化幅度随纬度由高到低呈现先增后减的趋势,纬度从高到低各群落的变化幅度为:井冈山常绿阔叶林33.33%;猫儿山针阔混交林和常绿阔叶林分别为39.39%、53.19%;南岭常绿阔叶林43.14%;鼎湖山常绿阔叶林和针阔混交林分别为29.73%、27.27%。

图1 各群落中物种显著聚集分布比例随尺度变化规律
在不同取样尺度上各群落物种空间分布格局类型比例见表2。不同纬度的群落均在小尺度上以显著聚集分布为主,大尺度上以随机分布为主,显著均匀分布仅出现在较大取样尺度时的部分群落,占比极少。具有明显尺度效应。4个常绿阔叶林群落以30 m作为主要格局类型变化的尺度节点,取样尺度<30 m时以显著聚集分布为主,尺度≥30 m时以随机分布为主;2个针阔混交林群落,主要格局类型变化的尺度节点明显更大。

表2 各群落中物种空间分布格局类型在不同尺度时所占的比例/%
3.2 各群落单个体种组和低多度种组空间分布格局及其变化规律
各群落中单个体种组空间点格局分析结果显示(图2):不同纬度群落中单个体种组均以随机分布为主,格局随尺度变化规律不明显。仅井冈山常绿阔叶林群落在取样尺度接近50 m时出现显著均匀分布,以及南岭常绿阔叶林群落在20—35 m尺度表现为显著聚集分布,其余群落所有尺度均呈现随机分布。单个体物种的空间分布点图验证了单个体种组在大多数尺度上随机分布的空间格局(图3)。
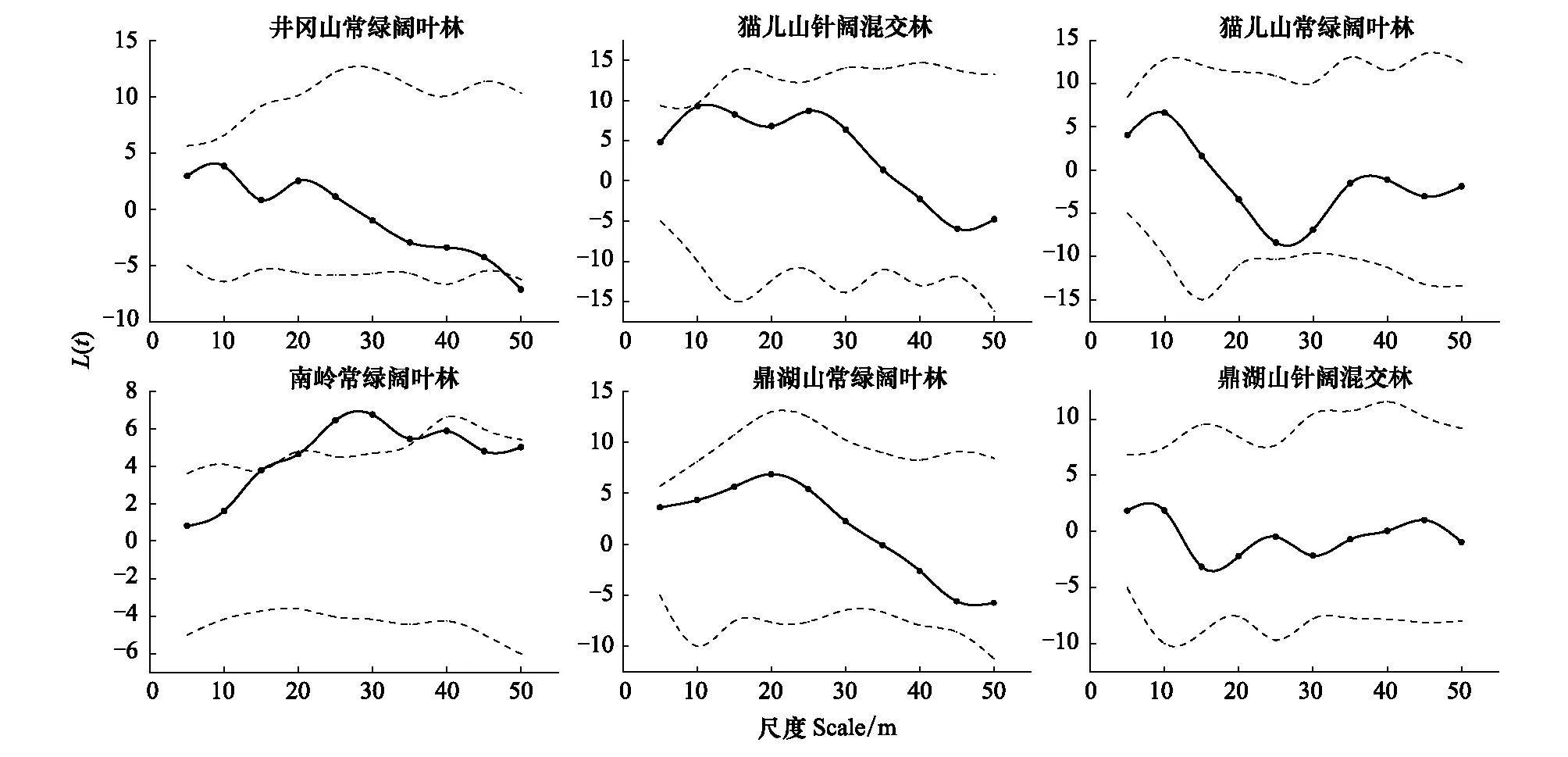
图2 各群落中单个体种组的空间点格局分析结果
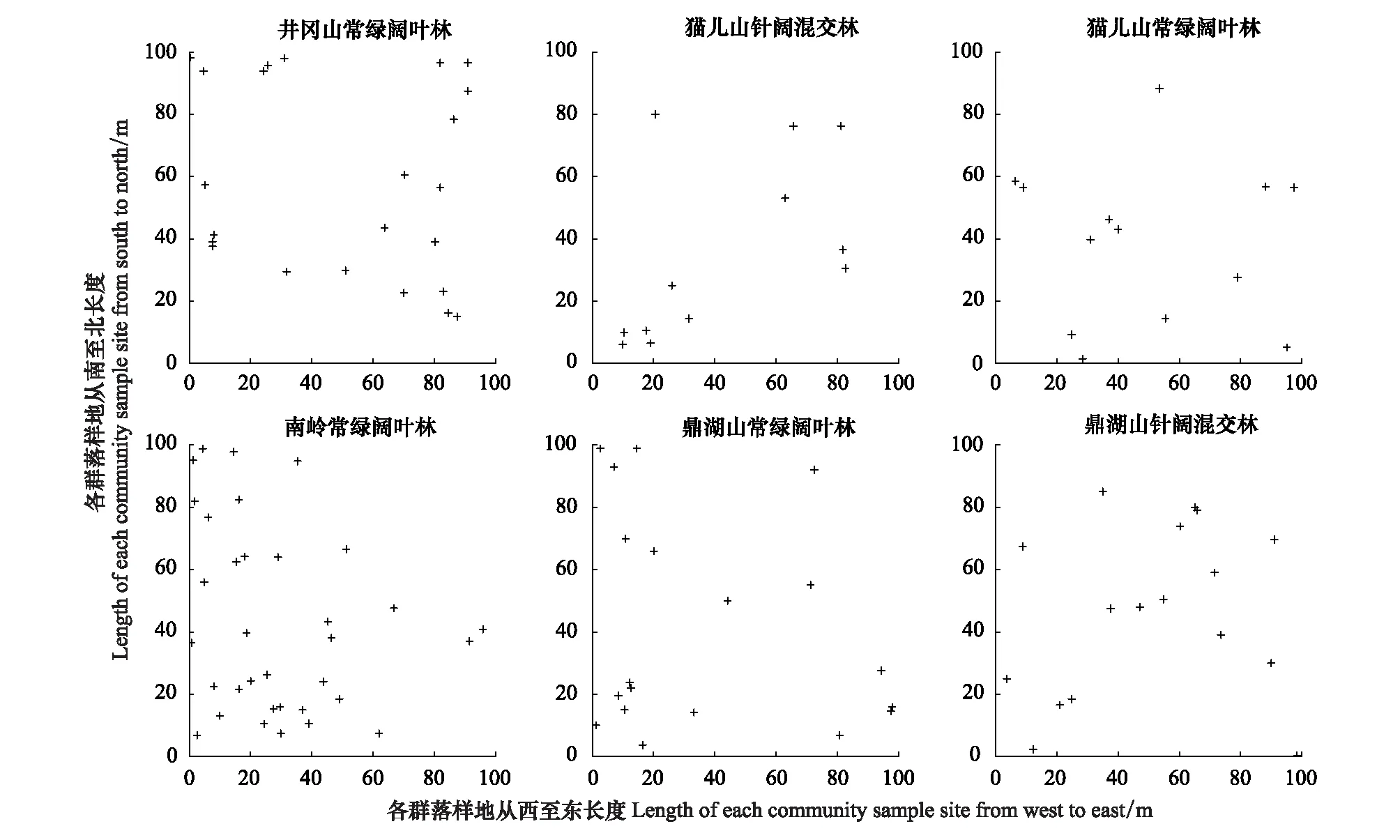
图3 各群落中单个体物种的空间分布点图
不同纬度群落低多度种组空间点格局呈现小尺度上显著聚集分布,大尺度上随机分布甚至出现显著均匀分布的特点,具有明显的尺度效应,但不同纬度群落间尺度效应存在差异(图4)。低多度种组在井冈山常绿阔叶林所有取样尺度均为随机分布,其余群落主要格局类型变化尺度各不相同:猫儿山、南岭和鼎湖山3个常绿阔叶林群落以35 m为主要格局类型变化的尺度节点;猫儿山和鼎湖山2个针阔混交林群落主要格局类型变化的尺度节点更大,为40—45m。规律性与各群落非低多度种组的物种相同。
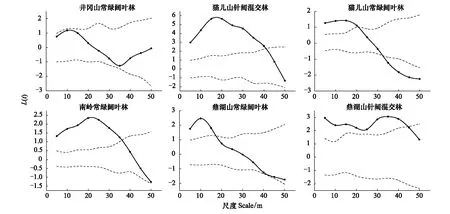
图4 各群落中低多度种组的空间点格局分析结果
3.3 物种多度与空间聚集程度的关系及其变化规律
物种多度与空间聚集程度指标L10的双变量相关性分析表明(表3):物种多度与L10均表现出负相关关系,纬度从高到低对应群落的相关性分析P值表现出数值递增的规律,P值表征的负相关关系从极显著变化到显著,最后到不显著。具体表现为:井冈山常绿阔叶林、猫儿山针阔混交林和常绿阔叶林群落的物种多度与L10极显著负相关(井冈山常绿阔叶林P=0.001,猫儿山针阔混交林P=0.006,猫儿山常绿阔叶林P=0.010);南岭常绿阔叶林群落的物种多度与L10显著负相关(P=0.048);鼎湖山常绿阔叶林和针阔混交林群落的物种多度与L10的负相关关系不显著。
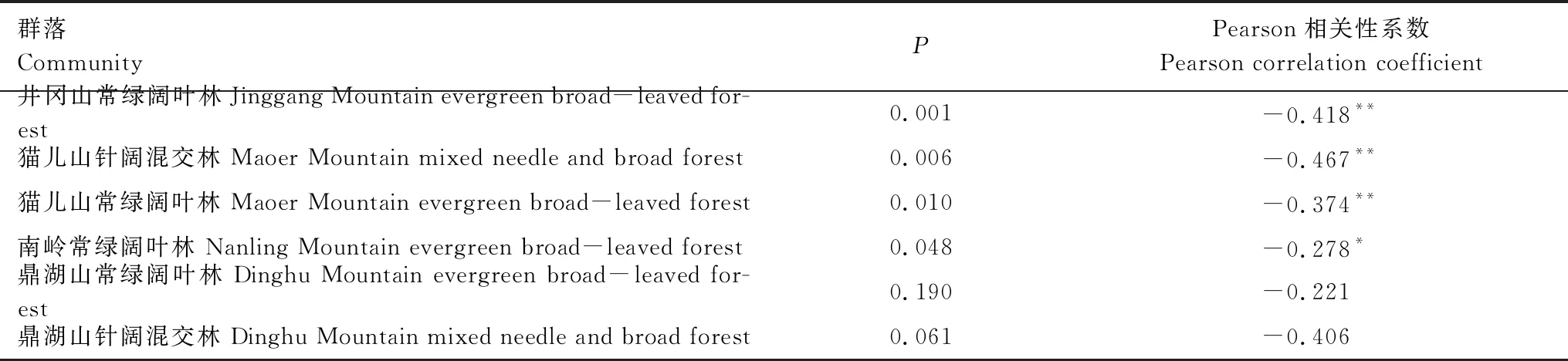
表3 物种多度与空间聚集程度的相关性分析
将多度的对数值与L10进行多次拟合后发现,幂函数的拟合优度(R2)明显高于线性拟合和其他曲线拟合方式。拟合后的幂函数曲线验证了物种多度与空间聚集程度的负相关关系(图5),各群落中物种的空间聚集程度随着多度的增大而降低,物种空间聚集程度的递减速率随着多度的增大逐渐降低。
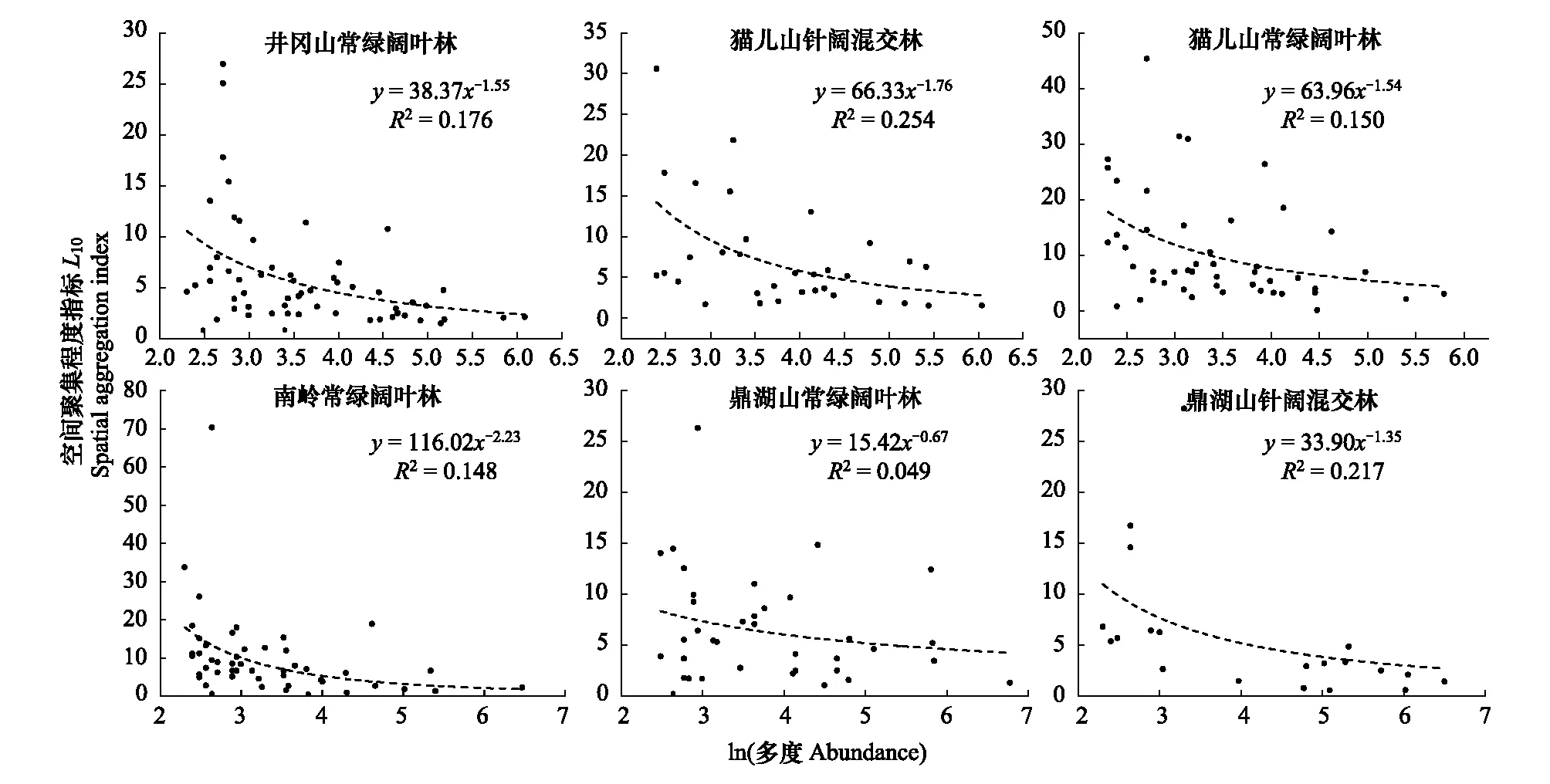
图5 各群落物种多度与空间聚集程度的关系
3.4 物种胸径与空间聚集程度的关系及其变化规律
各群落中物种平均DBH、最大DBH、最小DBH与空间聚集程度指标L10相关性分析表明(表4):各群落物种平均DBH与空间聚集程度L10的相关性系数均为负;最大DBH与L10的相关性系数主要为负;最小DBH与L10的相关性系数主要为正。显著性检验验证了猫儿山常绿阔叶林群落的物种平均DBH与L10极显著负相关;井冈山和猫儿山2个常绿阔叶林群落的物种最大DBH与L10极显著负相关;其他群落的相关关系均未达到显著性检验水平,不具统计学意义。

表4 物种胸径与空间聚集程度的相关性分析
同时,对物种的平均DBH、最大DBH、最小DBH与L10进行多元线性回归分析,发现随着纬度的由高到低,各群落物种胸径对空间聚集程度的解释率总体趋势降低(图6),由最高纬度的19.7%(井冈山常绿阔叶林)降低到了最低纬度的3.9%(鼎湖山针阔混交林),说明物种胸径对空间聚集程度的影响在高纬度群落更加明显。

图6 各群落的物种胸径对空间聚集程度的解释率
4 分析与讨论
4.1 不同纬度森林群落物种空间分布格局及其规律
本研究中亚热带不同纬度森林群落物种的分布格局规律验证了物种空间分布格局具有尺度依赖性[25],并且不同群落中物种空间分布格局的成因存在差异[26]。随取样尺度增大,显著聚集分布比例先增大后减小,小尺度上以显著聚集分布为主,大尺度上以随机分布为主,符合物种在森林群落中空间分布的一般规律[4]。物种显著聚集分布比例的变化幅度能够在一定程度上反映森林群落空间分布格局的稳定性[27],不同纬度各群落物种显著聚集分布比例随取样尺度的变化趋势虽然相同,但变化幅度存在差异:纬度最高的井冈山常绿阔叶林群落变化幅度处于中间水平,处于中间纬度的猫儿山常绿阔叶林、针阔混交林以及南岭常绿阔叶林群落变化幅度较大,纬度最低的鼎湖山常绿阔叶林和针阔混交林群落变化幅度较小,推测是由于纬度最低群落具有较高的生物多样性,因此具有较高稳定性和较强抵御能力。在同一纬度,不同海拔的森林群落林型会存在差异,而本研究发现猫儿山和鼎湖山的常绿阔叶林和针阔混交林群落物种空间分布格局存在差异性,常绿阔叶林群落显著聚集分布比例随尺度变化幅度明显高于针阔混交林群落;针阔混交林群落主要格局类型变化的尺度节点明显大于常绿阔叶林群落(图1),推测林型的不同可以通过影响林分密度以及群落物种组成,进而影响物种空间分布格局,使其有所差异[28],因此认为林型也是群落影响物种空间分布格局的因素之一。
目前普遍认为生境异质性、扩散限制、密度制约以及随机作用等是影响群落物种空间分布格局的重要因素和群落多样性维持的驱动机制[29]。森林群落的生物多样性分布具有明显的纬度梯度规律[30],纬度不同时,群落中的物种类型、气温、太阳辐射强度、地形因子以及其他环境因素均会随之发生改变,进而引起物种的空间分布格局随纬度梯度的变化,这可以解释不同纬度的群落虽然空间分布格局变化的趋势相同,但变化幅度存在差异的原因,说明生境异质性作用在亚热带森林群落中普遍存在。
聚集分布是群落优势种主要空间分布格局类型[1],在亚热带区域6个群落中多度≥10株的物种空间分布格局也在大部分取样尺度呈现聚集分布(表1)。而为了更全面的了解群落中所有物种的空间分布格局,还需要关注低多度物种在群落中的分布。低多度物种由于数量稀少,很少像优势种那样存在较强的种内竞争排斥作用[31],小尺度上形成聚集分布格局有助于发挥低多度种群的群体效应,增大种间竞争力来维持种群稳定[32],但是由于低多度物种个体数量有限,所以往往在大尺度上会表现为随机分布或均匀分布。这与本研究中低多度物种空间分布格局规律性一致:不同纬度亚热带森林群落中的低多度种组空间分布格局在小尺度上也是显著聚集分布,随尺度增大出现随机分布,各群落均存在物种分布的尺度效应,但在不同纬度上存在差异(图4)。 纬度最高的井冈山常绿阔叶林群落有别于其他群落,其低多度种组在所有尺度上均表现出随机分布格局,未检测到聚集分布。本研究以单个体种组为单位统计分析后发现各群落中单个体物种之间的分布是随机的,没有检测出预期的聚集分布趋势,所体现出的随机分布还需要进一步结合亲缘种组或与单个体物种邻近物种的关系等进行深入研究。
4.2 物种多度与空间聚集程度的关系
热带森林群落中的研究表明,相同取样面积时低多度物种空间聚集程度明显高于高多度物种[1]。本研究各群落物种多度与空间聚集程度指标L10的分析结果验证了以上规律在亚热带森林群落中同样适用。由于优势种在群落中数量较多,种间具有较强竞争排斥作用[31],高密度聚集情况下种内竞争凸显,受密度制约作用,反映为空间聚集程度降低;而相对于广布的优势种,低多度种对生境依赖性更强,反映为空间聚集程度提高。但不同纬度各群落物种多度对空间聚集程度的影响水平并不相同,各群落物种多度与空间聚集程度指标L10的负相关关系随纬度从高到低逐渐减弱,说明物种多度对空间聚集程度的影响逐渐降低。推测是由于纬度降低,生境多样性和复杂性随着群落生产力增大而提高[33],这使影响物种空间聚集程度的因素增多,从而使物种多度对空间聚集程度影响水平显著性降低。
4.3 物种胸径与空间聚集程度的关系
群落中物种个体生长早期的空间分布格局一般表现为明显的聚集性分布,随着个体径级的增大,种内竞争加剧[29],导致空间聚集程度降低,表明胸径与空间聚集程度存在负相关关系。但本研究中不同纬度森林群落物种胸径和空间聚集程度的规律性并不完全与之相同:仅纬度较高的井冈山常绿阔叶林和猫儿山常绿阔叶林、针阔混交林群落物种胸径与空间聚集程度指标L10间显著负相关,纬度较低群落物种胸径与空间聚集程度指标L10相关性均不显著。这说明在纬度较高的群落中,物种胸径是影响空间聚集程度的重要因素之一,而随纬度降低,生境复杂性增强,胸径对空间聚集程度的影响逐渐减弱。此外,多元线性回归发现,随着纬度由高到低,物种胸径对空间聚集程度的解释率逐渐降低,由19.7%降低到3.9%,这一规律同样反映了胸径对空间聚集程度的影响随纬度降低而减少。群落中物种胸径和空间聚集程度的关系亦受到监测尺度、生境类型和环境因子的影响,需综合考虑其他因素得出更可靠的规律。
4.4 亚热带森林群落的物种空间分布格局形成原因
森林群落物种空间分布格局的形成受到多种机制影响[34],而生境异质性被认为是大尺度上主要的影响因素[35],对群落中物种空间分布和多样性格局形成及维持有重要作用[36]。本研究中不同纬度群落物种空间分布格局所表现出来的差异性有力的支持了这一观点,例如不同纬度各群落显著聚集分布比例随取样尺度的变化幅度不同以及各群落低多度种组的空间分布格局存在差异。除生境因素以外,物种的种子性状和扩散模式不同也会导致空间分布格局产生差异[18],所以扩散限制同样被认为是群落格局有所差异的原因。有研究发现群落中物种的空间分布格局可能由生境异质性与扩散限制的联合作用决定[37],但生境异质性的贡献高于扩散限制[38]。本研究各群落的物种多度和胸径与物种空间聚集程度的负相关关系也从不同侧面反映出密度制约作用也是亚热带区域物种分布格局形成的影响因素,而大尺度上的随机分布以及单个体物种的随机分布也暗示着随机作用对群落物种分布格局有所影响。综上所述,推测影响亚热带森林群落物种空间分布格局形成的主要原因是由于因纬度变化引起的生境异质性,而林型、扩散限制、密度制约作用和随机作用为次要原因。
5 结论
本研究对我国6个不同纬度亚热带森林群落物种空间分布格局及其成因进行研究,所得结论如下:(1)亚热带不同纬度群落中多度≥10株物种的空间分布格局表现出相似的尺度依赖性规律:显著聚集分布比例随尺度增大而降低,不同纬度群落的变化幅度存在差异,纬度较低的群落稳定性更强。(2)各群落中低多度种组空间分布格局表现出小尺度显著聚集分布,大尺度随机分布的尺度效应,但尺度效应在不同纬度群落间存在差异;各群落单个体种组空间分布格局均以随机分布为主。(3)不同纬度各群落物种多度与空间聚集程度之间存在负相关关系,物种多度对空间聚集程度的影响随纬度从高到低逐渐减弱。(4)较高纬度群落物种胸径与空间聚集程度之间存在负相关关系;随纬度从高到低,物种胸径对空间聚集程度影响水平逐渐降低。对亚热带不同纬度森林群落物种空间分布格局形成原因的进一步探讨,认为由纬度差异引起的生境异质性是影响群落物种空间分布格局产出过程的主要因素,同时林型、扩散限制、密度制约和随机作用是影响各群落物种空间格局及多样性维持的次要因素。本研究通过不同纬度群落物种空间分布格局的分析和讨论,揭示了亚热带森林群落物种空间分布格局类型及其成因,为该区域的生物多样性保护工作提供了依据。
致谢:中国科学院华南植物园黄忠良研究员、曹洪麟研究员对样地调查数据给予支持, 广东鼎湖山森林生态系统国家野外科学观测研究站、猫儿山国家自然保护区提供帮助,特此致谢。
