喜旱莲子草在增温背景下的密度制约与生长权衡
2022-12-16王渟渟张桥英张运春唐梦娥
王渟渟,张桥英,张运春,*,唐梦娥,廖 望
1 西华师范大学环境科学与工程学院, 南充 637009
2 西华师范大学地理科学学院, 南充 637009
人类活动与工业生产使大气中二氧化碳浓度急剧增加,进一步加快全球气候变暖进程[1]。据IPCC第五次综合评估报告显示,截至2100年,全球地表温度将上升0.3—4.8℃。中国地表温度将上升2.2—4.2℃[2]。植物的生长发育离不开有效积温[3—4]。增温影响植物的生长策略与功能性状,改变物种竞争态势,进而引起种群结构和植物分布格局的变化。例如,气候变暖使甲虫向高纬度扩张,随纬度升高,喜旱莲子草与甲虫的相互作用增强,喜旱莲子草种群生长受限,覆盖度减少,种群数量降低[5]。增温促进天敌活动间接减弱外来植物入侵强度,改变喜旱莲子草种群数量与分布范围[5—6]。加拿大一枝黄花在增温作用下加大对繁殖构件的投资,种群出生率增加死亡率降低,种群数量增大,有助于占据更高生态位,从而成功入侵[7]。
密度制约调节机制反应了个体与种群数量的动态变化,是研究个体大小与密度关系的重要内容[8—9]。随密度压力增大,个体所获得的矿质营养与空间资源减少,植物发生可塑性调整,种群数量动态变化,个体生育率降低,死亡率增加,直到种群数量达到稳定状态[9—12]。对种群密度调控机理的研究已取得一些进展。Yoda等以草本植物为研究对象,基于植物等速生长的假设,提出-3/2自疏法则[13],得到植株个体平均生物量与种群密度的函数关系。Enquist等从生物能量学角度出发,基于植物异速生长的假设,推导出-4/3自疏法则[14—15],得到植株个体平均生物量与最大种群密度的异速比例关系。种群个体的竞争实质上是各构件对可获得性限制资源的争夺[16]。地上光资源与地下矿质资源、水资源的不同,导致植物地上、地下竞争机制有本质区别[17]。同时各构件特异的物理空间及微环境增强了地上、地下竞争机理的差异。如红葱种群地下构件生物量超过环境最大容纳量,地上构件生物量恰好达到环境最大容纳量,地下资源的竞争占主导地位[8];胡萝卜地上部分产量与密度呈渐近关系,根系产量与密度呈抛物线关系[18]。以往有关北美车前、沙米、红葱等响应种群密度的调控机理的研究多侧重于常温下植物的生长权衡,而对全球变暖背景下,密度制约与植株生长的关系、地上和地下部分生物量与种群密度的关系还缺乏足够的认识。
植物权衡机理的研究对了解植物资源配置策略与生理整合模式,解释植物生长发育的环境适应性至关重要[19—20]。植物的权衡生长分为结构特性的权衡和生物量分配的权衡[21]。密度制约影响植物资源的获取能力与能量吸收利用效率[22]。使植株改变各构件可获得性资源分配比例,权衡各功能构件能量分配,及时调整构件大小与表型可塑性来响应密度变化[23—24]。例如,随种群密度增加, 红葱叶片数减少,叶生物量减小,通过延长叶片长度使叶生物量得以积累,以此适应空间与资源的限制[8];猪毛菜扩大种群密度,根长与分枝数减少,其对生活史的权衡分配策略为减少资源对分枝的分配以适应不同密度竞争环境[25]。
喜旱莲子草是原产于南美洲的宿根草本植物。因其极强的适应性、忍耐性、抗逆性,极易在异质生境中大面积爆发,形成单优势种喜旱莲子草群落。是一种广泛分布于陆地和水域环境的广布性物种[26]。以往对喜旱莲子草的研究大多集中于单一或多种因素处理下的光合生理特性[27]、叶片特性[28]、种间或种内竞争[26,29]、表型可塑性[30]等方面。然而在全球增温背景下,喜旱莲子草为响应密度变化如何权衡构件特征,常温与增温处理下喜旱莲子草响应密度变化的调节机制是否发生变化尚未明确。因此深入研究增温背景下,植物构件、地上、地下的生长权衡策略以区分各构件功能差异。用异速生长分析种群密度、个体大小和各构件间关系,以排除个体大小的影响,揭示各构件特异的生物量-密度关系[31]。进一步理解种内竞争下喜旱莲子草对异质性资源利用特性与环境适应机制。
为此,本文在全球气候变暖的背景下,研究密度制约对喜旱莲子草形态特性与生物量的权衡特征;各构件响应不同种群密度的变化规律,不同温度下种群密度与生物量的异速关系;不同温度处理下密度制约对喜旱莲子草影响的差异。以揭示全球变暖背景下入侵植物构件水平、个体水平响应密度变化的适应对策,丰富入侵植物密度制约理论,为进一步解析喜旱莲子草入侵繁衍机理提供科学依据。
1 材料与方法
1.1 研究地概况
实验地位于四川省南充市西华师范大学环境科学与工程学院实验基地。地理位置为30°23′N,106°02′E。海拔276 m。冬季温和少雨,夏季炎热多雨,属亚热带湿润季风性气候。年平均气温17.5℃左右,年降水量约1020.8 mm,年日照时数1354.7 h[27,29,32]。
1.2 实验设计
采用密度和温度双因素实验设计。对密度采用加性实验设计[33],构建不同密度喜旱莲子草种群。分别为低密度处理(LD):1株/盆(37株/m2);中密度处理(MD):5株/盆(186株/m2);高密度处理(HD):9株/盆(335株/m2)。其中中密度与高密度实验盆中的植株围绕中心喜旱莲子草均匀种植。温度处理设两个梯度。分别为增温处理(HT):选用温室棚模拟增温,温度控制在高于室外温度2℃左右;常温处理(NT):在与温室棚同材质的有顶无壁棚内进行实验,选用样地的自然生境温度。实验共设置6个处理(表1),每个处理5个重复。

表1 实验设计和处理代码
喜旱莲子草于2019年3月底采集于实验基地附近,并将其扦插于培养箱进行为期14 d的适应性培养。移栽时,选择外部形态和长势大致相同的喜旱莲子草幼苗(高约8 cm)同时移植于口径9.25 cm、高11 cm的圆形塑料花盆,花盆中培养基质为混合土(营养土∶紫色土=4∶1;营养土中含少量泥炭、珍珠岩、草炭、蛭石等),并添加适量缓释肥混匀。每个花盆装土约2 kg。实验周期为60 d,实验期间每隔1周将样地中心位置的花盆和样地边缘位置的花盆对换位置,确保每盆植株受光均匀,各盆管理措施一致。
1.3 参数测定
2019年6月下旬,以盆为单位,测量每盆喜旱莲子草的根、茎、叶特征数据,测量过程中尽量避免损坏喜旱莲子草的根系。分别统计每盆植株的茎节数、茎分枝数、叶量;对每盆植株随机选取10片叶子,扫描仪扫描后用ImageJ测量其叶面积;用直尺测量植株的茎长;用根系扫描仪对根系扫描,WinRHIZO根系分析系统分析根长、根表面积、根平均直径、根体积。
把喜旱莲子草的根、茎、叶分别装袋标记,并带回实验室,置于65℃下烘24 h至质量恒重,测量植株各构件生物量。
1.4 数据处理与分析
实验数据应用SPSS 23.0软件进行分析处理。对数据进行正态分布和方差齐性检验,并对部分数据进行恰当的数据转换。用双因素方差分析(Two-way ANOVA)检验不同种群密度和温度对喜旱莲子草形态特征和生物量分配的影响。用SNK法进行多重比较,分析不同处理下喜旱莲子草各数量性状差异的显著性。用Origin 2019b软件绘图。
对喜旱莲子草异速比例关系的分析用经典异速方程Y=KNγ来描述。对分析数据进行log10转换。其中,方程斜率γ为异速指数,方程与Y轴的截距K为异速常数,Y是种群中存活个体的平均生物量,N是种群密度[22]。当γ<-1时,种群生物量积累大于环境最大容纳阈值;当γ=-1时,种群生物量积累等于环境最大容纳阈值;当γ>-1时, 种群生物量积累小于环境最大容纳阈值[34]。采用减小主轴回归(Reduced Major Axis regression, RMA)拟合数据,进行回归分析。确定异速指数:αRMA=αOLS×r-1(αOLS是最小二乘法得到的回归系数;r是相关系数),以去除变量偏差对回归系数的影响[33]。
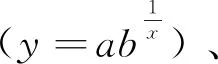
2 结果分析
2.1 不同温度下喜旱莲子草形态特性的密度制约
温度和种群密度对喜旱莲子草的茎长、茎节数、茎分枝数、叶量有显著影响,比叶面积在不同种群密度处理下有显著差异(P<0.05),温度和种群密度的交互作用对茎长、茎节数、茎分枝数、叶量有极显著影响(P<0.01,图1)。在喜旱莲子草的形态特性中,除了根长、根表面积、根平均直径、根体积外,茎长、茎节数、茎分枝数、比叶面积、叶量与密度有极显著线性函数关系(P<0.01,表2)。随种群密度增加,茎长变短,茎节数与茎分枝数变少,叶片数量递减,比叶面积递增。常温处理下,低密度喜旱莲子草的茎长、茎节数、茎分枝数、叶量分别比高密度的喜旱莲子草茎长、茎节数、茎分枝数、叶量增加248.1%、219.2%、652.9%、288.3%,而比叶面积减少77.8%。较高的种群密度限制了喜旱莲子草地上指标的生长,种群密度处理对喜旱莲子草地下指标影响不显著(P>0.05)。
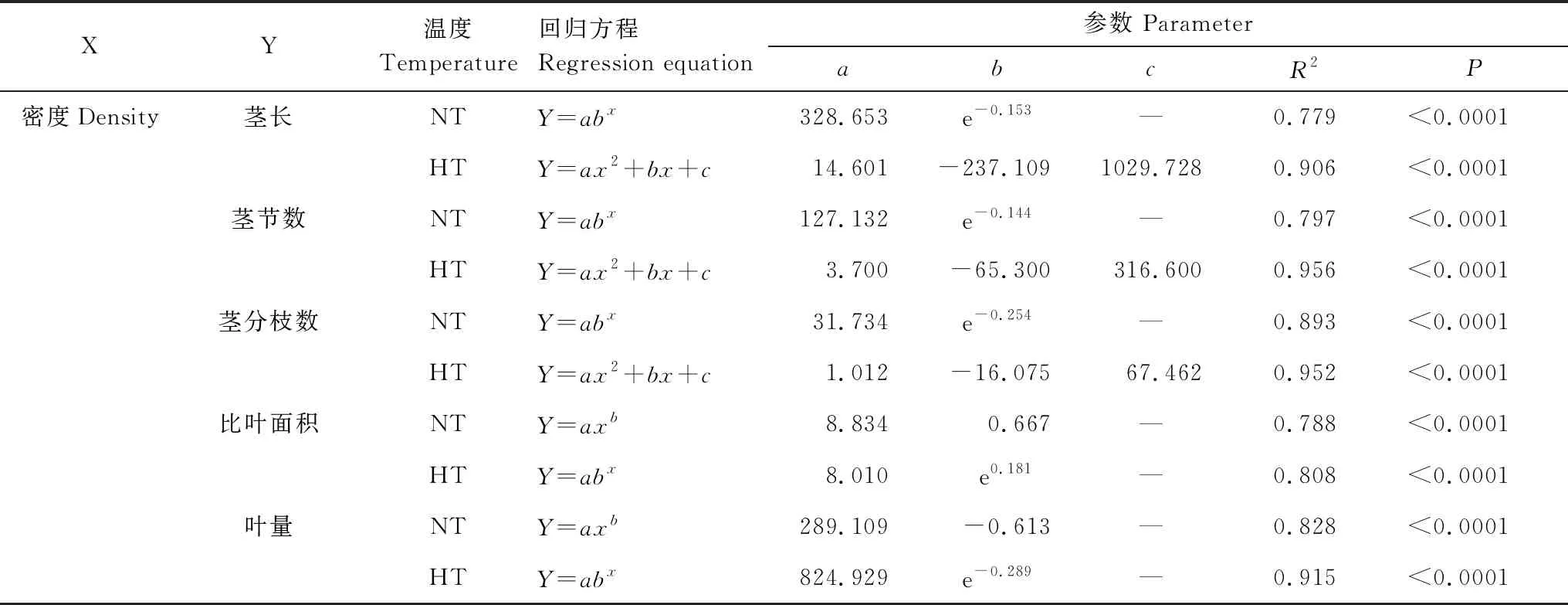
表2 不同温度下喜旱莲子草地上指标和生物量与密度之间关系的拟合模型
增温对喜旱莲子草的茎长有显著影响,温度对低密度下的茎节数、茎分枝数有显著差异。叶量只在低密度与中密度处理下受温度调控(P<0.05,图1)。增温促进了茎分枝数生长,常温处理下喜旱莲子草的茎长、茎节数、叶量在低密度与中密度时均不同程度的低于增温处理,高密度时变化趋势相反。增温在低密度与高密度时对比叶面积有促进作用,分别增加了5.2%、6.1%。中密度时比叶面积随温度升高逐渐降低。在低竞争强度下,增温对喜旱莲子草的茎长、茎节数、茎分枝数、叶量较中、高密度处理有显著促进作用,茎长、茎节数、茎分枝数、叶量分别增加了165.3%、125.7%、104.7%、123.1%。中、高竞争强度下,温度对喜旱莲子草的茎节数、茎分枝数、叶量促进作用不显著(P>0.05,图1)。温度与种群密度处理对喜旱莲子草地上指标的影响显著高于地下指标。温度、种群密度及其交互作用均会影响喜旱莲子草的形态特性。但种群密度处理的喜旱莲子草形态特性的变化值显著高于温度处理,种群密度对喜旱莲子草的形态特性的可塑性影响更大。
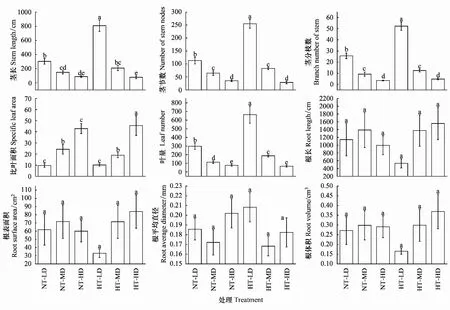
图1 喜旱莲子草在不同培养环境下生长指标变化趋势
2.2 不同温度下喜旱莲子草生物量的密度制约
喜旱莲子草各构件平均生物量、地上生物量和个体生物量均受温度调控,且表现出极显著的密度制约特性(P<0.05)。随种群密度的增加各构件平均生物量、地上生物量和个体生物量均逐渐降低(图2)。不同种群密度对喜旱莲子草生物量影响不同(表3)。常温处理下,随种群密度增大,茎、叶、根构件平均生物量、地上生物量、个体生物量异速指数分别为γstem=-1.605、γleaf=-1.685、γroot=-1.524、γabove-ground=-1.612、γtotal=-1.558(表4,P<0.01)。茎、叶、根构件平均生物量在低密度时取得最大值,是高密度种群根、茎、叶构件平均生物量的32.9倍、41.2倍、19.7倍。增温处理下,竞争强度增大,茎、叶、根构件平均生物量以异速指数γstem=-2.075、γleaf=-1.933、γroot=-1.800 递增;地上生物量、个体生物量以异速指数γabove-ground=-2.038、γtotal=-1.982有规律递增。无论常温还是增温处理,γroot、γstem、γleaf、γabove-ground、γtotal均小于-1,表明种群各构件生物量、地上生物量、个体生物量已经超过环境最大容纳阈值。
低密度时,温度对茎、叶构件平均生物量有显著差异(P<0.05),对根构件平均生物量无影响(P>0.05)。中密度时,增温对茎、根构件平均生物量有显著影响(P<0.05),叶构件平均生物量不受温度调控(P>0.05)。高密度时,仅根构件平均生物量受温度显著调控(P<0.05,图2、表3)。常温处理的茎、叶、根构件平均生物量在低密度与中密度时均大于增温处理。在高密度时,茎、根构件平均生物量随温度升高呈降低趋势,而叶构件平均生物量变化趋势相反。随温度的升高,茎平均生物量在低密度、中密度下分别增加了133.2%、66.2%、高密度下降低了17.0%;叶平均生物量在低密度、中密度、高密度下分别增加了71.8%、13.6%、2.2%;根平均生物量在低密度、中密度下分别增加了33.1%、10.2%,在高密度下降低了46.7%。增温对高密度下的喜旱莲子草有抑制作用,茎、叶、根构件平均生物量均随温度升高而降低。各构件平均生物量在增温低密度下取得最大值(图2)。增温与种群密度均影响喜旱莲子草的生长,其中种群密度对喜旱莲子草生长限制作用更为显著。
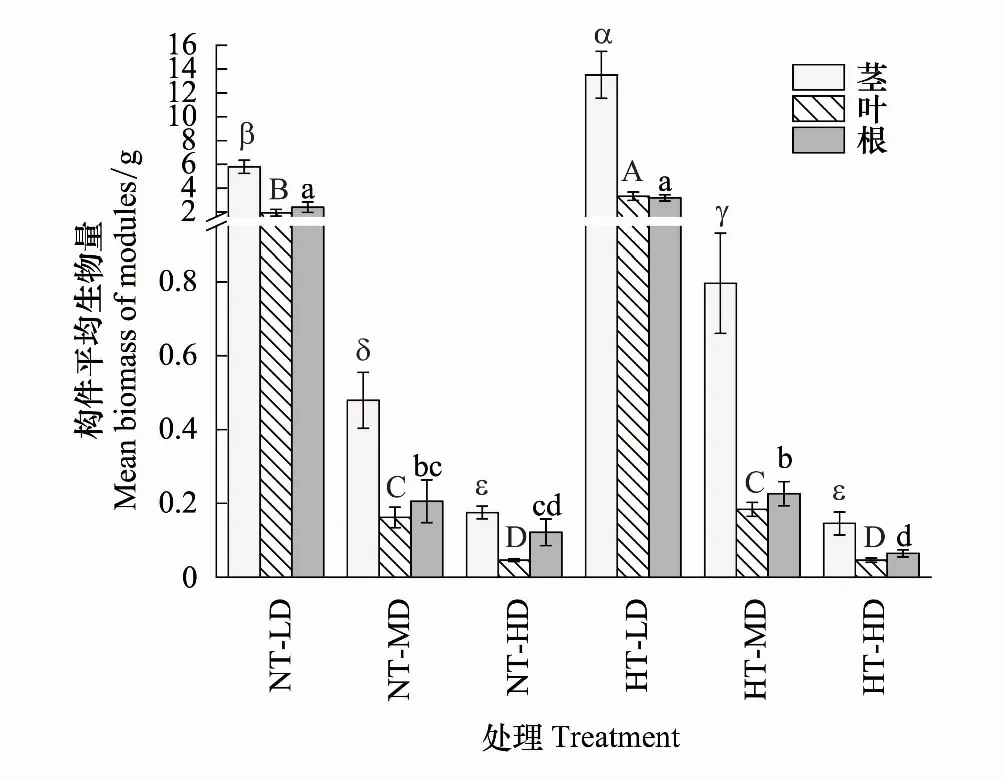
图2 不同培养环境对喜旱莲子草各构件生物量的影响
2.3 不同温度下喜旱莲子草生物量分配的密度制约
温度与种群密度及其交互作用对喜旱莲子草茎分配比、根分配比、根冠比有显著影响(P<0.05),叶分配比不受其调控(P>0.05,表3、图3)。随种群密度增加,茎分配比呈下降趋势,根冠比和根分配比呈上升趋势。常温处理下,叶生物量分配比例先增加后减少,与增温处理变化趋势相反。根分配比在低密度与中密度下无显著差异。茎、叶构件在高密度时生物量分配比例最小,而根冠比、根分配比在高密度时取得最大值。增温条件下,茎分配比最大,根冠比与根分配比在低密度最小。高密度条件下茎构件生物量分配比例最小,根、叶构件生物量分配比例和根冠比均取得最大值。喜旱莲子草在低密度处理的茎分配比比高密度处理高20.1%,说明无论增温与否,随竞争强度加大,喜旱莲子草能量分配更侧重地下部分,且生物量优先向根构件分配。高密度处理下的叶分配比、根冠比、根分配比分别比低密度处理增加了13.2%、73.6%、53.8%。
随温度升高,根冠比逐渐降低(表3)。低密度处理下,喜旱莲子草茎生物量分配比例随温度的升高而增加(表3、图3)。在低密度与中密度处理下,增温促进叶生物量分配比例增加。而高密度下,增温使叶生物量分配受阻,叶分配比减小。根冠比、根分配比随温度的升高而降低。增温处理下喜旱莲子草对地上部分的投资大于地下部分。中密度处理下,增温处理的喜旱莲子草茎生物量分配比例增加了13.8%,叶分配比、根冠比、根分配比分别比常温处理降低了20.9%、20.1%、15.4%。高密度下增温,茎相对生物量与叶相对生物量增加,根冠比与根相对生物量减小。增温使喜旱莲子草加大对地上部分的投资以适应环境的异质性。
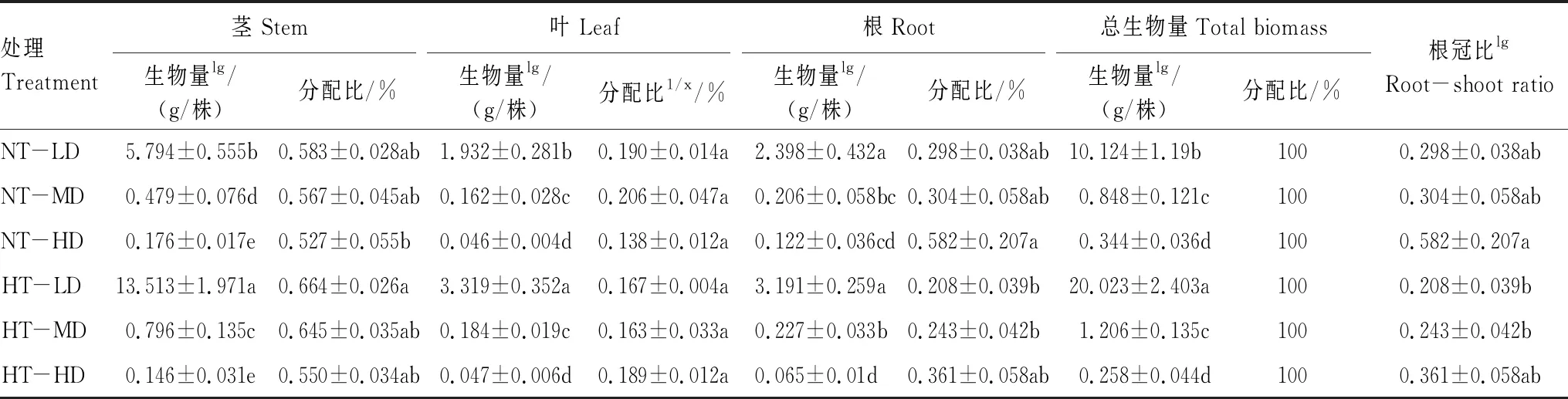
表3 喜旱莲子草不同培养环境的生物量与分配比
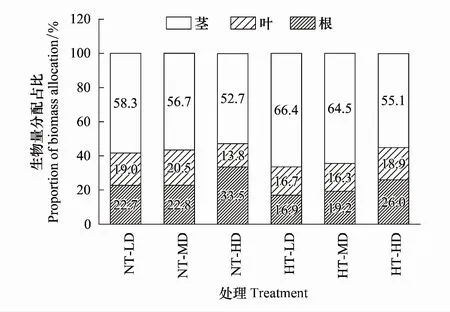
图3 不同处理下喜旱莲子草生物量分配比例
2.4 不同温度下喜旱莲子草生物量与密度的异速关系
常温处理下,平均茎、叶、根器官生物量、地上生物量、个体生物量与密度的异速关系指数γ分别为-1.605、-1.685、-1.524、-1.612、-1.558(表4)。从95%置信区间可知,γstem、γleaf、γabove-ground、γindividual与-3/2无显著性差异(P>0.05),茎生物量、叶生物量、地上生物量、个体生物量与密度的异速关系遵循-3/2自疏法则;γroot与-4/3、-3/2无显著性差异(P>0.05),说明根生物量与密度的关系遵循-3/2自疏法则和-4/3自疏法则。
增温处理下,平均茎、叶、根器官生物量、地上生物量、个体生物量与密度的异速指数γ分别为-2.075、-1.933、-1.800、-2.038、-1.982(表4)。由95%置信区间可知,γstem、γleaf、,γroot、γabove-ground、γindividual小于-1、-4/3、-3/2(P<0.05)。说明各构件生物量、地上生物量、个体生物量与密度的异速关系不遵循-3/2、-4/3自疏法则,且和最终产量恒定法则无关。各器官生物量、地上生物量、个体生物量在常温与增温处理下都与密度呈极显著负相关关系(P<0.01)。
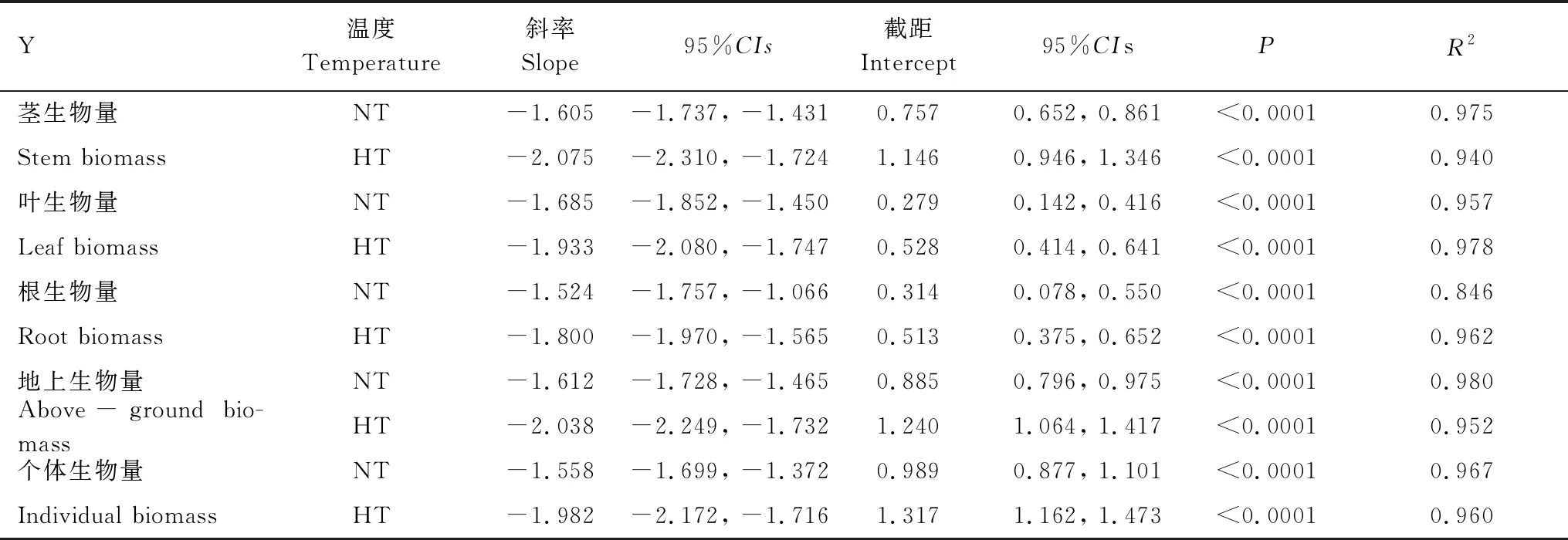
表4 不同温度下喜旱莲子草构件生物量、地上生物量、个体生物量与密度之间异速比例关系
3 讨论
3.1 喜旱莲子草形态特性的权衡
除了地下指标不受种群密度的影响外,喜旱莲子草地上构件的各个特征(茎长、茎节数、茎分枝数、叶量)和比叶面积表现出显著的密度制约特性。密度的负效应在一定程度上决定了各构件限制性资源的获取,植株生长受阻,此时改变形态结构是最高效最经济的适应对策[35]。植物为躲避邻体植株的遮阴干扰,减弱分株密度,获得更多光资源,株高一般会随密度增加而增高[30,36]。本研究中,随密度增加,喜旱莲子草茎长变短,可能是通过减少茎长伸长来减慢植株生长速度增强竞争胁迫的抗逆性。茎节数、分枝数、叶量减少,可能是邻体植株的横向与纵向伸长形成了荫蔽环境,植物可获得性光资源受限,此时通过形态可塑性调整,减少构件的生长以降低营养资源的分配和生存消耗,提高植物适合度[37]。这与冯银平等对紫花苜蓿的研究结果一致[38]。低密度种群生存压力小,空间充足,内部通风性能好,单位质量叶片获得的光资源较多,光合速率增强,比叶面积减小,叶片数和侧枝数增多[39]。高密度种群资源压力大,个体间相互干扰,植物偏向于减少叶面积,降低植株直径,减缓株高径向生长,增加对邻体胁迫的忍耐性[40]。
除了比叶面积和地下指标不受温度的影响外,喜旱莲子草其他数量性状与温度呈显著的正相关关系。增温促进外来入侵植物在异质生境扩散[41],促使茎构件生长,扩大植株的横向宽度与纵向高度,促进叶构件对光能的截取,积累更多干物质,缓解分株内部的资源压力,有利于种群的稳定与扩张。说明植株响应温度变化调整形态特征,迅速占据生态位扩宽生态幅,最大限度捕获潜在可利用资源,提高环境适应性。温度与种群密度及其交互作用对喜旱莲子草地下指标影响不显著。
3.2 喜旱莲子草生物量积累的权衡
在生境资源一定的情况下,种群密度对喜旱莲子草茎、叶、根生物量均表现出显著的负效应[42]。相邻植株生态位相同,个体各构件为争夺必需资源,构件生物量发生可塑性变化,竞争力弱的植株构件生长速率减慢,生物量减小[22]。本试验中,喜旱莲子草各构件生物量随密度增加逐渐降低,表现出负密度制约现象。密度压力增加,资源压力增大,有限资源获取受限,导致各构件生长受阻,植株总生物量下降,各构件生物量也随之降低。姚杰等对水曲柳和黄檗的研究也支持这一结论[43]。各构件生物量随密度变化速率略有不同。用生物量-密度异速指数γ衡量种群密度调控强弱。常温处理下,γleaf(-1.685)<γabove-ground(-1.612)<γstem(-1.605)<γindividual(-1.558)<γroot(-1.524),叶受密度制约最强,根受密度制约最弱。喜旱莲子草对地上资源竞争占主导地位。这是因为随植物生长,各构件受可获得性光资源比率限制,叶片对光资源的竞争更加激烈,冠层底部叶片受植株内部及邻体植株的遮荫,生长受抑制[42],因此叶片受到更强的密度制约。γroot与-4/3、-3/2无显著性差异(P>0.05),说明根生物量与密度的关系遵循3/2自疏法则和-4/3自疏法则;γstem、γleaf、γabove-ground、γindividual与-3/2无显著性差异(P>0.05),茎生物量、叶生物量、地上生物量、个体生物量与密度的异速关系遵循-3/2自疏法则。增温处理下,γstem(-2.075)<γabove-ground(-2.038)<γindividual(-1.982)<γleaf(-1.933)<γroot(-1.800),因此,各构件大小、地上大小、个体大小随密度变化不一致。受密度制约调控强度的大小依次为:茎>地上>个体>叶>根。喜旱莲子草种群地上构件受密度制约作用更强。可能是地上地下资源不同导致地上地下竞争机制差异;且随密度增加,喜旱莲子草以牺牲茎为代价,将更多光合产物分配给根,茎构件对资源竞争能力增强,因此受密度压力调控更大。由各器官自疏斜率、地上部分自疏斜率和植株个体自疏斜率的95%置信区间看出,其斜率均小于-1、-3/2和-4/3。所以都不遵循-3/2、-4/3自疏法则和最终产量恒定法则。
密度制约理论将种群密度与种群中存活个体平均生物量动态关系用异速方程表示(Y=KNγ)。本研究中,无论常温还是增温处理,,各构件生物量-密度关系的异速指数γ均小于-1,说明喜旱莲子草种群茎、叶、根生物量的积累已超过环境最大容纳阈值,地上部分的光资源、地下部分的矿质营养与水分、地上地下环境空间已被充分利用,种群达到最大资源利用率,喜旱莲子草的总产量随密度的增加而减少。密度制约对各构件作用强度不同,导致各构件获得的光合产物不一致。这可能是引起喜旱莲子草各构件生物量-密度差异的原因。
3.3 喜旱莲子草生物量分配的权衡
各构件生物量分配比例的变化反映了植物在异质生境的资源获取以及能量分配的调整[42]。植物通过改变生产力,权衡光合产物在各构件中的分配占比,以提高资源的利用效率[38,44]。本研究中,除叶生物量分配占比在温度与种群密度调控下无显著差异外,茎、根生物量分配占比对温度与种群密度有积极响应。随种群密度增加,营养资源与环境空间的竞争压力增强,喜旱莲子草对各构件生物量分配进行可塑性调整。在中密度与高密度下,茎构件的能量投资减少,叶生物量分配在各密度下无显著变化,光合产物优先向根构件分配,根冠比增加。说明低竞争强度下,植株侧重于将能量分配给支持器官。这可能是R/FR比值减小,光有效性降低,气孔导度减少,植物固定的CO2少,生长速率减慢[45],因此对营养物质及水分的需求少,植物对地上光资源竞争能力增强。此时植物以牺牲根为代价将更多能量分配给茎。茎构件产生支撑结构,一方面保持植物机械稳定,另一方面最大限度占据地上空间捕获地上资源;中、高竞争强度下,植物偏向于将能量分配给资源获取器官。这是由于资源压力大,水分与矿质营养成为限制性资源,植物对地下资源竞争能力增强。植物减少茎生物量分配,增加根生物量分配以充分利用地下资源,维持植株生存。这支持了Poorter等人的结论[46]。Schimitt & Wulff认为植物茎生物量分配比例增加是以叶、根生物量分配比例减小为代价[47]。本研究发现,随种群密度增加,茎生物量分配下降,叶生物量分配在各密度下维持恒定,根生物量分配上升。这可能是因为喜旱莲子草对土壤养分、光照强度、环境空间的竞争能力随密度增加而增强。喜旱莲子草增加对根构件的能量投资,这与最优化理论中水资源受限情况一致。叶生物量随密度增加而下降,叶生物量分配不受温度与种群密度的影响,这与上述中Schimitt & Wulff提出的叶生物量分配比例减小不一致。可能是研究的目标物种不同,密度压力增大,植株总生物量下降导致叶生物量减小。说明随竞争强度增大,喜旱莲子草选择增大根生物量分配,邻体间对地下资源的竞争逐渐变得“对称”,对光资源的竞争逐渐变得“不对称”[48],种群地下构件对资源竞争能力逐渐增强。密度压力增大,喜旱莲子草茎生物量和茎生物量分配明显大于根生物量和根生物量分配,叶、根生物量分配的上升趋势是以茎生物量分配的下降趋势为代价。这是因为在较低的密度压力下,根系提高对水分与矿质元素的吸收利用效率,将能量通过茎构件输送到地上,同时叶构件充分利用光照合成更多光合产物,促进茎生物量的分配,这与范高华等人的结果类似[42]。在较高的密度压力下,环境空间有限,地下资源竞争激烈,且邻体植株的拥挤效应使叶片截获的光资源减少,叶构件光合作用效率降低[49]。喜旱莲子草为维持生存,植株积累的干物质优先向叶构件和地下分配。范高华也证实,随种群密度增大,地上生物量的分配逐渐减小[42]。不同密度下各构件生物量分配占比的差别可能是造成各构件特异的生物量-密度关系的原因。
4 结论
综上所述,增温促进喜旱莲子草生长,密度制约使喜旱莲子草生长受阻,增温在一定程度上削弱了密度的负效应。随种群密度压力增大,喜旱莲子草种群地下资源的竞争能力增强,地上资源竞争能力减弱,喜旱莲子草地上部分调节强于地下部分,常温处理下,叶受密度制约最强,增温处理下,茎受密度制约最强。地上资源的竞争占主导地位。
