丘脑背内侧核损毁对帕金森病大鼠内侧前额叶皮层电活动的影响
2022-12-15范玲玲丁书平沈国民胡志红任爱红
范玲玲, 丁书平, 沈国民, 胡志红, 任爱红, 邓 博
(1. 河南科技大学基础医学院生理教研室, 河南 洛阳 471000;2. 河南科技大学基础医学院 河南省血栓与止血国际联合实验室, 河南 洛阳 471000;3. 洛阳职业技术学院公共基础教学部, 河南 洛阳 471023)
帕金森病(Parkinson’s disease, PD)主要是由于中脑黑质多巴胺神经元变性脱失导致纹状体去多巴胺能神经支配, 从而改变基底神经节的神经环路, 使PD 患者控制随意运动的能力丧失, 震颤、强直、动作迟缓和步态异常等是PD 患者运动障碍的主要临床表现。研究[1]显示: PD 患者中普遍存在以抑郁、焦虑和认知功能障碍为突出表现的非运动症状。然而, PD 患者非运动症状的病理生理学基础尚不明确。PD 患者的黑质-纹状体通路变性会导致多个脑区功能和(或)结构的变化[2]。丘脑背内侧核(mediodorsal thalamic nucleus, MD) 是丘脑的一个中继核团, 与基底神经节环路中的苍白球和纹状体等部位存在复杂的纤维联系, 参与PD 非运动系统症状发生机制的前额叶皮层(prefrontal cortex, PFC)与MD 之间也有大量的交互纤维联系。MD 参与认知功能、情感和学习记忆等方面的神经调控。研究[3-4]显示:由意外伤害、肿瘤和退行性疾病引起MD 损伤会导致痴呆和健忘症。发生PD 时所出现的认知功能障碍、睡眠障碍、抑郁和嗅觉功能障碍等与发生MD 功能障碍时的临床表现相似。研究[5-6]显示:MD 形态改变和代谢变化与抑郁有直接关联。神经影像学研究[7]显示:与非抑郁的PD 患者比较, 有抑郁症状的患者左侧MD和内侧前额叶皮层(medial prefrontal cortex, mPFC) 的活动减弱, 同时其双侧MD 体积增大, 神经元数量增加。研究[8]显示:PD 状态下MD 神经元的电活动增强, 提示MD 可能涉及PD 患者的非运动症状。mPFC 是一个精细和综合的认知加工的最高级解剖功能区, mPFC 不直接接受中枢神经系统之外的纤维投射[9], mPFC 中80%的神经元为谷氨酸能锥体投射神经元。对于许多种属的动物来讲, PFC 在情绪调控和认知过程中均发挥重要作用, mPFC 的功能异常与智能和情感障碍以及PD 非 运 动 症 状 的 发 生 有 关[10-11]。MD 是 处 理 进 出不同PFC 区域的神经信息的核心结构[12], 提示MD 可能参与了PFC 的重要生理功能。MD 的功能改变可能与PD 状态下mPFC 功能障碍的发生有一定关系, 但是尚未见有关MD 在PD 病理生理过程中的具体作用的报道。本文作者观察毁损MD 后, 正常和PD 模型大鼠mPFC 锥体神经元电活动的变化, 探讨MD 核团对PD 病理过程的影响, 为进一步阐明其在PD 非运动症状中的作用提供依据。
1 材料与方法
1.1 实验动物、主要试剂和仪器24 只健康、雄性SD 大鼠, 体质量270~350 g, 购自西安交通大学实验动物中心, 动物使用许可证号:SYXK(陕)2020-005。在标准环境下饲养, 室温(21±2)℃, 24 h 昼夜循环光照, 自由摄食、饮水。大鼠随机分为正常组(n=6)、黑质致密部(substantia nigra pars compacta, SNc)损毁组(n=6)、MD损毁组(n=6) 和SNc+MD 联合损毁组(n=6)。所有实验均遵循西安交通大学实验动物管理和使用的相关规定, 并尽量减少实验动物的使用数量, 减轻动物的痛苦。
盐酸6- 羟基多巴胺(6-hydroxydopamine, 6-OHDA)、盐酸阿扑吗啡、鹅膏蕈氨酸(ibotenic acid, IBO) 和盐酸地昔帕米均购自美国Sigma 公司, 盐酸阿扑吗啡和盐酸6-OHDA 均溶于生理盐水(含0.02% 抗坏血酸), IBO 溶于PBS 缓冲液, 盐酸地昔帕米溶于双蒸水。SN-2N 动物脑立体定位仪购自日本Narishige 公司, AVB-11A 生物物理放大器、MEZ-8301 微电极放大器和VC-11 双线记忆示波器购自日本Nihon Kohden 公司, CED1401 Spike2 生物电信号采集与分析系统购自英国Cambridge Electronic Design 公司, WD-Ⅱ型恒温控制操作台购自西安万东仪器有限公司。
1.2 PD 模型的制备PD 动物模型采用6-OHDA单侧毁损大鼠中脑SNc 的方法制备。术前30 min先给予大鼠注射地昔帕米(25 mg·kg-1)保护去甲肾上腺素神经元。采用4%水合氯醛(400 mg·kg-1)麻醉大鼠, 将其头部固定于立体定位仪上, 将6-OHDA(2 g·L-1)注射至右侧SNc(坐标位置:AP -5.0~5.3 mm, L 1.9~2.1 mm, D 7.1~7.3 mm), 给药速度为0.5 μL·min-1, 注射完毕微量注射器留置5 min 后缓慢退出。术后1 周, 给予大鼠阿扑吗啡(0.05 mg·kg-1), 选择每5 min 向对侧旋转超过20 圈的模型大鼠进行后续实验。
1.3 IBO损毁MD采用4%水合氯醛(400 mg·kg-1)麻醉大鼠后, 头部固定于立体定位仪上, MD 三维坐标 位 置 为AP -2.8 mm, L 0.5~0.6 mm, D 5.3 mm(根据Paxinos-Watson 的大鼠脑图谱)。开颅, 将1 μL 微量注射器的尖端与玻璃微电极密闭相连, 缓慢进针, 在MD 的坐标位置注入0.3 μL IBO溶液(10 g·L-1), 速度为0.1 μL·min-1, 注射完毕后将微注射器留置5 min, 缓慢退出, 以免药物逆流, 缝合皮肤, 继续置入笼中喂养。术前和术后皮下给予大鼠8 万单位青霉素以防感染。PD 模型大鼠的MD 毁损在毁损SNc 1 周后进行, 且在损毁SNc 的同侧损毁MD。
1.4 在体神经元电活动观察分别在6-OHDA 毁损SNc 3 周后、MD 毁损2 周后和SNc+MD 联合损毁3 周后(SNc 损毁1 周+MD 损毁2 周后) 进行mPFC 锥体神经元电活动记录。4% 水合氯醛(400 mg·kg-1)麻醉大鼠, 气管和颈外静脉插管手术后, 采用电热垫使大鼠体温维持在(37.0±0.5)°C。采用细胞外记录法记录mPFC 锥体神经元的电活动, 玻璃微电极充灌0.5 mol·L-1醋酸钠溶液(含2%滂胺天蓝), 按大鼠立体定位图谱定位至mPFC:AP 2.7~3.4 mm, L 0.5~1.0 mm, D 1.5~4.0 mm, 电极尖端阻抗维持在8~12 MΩ。为了更接近动物正常状态, 电生理记录在稳定的浅麻醉状态下进行。细胞放电通过微电极放大器在VC-11 双线记忆示波器上显示, 观察神经元放电的波形和形式, 同时输入音箱进行监听。将稳定的、信噪比大于3 的神经元放电输入计算机分析软件, 进行在线或离线分析。当记录到稳定的细胞放电后, 持续采样5~10 min。记录放电频率、放电间隔(interspike interval, ISI) 的 变 异 系 数 和 放 电形式。
1.5 免疫组织化学染色观察酪氨酸羟化酶免疫阳性神经元数在神经元电活动记录完毕后, 将玻璃微电极内的滂胺天蓝电泳(-20 μA, 15 min)至最后一个记录位点。过量麻醉大鼠后灌注固定, 取脑组织并进行连续冰冻冠状切片, 采用酪氨酸羟化酶的免疫组织化学染色确定SNc 和腹侧被盖区(ventral tegmental area, VTA)中多巴胺神经元损毁情况。尼氏染色判断电生理记录点的位置和MD损毁情况。
1.6 统计学分析采用SPSS 20.0 统计软件进行统计学分析。各组大鼠平均旋转圈数、SNc 和VTA 中酪氨酸羟化酶免疫性神经元数、锥体神经元平均放电频率和ISI 的变异系数以±s表示, 各组大鼠锥体神经元放电频率比较采用单因素方差分析, 平均ISI的变异系数比较采用Mann-WhitneyU检验, 各组神经元放电形式分布比较采用χ2检验。6-OHDA 损毁大鼠损毁侧与未损毁侧SNc 和VTA 中酪氨酸羟化酶染色阳性细胞数比较采用配对t检验。以P<0.05 为差异有统计学意义。
2 结 果
2.1 mPFC 锥体神经元的放电特征和电活动记录位点mPFC 锥体神经元的电生理学特征见图1A。所有记录位点被滂胺天蓝标记, 经尼氏染色确定位于mPFC 中(图1B)。6-OHDA 损毁SNc 的大鼠皮下注射阿扑吗啡后均向损毁侧的对侧旋转, 且每5 min 旋转圈数大于20 圈, 平均旋转圈数为(43±4)圈。
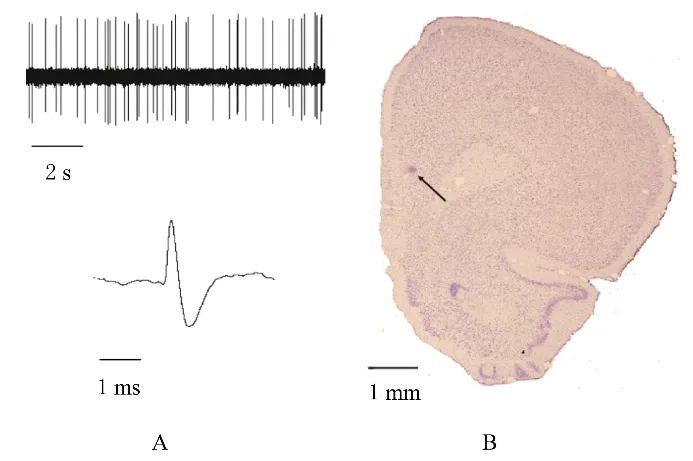
图1 大鼠mPFC 中锥体神经元和神经元记录位点的放电特征Fig. 1 Discharge characteristics of pyramidal neuron and recording sites of neurons in mPFC of rats
2.2 各组大鼠mPFC 中锥体神经元的自发电活动正常组大鼠锥体神经元的平均放电频率为(1.26±0.12) Hz, 多数锥体神经元(75%)呈现不规则放电, 仅有25%神经元呈现爆发式放电, 正常组大鼠ISI 的变异系数为1.20±0.07。在损毁SNc 后第3 周, 损毁组大鼠mPFC 锥体神经元的放电频率较正常组升高(P<0.05);放电形式更趋向于爆发式, 53%锥体神经元呈现不规则放电, 有47%神经元表现为爆发式放电(χ2=10.5,df=1,P=0.001), 与放电形式的改变一致, ISI 的变异系数明显高于正常组(P<0.01)。
IBO 注 射 损 毁 大 鼠MD 第2 周 后, mPFC 中 锥体神经元放电频率明显高于正常组(P<0.01), 且爆发式放电神经元百分率(64%)也较正常组明显升高, 而不规则放电的神经元百分率(36%)明显降低(χ2=30.8,df=1,P<0.001), ISI 的变异系数升高(P<0.01)。
SNc+MD 联合毁损组大鼠mPFC 锥体神经元自发放电频率与正常组比较差异无统计学意义(P>0.05);但其放电形式趋向于爆发式(χ2=19.9,df=1,P<0.001), 呈爆发式放电的神经元百分率为56%, 44%神经元呈现不规则放电, ISI的变异系数高于正常组(P<0.01)。见图2、图3和表1。
表1 各组大鼠mPFC 锥体神经元放电频率和ISI 的变异系数Tab. 1 Firing frequencies and coefficients of variation of ISI of pyramidal neurons in mPFC of rats in various groups(n=6, ±s)
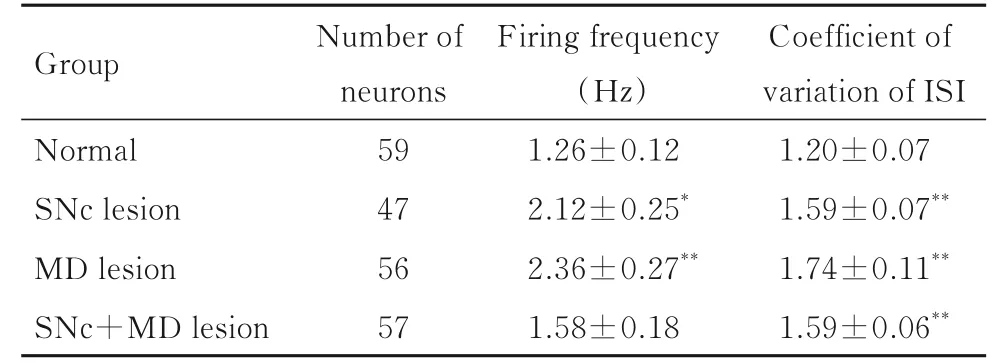
表1 各组大鼠mPFC 锥体神经元放电频率和ISI 的变异系数Tab. 1 Firing frequencies and coefficients of variation of ISI of pyramidal neurons in mPFC of rats in various groups(n=6, ±s)
*P<0.05, **P<0.01 vs normal group.
Group Normal SNc lesion MD lesion SNc+MD lesion Coefficient of variation of ISI 1.20±0.07 1.59±0.07**1.74±0.11**1.59±0.06**Number of neurons 59 47 56 57 Firing frequency(Hz)1.26±0.12 2.12±0.25*2.36±0.27**1.58±0.18
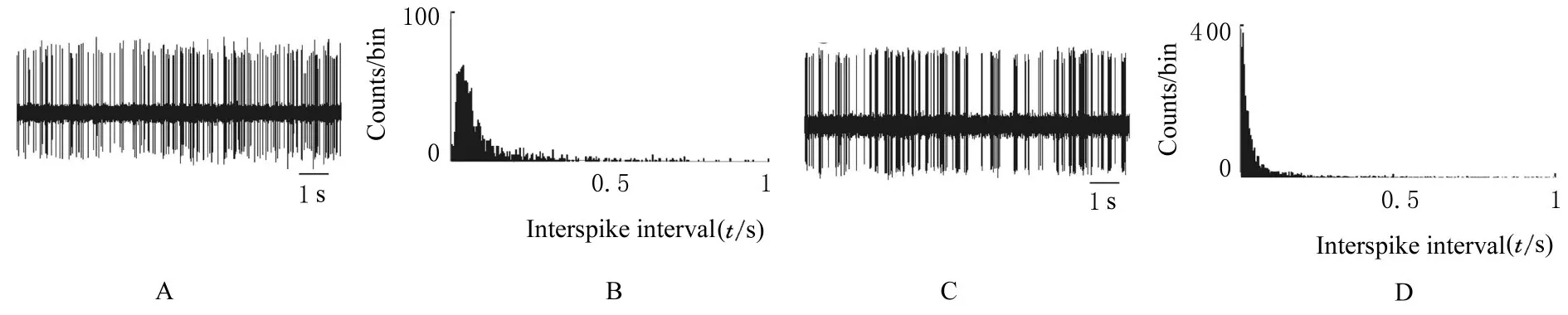
图2 mPFC 锥体神经元的放电形式和ISI 图Fig.2 Diagrams of discharge patterns and ISI of pyramidal neurons in mPFC
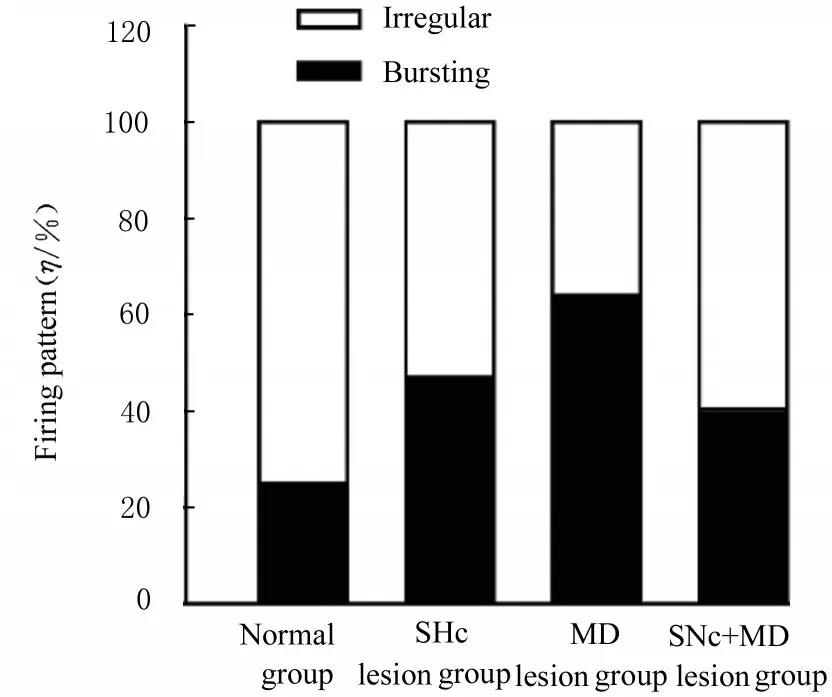
图3 各组神经元放电形式的分布Fig. 3 Distributions of firing patterns of neurons in various groups
2.3 SNc 和VTA 中多巴胺能神经元的数量和MD损毁情况在6-OHDA 注射后第3 周, 免疫组织化学染色结果表明:损毁侧SNc 中酪氨酸羟化酶免疫阳性神经元完全消失, 同侧VTA 中的酪氨酸羟化酶免疫阳性神经元数较对侧减少。见图4。

图4 SNc 和VTA 的定位图和SNc 和VTA 中酪氨酸羟化酶免疫阳性神经元形态表现Fig. 4 Location map of SNc and VTA and morphology of tyrosine hydroxylase immunopositive neurons in SNc and VTA
在电生理记录完毕后, 所有MD 损毁大鼠脑组织行冠状切片, 尼氏染色观察MD 损毁效果的结果显示:IBO 注射2 周, MD 中神经元发生固缩, 且毁损范围仅限于MD, 邻近结构很少累及, 偶尔有外侧缰核被轻微损毁的情况。凡是损伤位置有偏移, 波及到MD 周围核团, 如丘脑中央内侧、丘脑、室旁核和内侧缰核等, 电活动记录结果将剔除。只有MD 损毁位置正确的大鼠电生理记录数据才可以纳入统计学分析。典型的MD 损毁图像见图5。
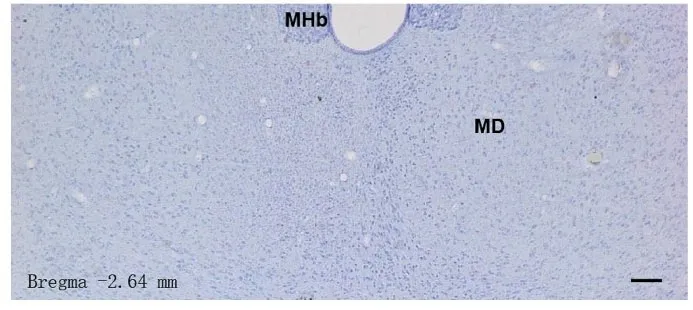
图5 MD 损毁大鼠脑组织形态表现(尼氏染色, Bar=200 μm)Fig. 5 Morphology of brain tissue of rats with MD lesion(Nissl staining, Bar=200 μm)
3 讨 论
本实验中6-OHDA 注射入SNc 中引起大鼠VTA 中相当一部分多巴胺神经元变性坏死, mPFC 主要接受来自VTA 的多巴胺能纤维投射, mPFC 锥体神经元的电生理学特征与前期报道[13]的锥体神经元特征一致, 且其神经元上表达多巴胺递质的D1和D22 种受体[14]。电或化学刺激VTA 均能够抑制PFC 中锥体神经元的自发电活动[15], 这种抑制作用是由D2受体直接介导或是通过γ-氨基丁酸能中间神经元的作用间接实现的[16-17]。研究[18]显示:采用6-OHDA 损毁内侧前脑束会导致大鼠PFC 中D1和D2这2 种受体表达均减少, 故PD大鼠mPFC 锥体神经元去抑制。神经元的放电形式是由细胞本身的特性以及来自其他脑区的神经活动共同决定的。放电形式改变与PD 症状也有一定的内在关系。FILION 等[19]研究显示:帕金森病患者中内侧苍白球神经元放电频率无明显变化, 而爆发式放电明显增加, 与病情和多巴胺能神经元减少程度成正相关关系, 并且低频爆发式放电与震颤有关。PD 状态下mPFC 神经元放电特性与PD 患者症状之间的关系尚需进一步研究。
研究[20]显示:MD 和PFC 两者之间的交互投射纤维是兴奋性的谷氨酸能纤维。MD 接受电刺激会 引 起PFC 前 扣 带 区 的 神 经 元 兴 奋[21-22]。KURODA 等[23]在一系列电镜研究中发现:MD 发出的投射纤维主要与PFC 的锥体神经元形成突触联系。近年来, 研究[24]显示:PFC 的γ-氨基丁酸能中间神经元同样受来自MD 的纤维支配。即MD不仅可以通过与PFC 中的锥体神经元形成突触连接, 直接影响锥体神经元的活动, 还可以通过中间神经元, 经过皮层的局部环路间接影响锥体神经元的兴奋性。因此, MD 损毁后锥体神经元的电活动有所增强, 表明MD 对mPFC 神经元的活动起重要调节作用。
mPFC 既接受来自MD 的纤维投射, 也接受来自VTA 的多巴胺神经元投射, 而且来自MD 和VTA 的投射纤维在PFC 中有重叠, PFC 神经元的活动可能同时受到2 种传入纤维的直接影响。VTA发出的抑制性轴突不仅支配PFC 的锥体神经元, 而且也支配MD, 因此, VTA 在一定程度上参与了MD-PFC 之 间 兴 奋 环 路 的 调 控[21]。PFC 中 的 中间神经元也参与调控MD 和VTA 对锥体神经元的传入效应, 从而在PFC、MD 和VTA 三者之间起重要的神经活动调节作用[23]。大鼠完全损毁SNc后, mPFC 锥体神经元处于去抑制状态, VTA 对MD-PFC 兴奋环路的抑制作用减弱, 在此基础上进一步损毁MD, 即PFC 中间神经元的局部抑制作用也减弱, 故锥体神经元的电活动与正常状态比较应有所增强。本研究结果显示:MD+SNc 联合损毁以后锥体神经元的放电频率与正常组比较差异无统计学意义, 爆发式放电频率亦有所增加。MD 主要发出纤维投射到前额叶皮层, 同时MD 也接受来自杏仁核、嗅觉系统、基底神经节和海马的纤维传入[25]。研 究[21]显 示:海 人 藻 酸 损 毁MD 15 d 以后, 再将6-OHDA 注入内侧前脑束, 与MD 未损毁鼠比较, VTA 对MD 诱导的PFC 神经元兴奋效应的抑制作用减弱, 说明MD 参与调控VTA 对PFC的抑制作用。也有研究[26]显示:刺激中缝中核可以抑制由MD 诱导的mPFC 激活, 而中缝中核是脑中5-HT 递质的主要来源之一。由此可见, 5-HT也参与MD 对mPFC 的调控。这种复杂的网络联系和PD 状态下多巴胺及5-HT 系统形态以及功能的变化对MD-PFC 环路的影响尚需进一步探讨。MD功能缺陷和(或) MD 对PFC 的纤维支配改变在PD 病程中的作用也有待更进一步研究。MD 损毁术治疗精神分裂症目前已在临床上广泛应用, 其理论 基 础 即 是HASSLER 等[27]提 出 的MD 损 毁 术 相当于在丘脑水平的前额叶切除术。MD 有可能成为未来临床上深部脑刺激治疗PD 的有效靶点, 故继续深入探讨mPFC 和MD 在PD 非运动症状中的作用对于PD 的临床治疗具有重要的指导意义。
