雌性大鼠吸烟对仔鼠行为学影响的研究
2022-12-10韩雨陈晨王鑫峰张海阳陈俞许德玮姚铁翼邵晓云徐绍业
韩雨陈晨王鑫峰张海阳陈俞许德玮姚铁翼邵晓云徐绍业
(1.桂林医学院基础医学院,广西 桂林 541004;2.桂林医学院科学实验中心,广西 桂林 541004;3.桂林医学院脑与认知神经重点实验室,广西 桂林 541004;4.桂林医学院临床医学院,广西 桂林 541004)
香烟烟雾中含有大量尼古丁等有害化合物,长期处于此环境中的孕期哺乳类动物可导致胎儿流产、死胎、畸形等现象,同时吸烟对神经系统的发育和功能的影响也是其众多影响之一,其中烟草对神经传递和认知功能的发育有显著的负面影响[1]。但香烟烟雾暴露对子代神经系统的影响及相关分子机理尚不清楚。坍塌反应调节蛋白(Collapsing response mediator proteins,CRMPs) 家族包含CRMP1、2、3、4、5 共5 个成员,在神经系统发育过程中高度表达,是神经系统发育中的主要磷酸化蛋白之一[2]。研究显示CRMP1 能够调节微管的组装和微丝肌动蛋白的重组,介导神经营养因子诱导神经元突起的延伸,而CRMP1 表达的减少同时影响微管的装配导致神经元轴突的稳定性下降和胞浆的运输障碍,直接导致神经元功能异常和退化,并促进神经元死亡。另外CRMP1 和CRMP2 能通过不同的信号通路调节树突棘成熟,促进/增加海马神经元突起的生长、脊柱的形成和树突棘的密度[3]。CRMP1 和CRMP4 是发育中的小鼠大脑锥体神经元树突正确定向所必需的[4]。CRMP5 对视网膜神经节细胞有着神经保护和神经再生作用[5]。死亡相关蛋白激酶1(Death-associated protein kinase 1,DAPK1)是一种钙调蛋白调节的丝氨酸/苏氨酸蛋白激酶[6],在机体中广泛参与由多种信号介导的细胞死亡过程[7],同时在大脑中高度表达[8]。研究表明,它与神经系统疾病相关的多种神经元损伤有关[9]。DAPK1 作为细胞凋亡正调控因子,可通过上调p53 基因的表达而促使细胞进入Ⅰ型细胞凋亡程序,也可与非p53 依赖的经典凋亡途径发生关联[10]。另有研究发现,DAPK1 亦可诱导一种不需要半胱天冬酶参与的Ⅱ型细胞死亡或称自体吞噬性细胞死亡[9]。
实验前期研究初步证明了香烟烟雾暴露对胎儿多器官发育均有一定影响[11]。本实验进一步研究亲代雌鼠香烟烟雾暴露对子代大鼠行为学的影响,通过旷场实验、转棒疲劳实验和Morris 水迷宫实验从整体水平检测大鼠的焦虑、运动耐受能力、以及对空间位置和方向的学习记忆能力的影响。同时,在分子水平通过Western Blot 检测亲代雌鼠香烟烟雾暴露对子代大鼠大脑皮层组织中DAPK1 和CRMP1 蛋白表达的影响。旨在阐明亲代雌鼠香烟烟雾暴露对子代神经系统行为学的影响及其机制,为预防和治疗香烟烟雾暴露引起的神经系统损伤提供分子靶点。
1 材料与方法
1.1 材料
1.1.1 实验动物
选用18 只7~8 周龄SPF 级SD 雌性大鼠,体重200~220 g,雄性9 只,体重220~240 g,均购于湖南斯莱克景达实验动物有限公司【SCXK(湘)2019-0004】。大鼠饲养提供标准饲料,饮用高压灭菌水,光照和黑暗各12 h 轮流交替,湿度保持在60%左右,温度控制在22℃左右,饲养于桂林医学院实验动物中心【SYXK(桂)2020-0005】。动物实验经桂林医学院实验动物伦理委员会审查,符合动物福利伦理相关要求(GLMC-IACUC-2021006)。
1.1.2 主要试剂与仪器
广西某品牌香烟(20 支/包);RIPA 裂解液(Beyotime Biotechnology,上海,中国);PMSF 苯甲基磺酰氟(Solarbio Life Sciences,北京,中国);磷酸酶抑制剂(MCE,Monmouth Junction,NJ,美国);兔源CRMP1 多克隆抗体(Proteintech Rosemont,IL,美国);兔源 DAPK1 多克隆抗体(Proteintech Rosemont,IL,美国);鼠源GAPDH 单克隆抗体(Proteintech Rosemont,IL,美国);Protein marker(Thermo Waltham,MA,美国);BCA 试剂盒(Beyotime Biotechnology,上海,中国);ECL 发光试剂盒(Thermo Waltham,MA,美国)。
自制半封闭式烟熏染毒箱(150 cm × 60 cm ×60 cm);旷场装置(型号XR-XZ301)、转棒疲劳仪(型号XR1514-RZPM)、Morris 水迷宫(型号XRXM101),均由上海欣软信息科技有限公司生产。
1.2 方法
1.2.1 烟熏大鼠模型的建立及分组
参照徐绍业等[12]使用的自制半封闭式烟熏染毒箱(150 cm × 60 cm × 60 cm),以饲养箱为骨架,分为上下两层,箱两边预留1 cm × 1 cm 通气孔各3个。参照陈茜等[13]烟熏模型方法并改进,在箱内上下层各设置一个燃烟点,每个燃烟点每次连续自燃香烟10 支,烟熏维持约60 min,每天2 次(9:00,15:00)。将亲代雌鼠随机分为烟熏组和对照组各9只,烟熏组雌鼠置于半封闭式烟熏染毒箱,对照组雌鼠于同环境、同饲料喂养,但不给予烟雾吸入;用于合笼交配的雄鼠9 只不作处理,正常饲养。烟熏1 个月后,将烟熏组和对照组雌鼠分别以2∶1 比例与雄鼠合笼(每晚19:00),次日晨分笼,反复数日,直至确定雌鼠怀孕则停止合笼。待孕鼠分娩出子代大鼠后则停止烟熏处理,烟熏持续两个月左右。随后在烟熏组和对照组中不同亲代大鼠繁育的后代中筛选出体重相当的3 窝仔鼠(每窝分别为11只、12 只、13 只),待成年(约4 周龄)进行旷场实验、转棒疲劳实验、Morris 水迷宫实验。同时,收集不同发育阶段(0、7、21 d 和4 周)烟熏组和对照组子代大鼠的大脑皮层脑组织,Western Blot 检测CRMP1 和DAPK1 蛋白的表达情况。实验使用戊巴比妥钠过量麻醉对大鼠进行安乐死处理。
1.2.2 旷场实验
旷场实验检测大鼠自发探索能力、活动能力和焦虑情绪。SD 大鼠选用旷场装置(50 cm × 50 cm ×50 cm)为底部和侧壁均为黑色有机板的自发活动箱,应用自发活动视频分析系统将自发活动箱平均分为25 格,设定周边16 格为外周区,中间9 格为中央区。实验前将待测大鼠置于旷场装置自由停留5 min 进行环境适应,每天1 次,连续2 d。正式实验时间尽量与环境适应时间一致。正式实验即将待测大鼠放入旷场装置底板中心,随后使其自由活动300 s,记录子代大鼠全程运动总距离、平均运动速度,以及进入中央区和外周区的次数、在中央区和外周区的运动距离和停留时间,子代大鼠运动路径图见图1A。
1.2.3 转棒疲劳实验
转棒疲劳实验检测大鼠的运动耐受能力。预先将待测大鼠置于转棒疲劳仪(转棒直径60 mm,跑道宽度80 mm)进行训练,每次以40 r/min 转速持续5 min,每只大鼠每天训练3 次,期间间隔15 min,共训练2 d。2 d 后进行正式测试,每只鼠测试3 次,方法与训练期相同,记录大鼠在转棒疲劳仪上的持续运动时间及运动距离,取平均值。
1.2.4 Morris 水迷宫实验
Morris 水迷宫实验检测大鼠对位置和方向的空间学习记忆能力。水迷宫水池直160 cm,将水池均分为4 个象限,设置隐匿逃生平台(直径8 cm)于其中1 个象限的中央,根据4 个象限分别以A、B、C、D标记为大鼠4 个入水点。注水深度50 cm,注水后使平台没入水面以下1 cm,实验时向水池内注入黑色墨水使之与池水混匀。测试期间迷宫外部各参照物保持不变,水温维持(25 ± 2)℃。首先进行获得性训练(即定位航行实验),随机选择一个入水点将大鼠面向池壁轻轻放入水中,若2 min 内四肢登上隐匿平台,则使其在平台上停留15 s;若2 min 内尚未找到平台,则人为引导其寻找平台并停留15 s,每只大鼠每天训练4 次,分别从4 个不同的入水点入水,每次训练间隔15~20 min,连续5 d,记录大鼠的逃避潜伏期、到达平台的游泳路程、以及平均游泳速度。第6 天进行空间探索实验,即在定位航行实验基础上撤除逃生平台,将待测大鼠从原平台象限(目标象限)的对侧入水,计时2 min,记录大鼠到达目标象限的时间、进入目标象限的次数、在目标象限停留时间、平均游泳速度、总游泳路程。
1.2.5 Western Blot
取烟熏组和对照组子代大鼠不同生长阶段的大脑皮层组织,用RIPA 裂解液(临用时添加蛋白酶抑制剂PMSF 和磷酸酶抑制剂)完全裂解脑组织,提取大脑皮层组织总蛋白,采用BCA 试剂盒法测定蛋白浓度。将各组蛋白样品按一定顺序加入上样孔,进行电泳、转膜、5%脱脂奶粉封闭2 h 后,选择目的抗体在4℃冰箱旋转孵育过夜,次日用1 × TBST 洗膜后进行室温二抗孵育1 h,再次洗膜后应用ECL化学发光液对目的蛋白条带进行曝光显影。
1.3 统计学分析
2 结果
2.1 亲代雌鼠香烟烟雾暴露增强了子代大鼠的焦虑感
旷场实验结果显示,烟熏组子代大鼠在总区域较对照组的运动总距离(P<0.05,图1B)和平均运动速度(P<0.05,图1C)明显增加;子代大鼠在中央区的运动情况显示,烟熏组子代大鼠进入中央区的次数(P<0.05,图1D)、在中央区的运动距离(P<0.05,图1E)、以及在中央区的停留时间(P<0.05,图1F)明显增加。同时,子代大鼠在外周区的运动情况表现为,烟熏组较对照组子代大鼠进入外周区的次数(P<0.05,图1G)和在外周区运动的距离(P<0.05,图1H)明显增加,但烟熏组子代大鼠在外周区停留时间有所减少(P<0.05,图1I)。上述结果推测,亲代雌鼠香烟烟雾暴露可能促使子代大鼠运动亢奋激发其探索新知的能力,同时也增加了子代大鼠的焦虑情绪(见图1)。
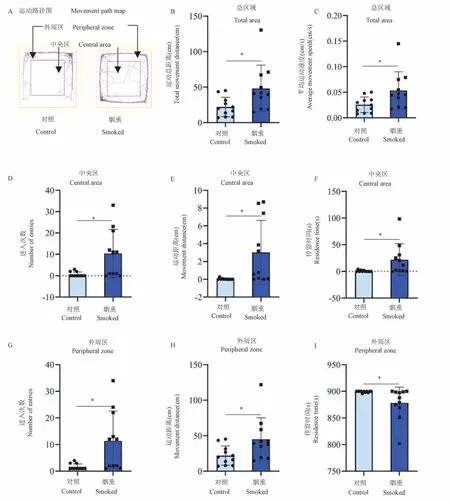
图1 烟熏组和对照组子代大鼠的旷场实验结果(n=11)Note.A.Movement path map of offspring rats on the open field device.B.The total movement distance of offspring rats in the total area.C.Average movement speed of offspring rats in the total area.D.The number of offspring rats entering the central region.E.Movement distance of offspring rats in the central region.F.Residence time of offspring rats in central region.G.The number of offspring rats entering the peripheral area.H.Movement distance of offspring rats in peripheral area.I.Residence time of offspring rats in peripheral area.Compared with the control group,∗P <0.05.Figure 1 Open field test results of offspring rats in the smoked group and the control group(n=11)
2.2 亲代雌鼠香烟烟雾暴露降低子代大鼠的运动耐受能力
转棒疲劳实验结果显示,烟熏组子代大鼠在转棒疲劳仪上的持续运动时间(P<0.001,图2A)和运动的距离(P<0.001,图2B)较对照组子代大鼠显著减少。表明亲代雌鼠香烟烟雾暴露会导致子代大鼠运动耐受能力显著下降(见图2)。
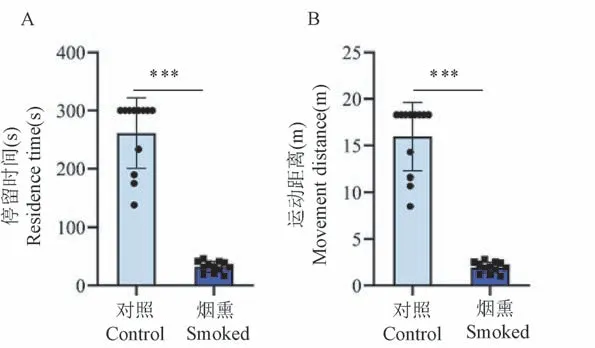
图2 烟熏组和对照组子代大鼠转棒疲劳实验结果(n=12)Note.A.Exercise time of offspring rats.B.Movement distance of offspring rats.Compared with the control group,∗∗∗P <0.001.Figure 2 Results of rod-rotating fatigue test of offspring rats in the smoked group and control group(n=12)
2.3 亲代雌鼠香烟烟雾暴露对子代大鼠空间学习和记忆能力的影响
Morris 水迷宫实验结果显示,在获得性训练的定位航行实验中,烟熏组子代大鼠的逃避潜伏期(P>0.05,图3A)和抵达平台的运动路程(P>0.05,图3B)与对照组无显著性差异。随着训练天数的增加,子代大鼠逃避潜伏期和抵达平台的运动路程均逐渐缩小,表明亲代雌鼠香烟烟雾暴露并没有影响子代大鼠的学习能力,但烟熏组子代大鼠的游泳速度较对照组明显下降(P<0.001,图3C)。
在撤除平台后进行的空间探索实验中,烟熏组子代大鼠较对照组在目标象限停留的时间占比下降(P<0.05,图3D),进入目标象限的次数明显减少(P<0.05,图3E)。整个游泳过程中烟熏组子代大鼠的平均游泳速度明显降低(P<0.01,图3F),总游泳路程也较对照组显著减少(P<0.01,图1G)。这表明亲代雌鼠香烟烟雾暴露会导致其子代大鼠的空间记忆能力减退,且运动能力也明显减弱(见图3)。

图3 烟熏组和对照组子代大鼠Morris 水迷宫实验结果(n=13)Note.The positioning navigation experiment.A.Escape latency of offspring rats.B.The swimming distance of offspring rats reaching the platform.C.Average swimming speed of offspring rats.The space exploration experiment.D.Proportion of residence time of offspring rats in target quadrant.E.The number of offspring rats entering the target quadrant.F.Average swimming speed of offspring rats.G.Total swimming distance of offspring rats.Compared with the control group,∗P <0.05,∗∗P <0.01,∗∗∗P <0.001.(The same in the following figures)Figure 3 Morris water maze test results of offspring rats in the smoking group and control group(n=13)
2.4 不同发育阶段子代大鼠大脑皮层中CRMP1、DAPK1 的蛋白表达情况
Western Blot 结果显示,烟熏组子代CRMP1 蛋白在0 d(P<0.001,图4A,4B)、7 d(P<0.05,图4C,4D)、21 d(P<0.05,图4E,4F)的表达水平均显著低于对照组。而在4 周(P>0.05,图4G,4H)时,两组间CRMP1 蛋白表达无明显差异。DAPK1蛋白的表达水平在各个检测阶段0 d(P<0.001,图4A,4B)、7 d(P<0.01,图4C,4D)、21 d(P<0.05,图4E、4F)、4 周(P<0.001,图4G,4H)均具有显著性差异,尤其是在发育早期。表明亲代雌鼠香烟烟雾暴露可影响子代大鼠大脑皮层中CRMP1 和DAPK1 的表达(见图4)。

图4 Western Blot 检测烟熏组和对照组子代大鼠不同发育阶段大脑皮层CRMP1、DAPK1 蛋白表达情况Note.A,B.Progeny rats were born 0 d.C,D.Progeny rats were born for 7 d.E,F.Progeny rats were born for 21 d.G,H.The progeny rats were born 4 weeks.Figure 4 Western Blot analysis of CRMP1 and DAPK1 protein expression in cerebral cortex of offspring rats at different developmental stages in the smoking group and the control group
3 讨论
众所周知,中国是一个烟草生产和消费大国。虽然中国女性有吸烟习惯者较欧美女性少,但是中国男性吸烟者众多,由此导致的女性被动吸入二手烟的几率亦随之增加。香烟烟雾中含有的大量尼古丁等有害化合物被母体吸收后将导致孕妇妊娠期并发症[14],也可能通过胎盘屏障进入胎儿体内影响胎儿大脑发育[15],怀孕期间长期处于此环境可导致胎盘早剥、流产、死产、早产(<37 周妊娠)和低出生体重(<2500 g)[16]等,并且会导致胎儿神经发育不良及后代多指(趾)畸形等有害影响。
到目前为止,孕妇香烟烟雾暴露对后代大脑神经系统发育的影响及相关分子机理尚不清楚。为了探究香烟烟雾暴露对其子代神经系统发育的影响,本研究通过香烟烟雾暴露对亲代雌鼠进行烟熏处理,并通过动物行为学检测子代大鼠在情绪、运动能力及学习记忆等方面的变化。烟熏组和对照组亲代雌鼠分别娩出53 只和72 只子代大鼠,其中对部分成年期子代大鼠(4 周)进行动物行为学检测,部分发育期(0 d,7 d,21 d,4 周)子代大鼠进行Western Blot 检测。
旷场实验结果显示,烟熏组子代大鼠进入旷场中央区的频率更高,且其在总区域的运动总路程和平均运动速度均较对照组明显增加,这可能与亲代雌鼠香烟烟雾暴露导致其子代大鼠过度活跃有关。有研究比较了在亲代雄鼠和雌鼠中尼古丁暴露对其子代大鼠的行为学影响,结果发现尼古丁暴露的雌鼠其子代大鼠表现为过度活跃和冲动性行为增加[17]。Liu 等[18]曾报道亲代雌性接触尼古丁可能会导致子代大鼠产生神经生物学障碍,如多动症、学习和记忆问题等。同时,本研究结果还显示,烟熏组子代大鼠在外周区和中央区之间的穿越次数亦较对照组明显增多,由此表明,亲代雌鼠香烟烟雾暴露后将导致其子代更强烈的焦虑感,这与Zhou等[19]的研究结果一致,即母体尼古丁暴露的子代在幼年期表现出身体和神经行为发育迟缓,增加焦虑水平。
二手烟暴露将会引起哮喘、呼吸道感染、肺生长和功能受损、运动耐受性下降、认知障碍、行为问题等多种不良健康风险。多项研究表明,香烟烟雾暴露可减缓小鼠的生长发育,并使小鼠的运动能力明显减弱[20-21]。但目前关于亲代香烟烟雾暴露后对子代运动能力影响的报道甚少。本研究转棒疲劳实验结果显示,亲代烟熏组的子代大鼠运动耐受能力显著下降,与对照组存在极显著性差异(P<0.001)。由此可见,亲代香烟烟雾暴露也可影响其子代的运动耐受能力。
Morris 水迷宫主要用于检测实验动物的学习和记忆能力。本研究在Morris 水迷宫前5 d 的获得性训练中表现为,子代大鼠的逃避潜伏期和寻找平台的总路程在烟熏组和对照组之间无明显的差异;随着训练天数的延长,二者潜伏时间和寻找路程均随之缩短,且二者几乎趋于一致。上述研究表明,亲代大鼠香烟烟雾暴露并不影响子代大鼠的学习能力。这与Alzoubi 等[22]在水烟暴露对大鼠学习能力影响的研究结果相一致。但是,烟熏组子代大鼠的平均游泳速度却较对照组明显降低,这进一步说明亲代雌鼠香烟烟雾暴露会对子代大鼠的运动能力产生不良影响,这与转棒疲劳实验结果一致。在后续的空间探索实验中,烟熏组子代大鼠在目标象限停留的时间以及进入目标象限的次数都明显减少,由此表明,亲代雌鼠香烟烟雾暴露后会影响子代大鼠的空间记忆能力。此前一项关于水烟环境对大鼠学习记忆影响的研究发现,接触水烟环境的大鼠会导致短期和长期记忆受损[22]。而且,据报道,围产期尼古丁暴露会导致子代小鼠的注意力和记忆力的显著缺陷,有趣的是,这一缺陷具有性别依赖性,雄性后代而非雌性后代更明显[23]。同时,在空间探索实验中,亲代烟熏组雌鼠的子代大鼠的平均游泳速度以及总游泳路程也明显减少,同样表明,亲代雌鼠香烟烟雾暴露会减弱子代大鼠的运动能力。
有研究表明,香烟烟雾暴露会诱发脑部炎症和氧化应激[24],在子代小鼠大脑皮层产生炎症反应,如坏死和细胞质空泡化,还观察到大量的凋亡细胞和死亡的神经元,对大脑的功能产生负面影响,从而导致子代小鼠记忆功能的异常[25]。为了探究亲代香烟烟雾暴露对子代生长发育及凋亡的影响,本研究分别收集子代大鼠出生后不同阶段(0 d、7 d、21 d、4 周)的大脑皮层组织,并选择与发育和凋亡相关的靶分子CRMP1 和DAPK1 进行蛋白水平检测,从结果可以看出,烟熏组子代大鼠较正常组子代大鼠脑组织中CRMP1 和DAPK1 蛋白表达水平均显著降低。由于CRMP1 具有调节树突棘成熟、促进/增加海马神经元突起的生长、脊柱的形成以及增加树突棘密度的功能[3],推测亲代大鼠香烟烟雾暴露导致CRMP1 显著下降,可能造成树突棘的成熟障碍,并导致神经元突起生长受限,从而影响了脑细胞的正常生理功能;同时,DAPK1 作为细胞凋亡正调控因子,促进细胞凋亡[6],广泛参与由多种信号介导的细胞死亡过程[7],而亲代大鼠香烟烟雾暴露致使子代大鼠脑组织中DAPK1 的表达降低,可能打破了脑组织中正常细胞凋亡和发育的平衡。但亲代雌鼠香烟烟雾暴露后导致其子代大鼠产生的一系列行为学改变(焦虑、运动耐受能力减退、空间记忆能力减退等)是否与CRMP1、DAPK1相关联,该靶分子调节子代大鼠行为学改变的机制如何则仍需进一步探讨。
本研究结果表明亲代雌鼠香烟烟雾暴露可增强子代大鼠的焦虑感、降低其运动耐受能力,并导致子代大鼠的记忆力减退,这可能与CRMP1 和DAPK1 的调节有关。于此,我们认为有必要以多种形式开展反吸烟宣传活动,提高公众对二手烟危害的认识以及保护女性及儿童们免受其害,并从分子生物学角度进一步探索香烟烟雾暴露导致神经系统损害的治疗靶点。
