虾类活体贮运中应激诱发因素及其影响研究进展
2022-12-10罗小莹李雪红李述刚吴文锦丁安子熊光权
罗小莹,李雪红,李述刚,褚 上,石 柳,吴文锦,丁安子,熊光权,汪 兰,
(1.湖北工业大学生物工程与食品学院,湖北武汉 430064;2.湖北省农业科学院农产品加工与核农技术研究所,湖北武汉 430064;3.合肥工业大学食品与生物工程学院,安徽合肥 230000)
虾类为甲壳动物,隶属于节肢动物门、甲壳纲、十足目,是水产养殖中重要的经济物种,因其肉质鲜美,营养成分丰富,深受人们喜爱[1-3]。据统计资料显示[4],2020年我国虾类养殖总产量达4969475 t,与2019年同比增长13.04%,居世界第一位,并持续保持增长势头。相比于陆生动物来说,虾类水产品的生长具有明显的季节性和地域性,并且虾肉体内结缔组织少、肉质柔软、水分含量高、酶活力强,死后极易腐败变质[5],所以我国虾类水产品多以鲜销为主。但是虾在活体贮运过程中对各种应激源反应比较敏感,容易受到温度、溶氧量、高度拥挤、振动等环境因子的影响,诱发机体产生应激反应[6],对其机体功能及肌肉品质产生不利影响,导致其市场价值和竞争力大大降低。因此,研究虾类活体贮运中应激反应的诱发因素,贮运过程中虾体内综合生理反应情况及运输应激对肌肉品质的影响,对虾类活体流通和销售具有重要意义。
本文结合国内外关于虾类甲壳动物应激的研究成果,对虾在运输过程中受环境因素变化引发的机体应激问题进行综述,并列举了有关虾类活体贮运的对策与建议,以期为提高虾类运输的存活率提供指导性建议,同时也为揭示虾类活体贮运过程中的综合生理反应和肉质变化机制提供理论依据。
1 影响虾类活体贮运的环境因素
虾在活体贮运过程中容易受到外部环境胁迫,产生应激反应,使其机体发生不同程度的损伤[7]。环境胁迫因素大致可分为以下几个方面。
1.1 温度胁迫
温度是影响水生动物生长、发育的关键因素之一[8]。水温的变化会给生物带来不同程度的压力,并对它们的健康造成不利影响[9]。Guo等[10]发现高温胁迫会影响克氏原螯虾肠道器官的生理动态平衡,对其抗氧化状态和免疫功能产生不利影响。Xu等[11]研究发现南美白对虾在低温和空气暴露胁迫下,血细胞数量减少,形态变化,严重时会诱导血细胞凋亡,导致免疫系统恶化。因此,在虾的活体运过程中保持适宜的运输温度显得尤为重要。Coyle等[12]研究发现罗氏沼虾在运输水温为21 ℃时平均存活率为97%,而26 ℃时仅为24%。陈水春等[13]研究发现在长途运输过程中处理温度为12 ℃时龙虾存活率最高,达95.42%。但不同种类乃至不同规格的虾对温度的适应性不同,因此在运输过程中应针对性的做出调整。
1.2 低氧胁迫
据报道,水生动物暴露在空气中时会破坏体内氧气消耗,导致乳酸浓度增加发生无氧代谢相关的酸中毒[14],暴露在空气中还会增加甲壳类高血糖激素、血淋巴葡萄糖水平、降低颗粒细胞比例[15],增加菌血症和延长凝血时间等[16]。Broughton等[17]对水中缺氧小龙虾的行为、生理生化反应进行研究发现,小龙虾对低氧(3.5 kPa)有较高的耐受性,但重度低氧(1.7 kPa)时,会导致其心率加快,血淋巴酸碱状态紊乱,影响正常的酶功能和代谢调节。Zhou等[18]研究发现低氧(1 mg·L-1)会显著降低凡纳滨对虾的血细胞总数,降低对虾的免疫防御能力。由于不同生物对氧的需求量不同,因此低氧浓度的具体数值也有差异。较为合理的缺氧临界值应考虑生物的针对性。
1.3 氨氮胁迫
氨氮是水产动物活运中重要的环境胁迫因子,主要来自水产动物自身的代谢物质[19]。氨通常以电离状态(N H+4)和非电离状态(N H3)存在于水中[20]。当水体中氨的水平超过虾的耐受极限时,会对虾的生长、渗透调节、免疫、代谢和生存等产生负面影响[21]。Liu等[22]研究氨胁迫对南美白对虾血细胞的影响,结果表明氨暴露后对虾血细胞总数显著下降,细胞凋亡率升高,影响了对虾的免疫功能,并增加对虾对病原性感染的几率。Wang等[23]通过分析氨胁迫下克氏原螯虾血淋巴、鳃和肝胰腺的转录水平也发现,氨胁迫会显著改变克氏原螯虾的嘌呤代谢途径,破坏氧化-抗氧化平衡,并导致细胞凋亡,影响其正常的生命活动。因此,在运输过程中应及时对水质进行净化处理,防止氨氮物质大量积累,减少虾在运输途中的死亡率。
1.4 酸碱胁迫
pH是环境胁迫中最敏感的因素之一,虾类对pH胁迫的反应是一个复杂的过程,通常涉及免疫系统、抗氧化系统和肠道微生物区系的改变[24]。Yu等[25]将南美白对虾置于高pH9.5或低pH6.5的环境中时,对虾的抗氧化能力显著降低,免疫相关基因表达下调,肠组织变形。Cheng等[26]研究也发现pH的变化会导致罗氏沼虾的血细胞计数和酚氧化酶活性降低,并增加其对肠球菌的易感性。此外,pH的急性变化也会使得虾体内活性氧(ROS)增加,氧化损伤加剧,抗氧化调节能力失衡[27]。因此,在虾的活体运输过程中应实时监测水体的pH,并将其维持在适宜的范围,避免虾体发生酸中毒或碱中毒。
1.5 拥挤胁迫
虾类甲壳动物在运输期间的运输密度(生物量)是影响应激水平、损伤发生和存活率的重要因素[28]。高密度的运输量会使虾体产生拥挤胁迫,改变水生动物的行为和生理生化状态,使免疫力下降[29],还会导致含氮物质的积累和溶解氧的消耗,增加死亡率[30]。于郝男[31]在不同密度胁迫下对罗氏沼虾和凡纳滨对虾行为及能量分配模式的研究发现,随着密度的增大,罗氏沼虾和凡纳滨对虾的活动和攻击的频次明显增加,耗氧率和排氨率上升,能量消耗增多,免疫力下降。Zhang等[32]采用蛋白质组学的方法,分析拥挤和非拥挤条件下中国对虾肌肉蛋白质的差异表达,结果表明在拥挤应激后对虾肌肉代谢相关蛋白在翻译和转录水平上明显上调,这些蛋白质表达的变化可能会导致对虾肉质的下降。因此,在运输过程中应根据虾的种类、规格及装载系统对运输密度做出调整,确定生物量、存活率和成本之间的最佳交叉点。
1.6 机械损伤胁迫
水产品在装卸运输过程中容易受到搬运、振动、噪音等机械损伤胁迫而产生应激反应,一般会导致水产品的免疫能力下降、体内抗氧化平衡失衡、肌肉品质降低等[33],严重影响水产品的运输质量。另外在运输过程中水产动物个体间也会互相拥挤、互相攻击,导致机体的储能物质被大量消耗,生理代谢平衡被破坏。陈梓聪[34]以斑节对虾为研究对象,分析其力竭运动后的抗氧化能力和呼吸代谢恢复规律,发现力竭运动后对虾机体酶促抗应激体系被激活,呼吸代谢活动增强。Vermeersch等[35]研究发现在捕获、分拣和运输过程中对褐虾的翻滚、堆叠会导致虾的重要器官损伤或丢失,并且受伤的虾在2 h内表现出很高的死亡率(32.8%~69.8%)。因此,在虾的活体运输过程中需要改进装载和卸载的处理方式,优化运输设备,以排除机械损伤胁迫对虾体造成危害。综上所述,虾类甲壳动物在环境胁迫下生理动态平衡和免疫系统都会发生不同程度的适应性改变。除上述例举的影响外,许多学者还发现环境胁迫会使虾类甲壳动物的组织器官和抗氧化系统发生改变,主要表现为虾的肝胰腺结构受损及相关抗氧化酶活性变化,氧化损伤增加。如Tania等[36]研究发现,凡纳滨对虾在缺氧条件下肝胰腺中超氧化物自由基的产生及硫代巴比妥酸(TBARS)的含量要高于鳃或肌肉组织,这可能与肝胰腺中较高的代谢率或较高的脂质含量有关。Liu等[37]研究发现,空气暴露会对南美白对虾造成氧化损伤,使其抗氧化酶活性发生改变,但重新浸泡后这种损伤是可以逆转的。以上研究表明,环境胁迫能够对虾的组织器官及抗氧化能力造成一定的损伤,通常情况下这种氧化损伤是可以逆转的,但是当压力增强或持续时,就会造成不可逆的伤害,最终导致虾体死亡(表1)。
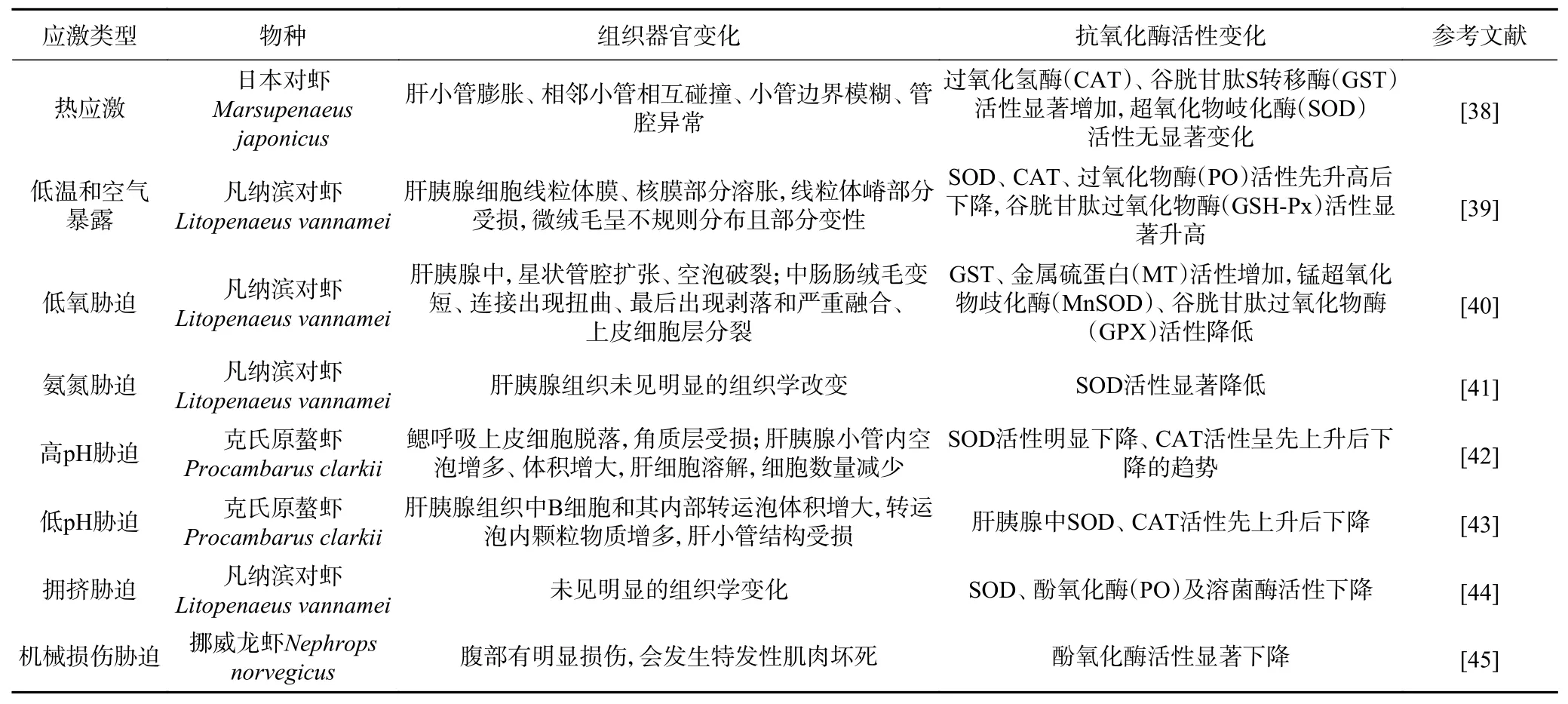
表1 环境胁迫对虾类甲壳动物组织器官和抗氧化酶活性的影响Table 1 Effects of environmental stress on the tissues, organs and antioxidant enzyme activities of shrimp crustaceans
2 运输应激对虾类综合生理反应及肌肉品质的影响
2.1 运输应激对虾类综合生理反应的影响
2.1.1 氧化应激 虾在环境胁迫下氧化状态会发生改变,当氧化和抗氧化系统之间的平衡被破坏时会增加活性氧(ROS)的产生并引起氧化应激[46-48]。活性氧(ROS)是一类高活性分子,包括超氧阴离子自由基过氧化氢(H2O2)、羟自由基(OH·)和单线态氧(1O2)等,过量的ROS产生和积累会损害脂质、蛋白质和DNA,导致氧化损伤[49]。抗氧化防御系统在清除过量ROS并维持ROS的平衡方面起着重要作用,主要由超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、谷胱甘肽过氧化物酶(GPx)和谷胱甘肽S-转移酶(GST)等抗氧化酶和谷胱甘肽(GSH)、金属硫蛋白(MT)、抗坏血酸(VC)、维生素A(视黄醇)和维生素E(α-生育酚)等非酶类抗氧化剂组成[50-51]。Wang等[52]研究发现,南美白对虾在急性pH应激下MnSOD mRNA、CAT mRNA、GPx mRNA的表达水平上调,表明抗氧化防御系统对pH应激做出了反应。
此外丙二醛(MDA)的含量也是氧化应激的指标,因为它是自由基与脂质反应产生的终产物,可以间接反映体内活性氧的积累情况[53]。Li等[54]研究发现,在低氧条件下南美白对虾肝胰腺和血淋巴中丙二醛(MDA)含量显著上升,说明低氧可诱导肝胰腺和血淋巴组织的氧化应激和脂质过氧化。一般情况下,虾都可以利用自身的抗氧化系统对生成的自由基进行及时清除,只有应激时间延长或者应激强度加大时才会使抗氧化系统的功能受到抑制,从而产生氧化损伤。
2.1.2 内质网应激 内质网(Endoplasmic reticulum,ER)是蛋白质折叠和运输的特殊细胞器,对细胞内稳态和细胞外刺激的变化高度敏感[55]。当虾类水产动物受到环境胁迫时,会扰乱内质网稳态,引起内质网应激[41],在这种情况下,错误折叠和未折叠的蛋白质会在内质网中积累,激活未折叠蛋白反应(Unfoldedprotein response,UPR)。UPR是一组适应性信号通路的集合,这些信号通路由位于内质网中的三个信号转导器受到刺激而启动。UPR包括三个经典的信号通路:肌醇需要蛋白1α-X盒结合蛋白1(inositolrequiring protein 1α-X-box binding protein 1,IRE1α-XBP1)通路、活化转录因子6α(activating transcription factor 6α,ATF6α)通路和蛋白激酶RNA样内质网(ER)激酶(PERK)-eIF2α[protein kinase RNA-like endoplasmic reticulum (ER) kinase-eIF2α,PERKeIF2α]通路。在正常生理条件下,BiP会结合并抑制IRE1α、ATF6α和PERK三种激酶,而当内质网应激发生时,错误折叠的蛋白就会将Bip与三种激酶解离并激活相应的信号转导通路(图1)。 首先,激活的PERK激酶可以使eIF2α磷酸化,并选择性地增加ATF4的翻译,以调节适应性基因的表达;其次激活的ATF6α激酶从内质网转位到高尔基体后被驻留的蛋白酶S1P和S2P水解产生有活性的ATF6α,活性ATF6α会转位到细胞核并诱导靶基因的转录;同时,IRE1α可以被二聚化和磷酸化激活,导致XBP1 mRNA的剪接,从而产生活性转录因子XBP1s,XBP1s转位到细胞核后调控一些与蛋白降解相关酶的表达[56]。通过这三个信号通路的共同作用可以增加内质网中蛋白质的折叠能力、降低蛋白质折叠负荷、减少蛋白质合成,从而缓解内质网应激[57]。
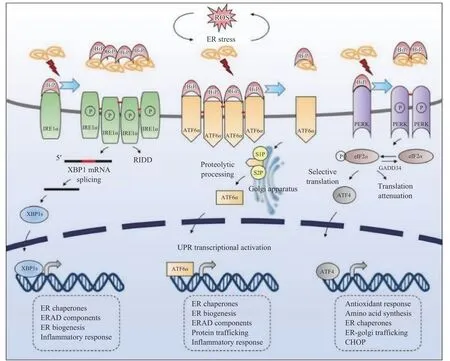
图1 内质网应激下的UPR途径[56]Fig.1 UPR pathway under ER stress[56]
内质网除了是蛋白质合成、折叠与运输的主要部位外,也是协调葡萄糖代谢和脂质代谢反应的重要细胞器[58]。严重或持续的内质网应激会导致细胞内葡萄糖代谢异常。Wang等[59]对急性冷应激后凡纳滨对虾差异表达基因(DEGs)的KEGG通路分析发现,22个DEGs参与了内质网途径中的蛋白质加工,其中内质网伴侣蛋白Bip-like(BIP)表达显著上调。BIP作为葡萄糖调节蛋白,在UPR过程中起着重要作用[60]。冷应激下内质网中BIP的表达变化,说明葡萄糖的合成或分解途径在UPR过程中受到转录调控。此外,UPR的三个通路也与细胞脂肪合成调节有关。Wang等[61]研究发现,对虾在氨氮胁迫下IRE1、XBP1、eIF2α和ATF4基因表达上调,说明UPR信号转导通路被激活,机体发生内质网应激,同时在内质网应激水平较高的实验组中脂质合成相关酶活性显著升高,肝胰腺脂质积聚,说明高水平的内质网应激会导致肝胰腺脂质合成和分解异常。UPR是在应激环境下内质网引发的一种保护性或适应性策略,目的是恢复内质网的稳态,但如果应激时间延长,就会启动细胞凋亡信号,最终导致细胞死亡[62]。因此在运输过程中要严格控制虾类水产动物周围的环境条件,避免应激造成的不利影响。
2.1.3 能量代谢 生物在不同的应激源刺激下一般会增加机体能量消耗,以维持体内的动态平衡。血糖水平的变化是虾类水产动物对不同应激源的主要典型反应[63-64]。在虾对应激的适应性反应中,葡萄糖水平的增加主要是由高血糖激素(CHH)介导的,当受到应激源刺激时,CHH通过脑啡肽能神经元或5-羟色胺能神经元的信号立即从窦腺释放到血淋巴中,释放的CHH会刺激肌肉和肝胰腺中糖原的水解,从而增加血淋巴中的葡萄糖浓度[65]。形成的葡萄糖随后在血淋巴中积累,导致高血糖,从而满足应激后的能量需求。这种葡萄糖水平的变化可以直接反映虾类水产动物利用糖原进行了碳水化合物代谢。
在高水平应激条件下,虾类水产动物也会进行脂质代谢来为机体提供更多的能量,其中甘油三酯(TG)和总胆固醇(T-CHO)含量的变化可以反映脂质代谢水平[66]。Li等[67]研究凡纳滨对虾在不同浓度亚硝酸盐应激后肝胰腺组织中甘油三酯(TG)、总胆固醇(T-CHO)水平和脂肪酸合成酶(FAS)活性的变化,发现亚硝酸盐暴露后,各组的甘油三酯(TG)和总胆固醇(T-CHO)水平以及脂肪酸合成酶(FAS)活性均呈下降趋势,表明亚硝酸盐胁迫促进了对虾的无氧代谢,脂质的β氧化被抑制,从而导致脂质动员的延迟,干扰了对虾的正常供能。
此外,在应激条件下虾类甲壳动物还会进行蛋白质代谢以维持正常能量供给。Xu等[68]研究发现,南美白对虾在长期低盐胁迫下会激活相应的信号转导通路来响应应激因子,并通过调节糖代谢和能量代谢来维持机体的平衡(图2)。Fan等[69]对冷胁迫下的南美白对虾进行蛋白质组学分析,发现在氨基酸代谢中,大部分差异表达基因(DEPs)表达上调,说明氨基酸是对虾应对冷胁迫的主要能量来源。另外,在应激条件下特定的游离氨基酸(如甘氨酸、脯氨酸和丙氨酸等)也会参与对虾的渗透调节[70]。综上所述,在应激过程中,虾类水产动物可以通过一系列有序的调控来维持机体的动态平衡。
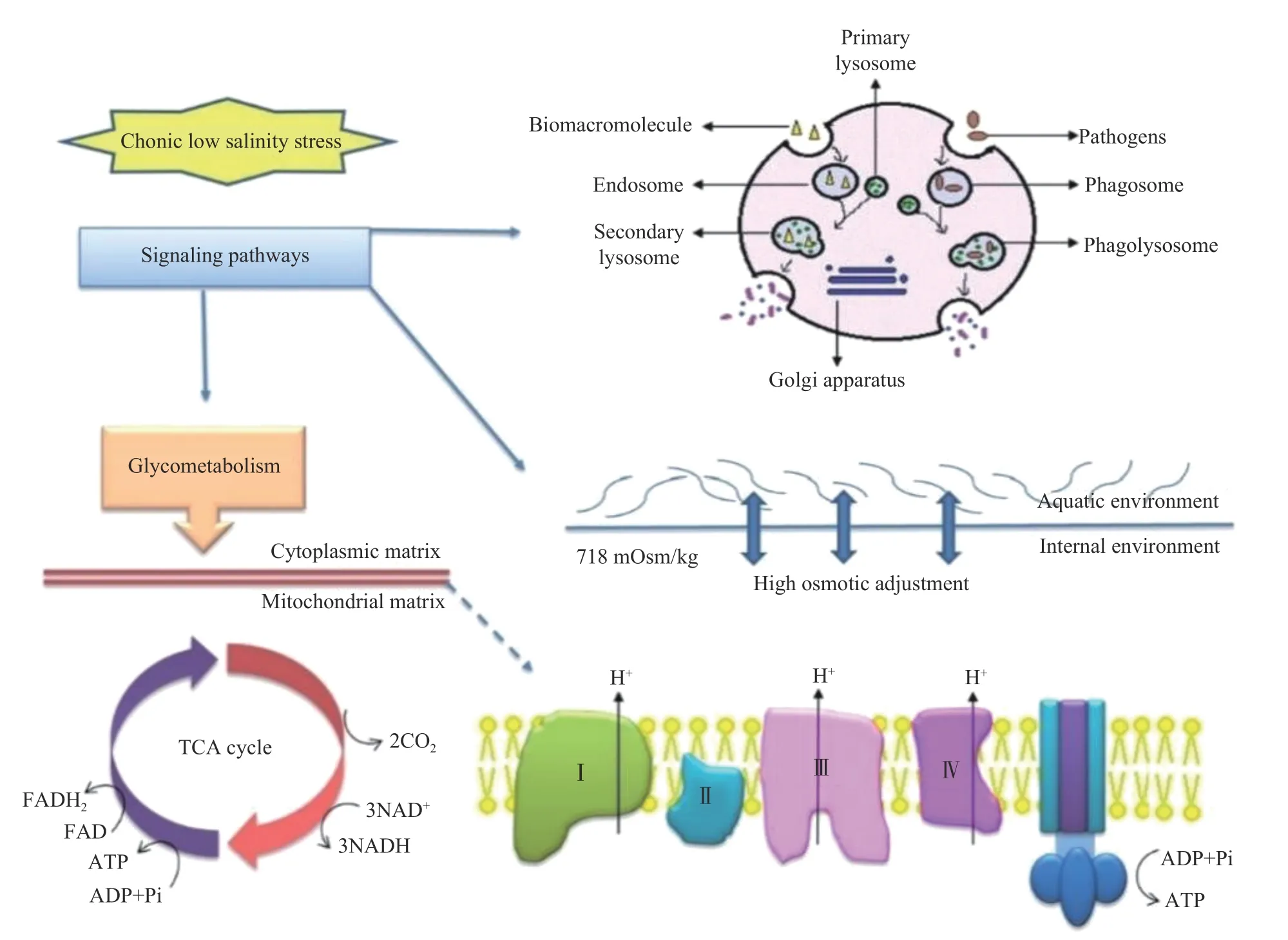
图2 凡纳滨对虾响应低盐胁迫的蛋白质组学分析及相关途径[68]Fig.2 Deduced schema of L. vannamei responding to low salinity stress according to the significantly changed proteins by proteomic analysis and related pathways[68]
2.1.4 免疫系统 虾类甲壳动物的免疫系统是非特异性的,主要包括细胞免疫和体液免疫两个方面[71]。环境因子对甲壳类动物免疫功能的影响研究主要集中在总血细胞计数和分类血细胞计数、抗菌活性、吞噬活性、酚氧化酶原激活系统以及相关的体液免疫因子[72-73]。当虾类甲壳动物受到环境胁迫时,免疫系统会产生免疫反应,以减少相关的有害影响。徐进等[74]在30和35 ℃水温下对克氏原螯虾进行高温应激试验,发现克氏原螯虾血淋巴中的总超氧化物歧化酶(T-SOD)和溶菌酶(LYZ)含量在应激后都出现了显著性降低,表明机体的抗氧化能力和对病原微生物的抵抗力减弱,应激对机体的免疫机能产生了负面影响。Dai等[75]研究也发现,虾在短期饥饿压力下溶菌酶(LZM)、酸性磷酸酶(LZM)和超氧化物歧化酶(SOD)的活性显著升高,直接影响虾的免疫活动。一般来说,强烈或持久的环境胁迫对虾类甲壳动物的先天免疫功能都有负面影响,只是不同的环境因子影响略有不同。
另外,通过转录组学和蛋白质组学技术研究发现,甲壳动物体液免疫信号通路中的一些基因在应对环境胁迫时存在差异表达,表明环境胁迫可能会影响甲壳动物体液免疫信号通路,这些信号通路主要包括TLR/IMD-NF-κB、JAK-STAT、MAPK和Wnt等[76]。于杰伦[77]利用转录组学的研究方法发现,日本对虾在亚硝酸盐胁迫转录组测序中共鉴定出825个差异表达基因,另外RNA转运通路、溶酶体通路、吞噬体通路和JAK-STAT信号通路均被富集,表明相关免疫应答基因和信号通路在日本对虾受亚硝酸盐胁迫时,对机体有一定的保护作用。Luo等[78]研究发现,在急性冷应激12 h后,对虾鳃中p38MAPK的转录水平和p38MAPK蛋白的磷酸化均显著上调,表明p38MAPK信号通路在凡纳滨对虾对急性冷胁迫的响应中起着重要作用。Xu等[68]利用蛋白质组学方法对凡纳滨对虾的综合生理反应进行研究发现,在低盐胁迫下对虾中Wnt信号通路、Foxo信号通路、G蛋白信号通路和酪蛋白激酶均显著变化,表明凡纳滨对虾可以通过改变多种信号途径来响应盐胁迫。
综上所述,虾类甲壳动物在不同环境因子胁迫下,其生理生化以及一定程度上的免疫学参数都会发生变化,以应对这些压力对机体造成的负面影响,但高强度的应激会对机体造成不可逆的损伤,从而导致体内平衡失衡,因此,在运输过程中应合理控制外界条件,以保持虾更高的存活量。
2.2 运输应激对虾类肌肉品质的影响
肌肉是虾体内生化能量转化的主要组织,也是维持体内平衡的最大能量库和氨基酸储存库[79-80]。研究表明在运输应激状态下虾的机体会动员能量代谢相关的理化反应以适应应激,包括糖代谢、脂代谢和蛋白质代谢,而在这些物质能量代谢转变过程中会积累大量的代谢物质,如乳酸、脂肪酸、氨基酸等,从而影响虾的肌肉品质[81]。运输应激还会使虾类肌肉持水能力下降、硬度弹性降低,这可能是由于蛋白质和脂质的结构和组成变化所致[82]。此外,虾类的应激反应还会使肌原纤维蛋白溶解度降低,肌原纤维内间隙增加、肌肉裂解面积增大,进而导致虾类肌肉品质劣化[83]。一般不同的运输应激类型对虾肉理化特性的影响不同。当虾体在受到温度、低氧、酸碱或饥饿胁迫时主要产生氧化应激反应和能量代谢异常,使虾体内生理功能失调,脂质氧化和蛋白质氧化水平升高,进而影响虾肉的营养品质,而拥挤和机械损伤胁迫主要影响虾的蛋白质结构,使虾肉质构发生变化,降低虾肉的食用品质(表2)。

表2 运输应激对虾肉理化特性的影响Table 2 Effects of transportation stress on physicochemical properties of shrimp meat
3 结语
虾类甲壳动物在活体贮运过程中会受到各种因素的影响使机体产生氧化应激、内质网应激,并使能量代谢和免疫系统发生改变,对虾机体健康和肌肉品质产生负面影响。因此,了解影响运输应激的因素以及运输中的健康管理对虾的活体贮运至关重要。在未来对活虾运输中的健康管理可以从以下几方面来实施:在虾的捕获和分拣中应避免机械胁迫对虾体造成的损伤,并及时丢弃软壳、体表发白和濒临死亡的虾体,确保适宜的运输密度;在运输途中应避免虾直接暴露在空气和阳光下造成氧化损伤,并加强对虾体运输环境的实时监测,确保所有参数保持在最佳水平,同时可以使用麻醉剂或探索新型的运输方式和运输材料来降低运输压力;严格执行贮运标准,实施科学化和标准化管控模式,加强冷链物流发展,同时相关部门应积极落实鲜活虾类水产品运输“绿色通道”政策,做好物流保障。总而言之,需要不断从各方面优化贮运条件,以提高虾类运输的存活量,减少经济损失。同时,为了促进创新和技术的发展,企业、科研机构和工业之间应协同作用,确保虾类运输技术能与时俱进,适应市场需求。
