104份黑龙江省水稻资源株型性状比较及遗传多样性分析
2022-12-10张希瑞王敬国王立楠
张希瑞, 王敬国, 王立楠
(1.黑龙江省农业科学院水稻研究所,黑龙江佳木斯 154026; 2.东北农业大学,黑龙江哈尔滨 150030)
水稻作为全世界一半以上人口的口粮,其安全生产对保障国家粮食安全至关重要[1]。黑龙江省作为北方粳稻的主产区,水稻资源丰富,但是将遗传多样性及株型演化相结合的研究较少,这不利于育种家们充分合理利用种质资源。研究黑龙江省不同类型水稻种质的遗传多样性,有利于充分挖掘、利用现有水稻资源,为育种家们合理选配亲本、培育理想株型新品种提供理论依据。
水稻株型即植株形态在空间上的分布,与水稻产量关系密切[2]。Donald于1968年提出了“理想株型”的概念,理想株型即能够最大限度利用光能来提高作物产量的株型模式[3]。随着此概念的提出,许多学者提出了多种水稻高产模式。陈温福等确立了利用籼粳远缘杂交创造新株型,再通过复交或回交聚合有利基因创制出超高产理想株型[4]。侯立刚等通过对吉林省主推的不同熟期水稻品种株型及穗部形态的研究,得出了吉林省水稻株高处于较低水平和不同品种穗部形态以中、小散穗和半直立穂型为主的结论[5]。周开达等提出了“重穂型”的超高产杂交水稻育种理论[6]。黄耀祥提出“矮秆蘖强早发”的水稻模式[7]。袁隆平提出“冠层高、穗大、穗层矮”的水稻模式[8]。韦还和等研究发现,在穗长度、平均穗粒数、一次枝梗数和二次枝梗数4个性状上超高产水稻群体显著高于其他水稻群体[9]。以上研究旨在通过改善水稻株型来提高其产量。
结合简单重复序列(SSR)分子标记进行水稻资源遗传多样性研究的报道很多,如杨静等利用52个SSR分子标记分析了54份黑龙江省水稻材料的遗传多样性[10]。李红宇等利用53对SSR引物分析了107份东北三省主栽水稻品种的遗传多样性,并得出黑龙江省水稻品种的遗传多样性大于吉林省和辽宁省水稻品种的结论[11]。王复标等利用60对SSR标记对190份水稻材料的遗传多样性和群体结构进行了分析[12]。郝伟等以35份东北三省水稻区域试验品种为材料,结合SSR标记对其遗传多样性进行了分析,并检测出237个等位基因[13]。宋泽等选用56对SSR引物分析了贵州省173份糯稻品种的遗传多样性,共检测到191个等位基因[14]。而目前关于黑龙江省地方品种和不同年代育成品种遗传多样性的研究较少。本研究利用152对SSR分子标记结合21种株型性状对来自黑龙江省的104份水稻种质的遗传多样性和株型演化规律进行分析,旨在为优化亲本选配和科学利用种质资源提供理论依据。
1 材料与方法
1.1 性状调查
104份参试材料由黑龙江省农业科学院水稻研究所提供,连续2年(2019年和2020年)种植于水稻研究所科研选种田,田间管理同生产田。依据不同水稻材料的育成年份将供试材料分为4组:组群Ⅰ包括53份地方品种;组群Ⅱ为9份1958—1970年育成的品种;组群Ⅲ是17份1971—1999年育成的品种;组群Ⅳ包括25份2000年及之后育成的品种(表1)。田间设计、操作参照张希瑞等的方法[15]。
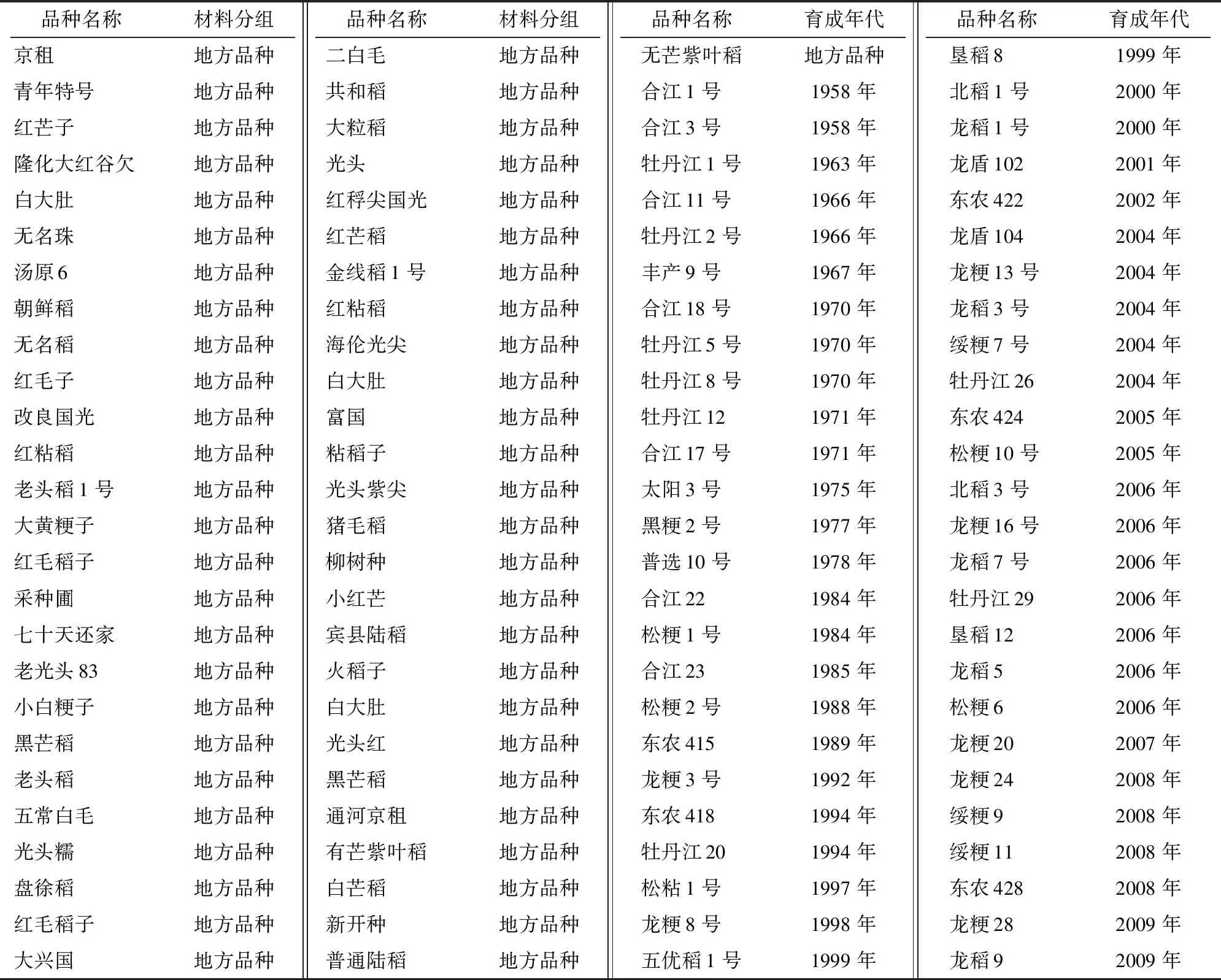
表1 供试品种
在供试材料齐穗后测定每份材料的株高、穗数、穗下第1至第4节间长度、穗抽出度、穗长、剑叶长度、剑叶宽度、倒二叶长度、倒二叶宽度、倒三叶长度和倒三叶宽度。另外,对穗颈角度、剑叶角度、倒二叶、倒三叶角度的测量参照徐正进等的方法[16]。
1.2 总DNA的提取
以每个参试品种的幼嫩叶片为样本,依据Chen等的方法[17]进行总DNA的提取和纯化,并利用琼脂糖凝胶电泳对提取的DNA质量进行检测。
1.3 引物合成及产物检测
参照Gramene网站(https://www.gramene.org)上的引物信息,委托哈尔滨博仕生物技术有限公司对均匀分布在水稻12条染色体上的152对引物进行合成,用于遗传多样性分析。PCR反应体系和产物检测参照王敬国的方法[18]。
1.4 数据分析
株型数据分析在DPS 9.50软件[19]中进行。遗传距离[20]、遗传一致度、基因流(Nm)和遗传分化系数(Fst)[21]均在POPGENE 1.32软件[22]中进行计算。分子数据的观测等位基因数量(Na)、遗传多样性指数(He)和多态性信息含量(PIC)[23]均在PowerMaker 3.25软件[21]中进行计算。在NTSYS-pc 2.1软件[24]中利用非加权配对算术平均法(UPGMA)对供试材料进行聚类分析。分子方差分析在GenAlEx 6.2软件[25]中进行。
2 结果与分析
2.1 性状变异
由表2可知,2年间变异系数最大的5个性状分别是穗下第1节间长度、倒二叶基角、穗颈基角、穗抽出度和倒二叶张角,其变异系数分别为0.515 5、0.388 3、0.369 9、0.363 7和0.340 9,变异系数最小的5个性状分别为穗下第4节间长度、株高、剑叶长、剑叶宽和倒二叶宽,其变异系数分别为0.091 6、0.110 4、0.114 5、0.116 1和0.127 9。就变异系数而言,穗颈张角、剑叶张角、倒二叶张角和剑叶基角、倒三叶张角在2年里都具有较大的变异系数,而倒二叶宽、倒三叶宽、剑叶宽、穗颈基角和穗下第1节间长度年际间相对稳定。
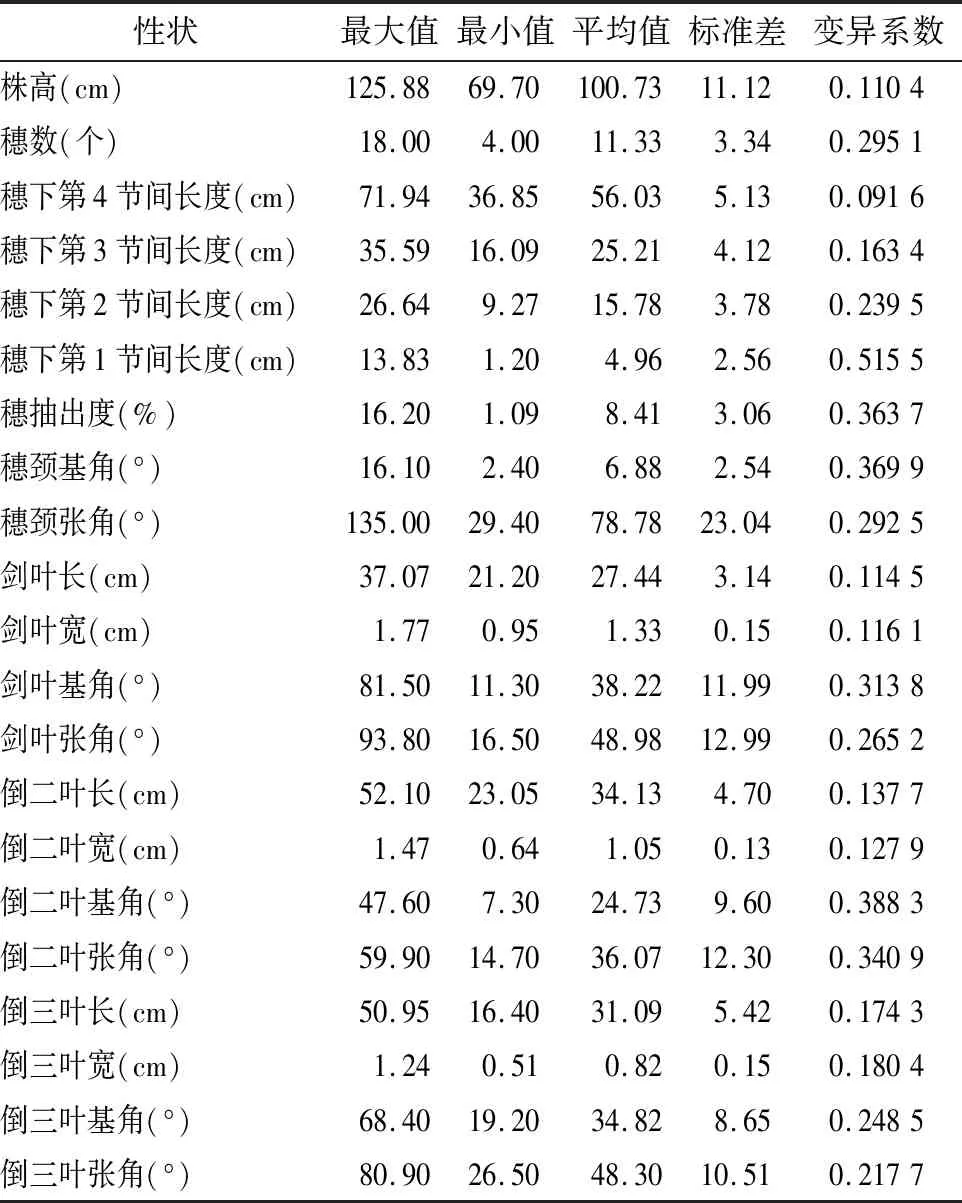
表2 供试材料2年各株型性状的均值统计
2.2 株型性状的差异分析
2019年和2020年供试材料株型性状年份间的多重比较结果见表3。不同年代品种的穗部呈现如下的特征:地方品种和1958—1970年间品种的穗抽出度均大于另外2个年代的品种;地方品种的穗抽出度极显著大于其他年代育成的品种,2000年及以后育成的品种在2年里均具有最小穗抽出度。1971—1999年品种的穗数大于其他年代的品种。在2年里,1958—1970年育成的品种具有较大的穗颈张角,1971—1999年的品种具有最小的穗颈张角。以上差异分析结果表明,育成品种的穗数增多,而穗抽出度逐渐缩小,弯曲穗型是黑龙江省水稻品种的主要穗型。
对比参试材料的剑叶性状可以看出,2年里,2000年及以后育成的水稻品种的剑叶宽度均大于其他2个年代育成的品种;就剑叶基角和张角而言,2年里均呈现的趋势是1958—1970年育成的品种>1971—1999年育成的品种>2000年及以后育成的品种。
对比参试材料的倒二叶性状可以看出,就倒二叶长和宽而言,2年里均呈现的趋势是2000年及以后育成的品种>1971—1999年育成的品种>1958—1970年育成的品种。按照倒二叶基角和张角排序,2年里呈现的趋势是1958—1970年育成的品种>1971—1999年育成的品种>2000年及以后育成的品种。地方品种在2年里倒二叶基角均极显著大于1971—1999年育成的品种和2000年及以后育成的品种。
参试材料倒三叶性状的比较结果表明, 在2年里,1958—1970年育成的品种具有最小倒三叶长度,且与其他年代品种间均差异极显著;2年中,地方品种具有最小倒三叶宽。地方品种的倒三叶基角和张角在2年里都显著大于1971—1999年育成的品种和2000年及以后育成的品种,2000年及以后育成的品种在2年里有最小的倒三叶基角和张角。上述有关剑叶、倒二叶和倒三叶的性状多重比较结果表明,育成品种的剑叶宽度和倒二叶宽度呈现增大的趋势,而剑叶基角和张角呈现减小的趋势。
2年中,1958—1970年育成的品种的植株株高均极显著低于其他组水稻品种,地方品种的株高在2年里均极显著高于其他年代育成的水稻品种,这说明地方品种株高最高。从节间长度的角度分析,2年里地方品种的穗下第1、第2和第3节间长度均极显著大于其他3组,这可能是导致地方水稻品种株高显著高于其他年代品种的主要原因;1958—1970年代育成品种的穗下第3和第4节间长度都低于其他年代品种,这在一定程度上导致了这一时期的品种株高较低。
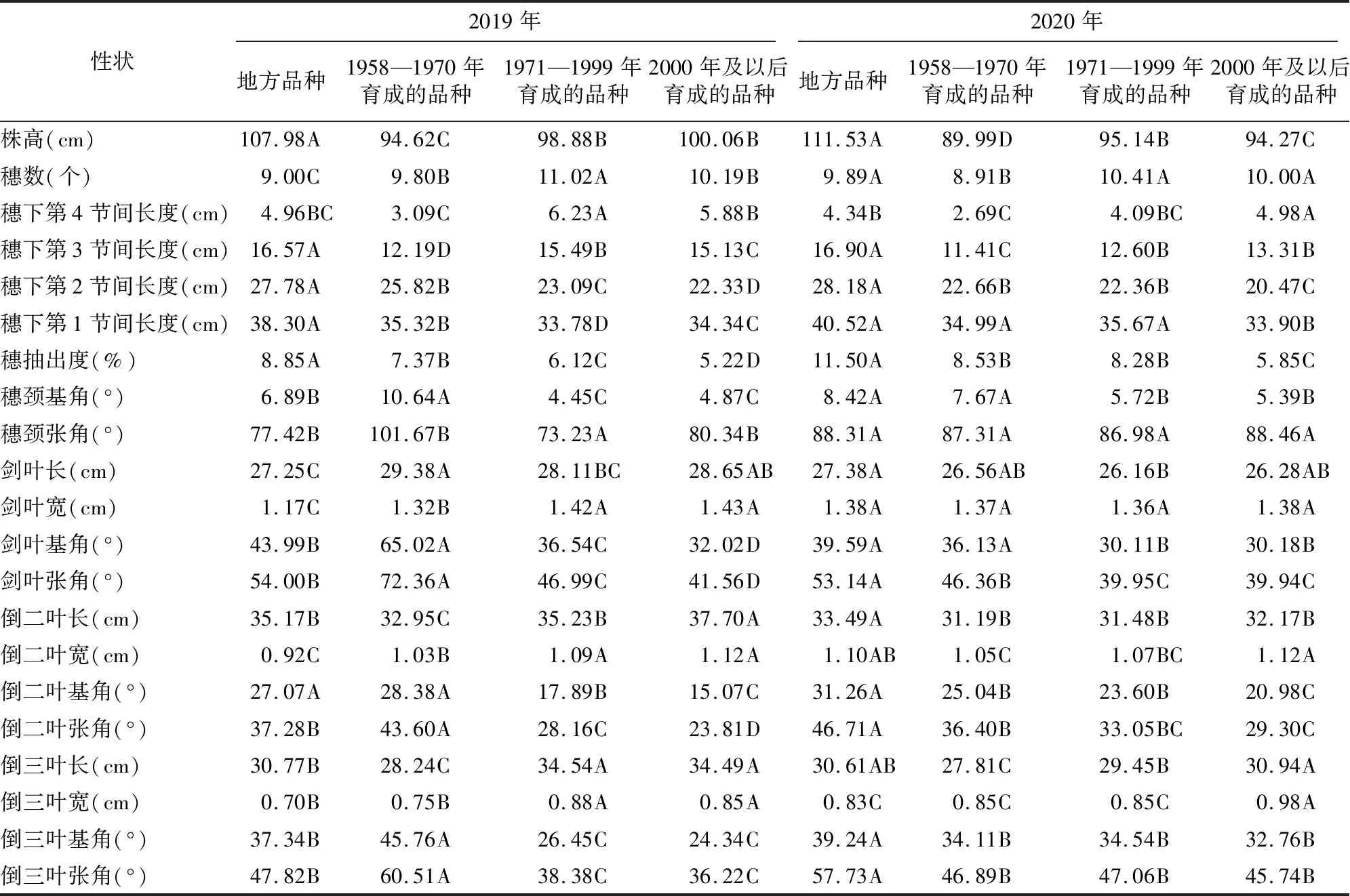
表3 2019年和2020年株型性状的比较
从上述株型差异分析结果可以归纳出参试的黑龙江省材料的株型特点:地方品种具有最大株高,穗下第1、第2、第3节间长度,穗抽出度;1958—1970年育成的品种具有最小穗下第3、第4节间长度;1971—1999年育成品种具有最大穗数;2000年及以后育成的品种具有最小穗抽出度、剑叶张角、倒二叶基角和张角、倒三叶基角和张角和最大剑叶宽度和倒二叶宽度。此外,弯曲穗型是黑龙江省育成品种的主流穂型。
2.3 SSR引物多态性分析
本研究利用152对SSR引物共检测到749个等位基因变异。观测等位基因数量(Na)的范围为1~9,平均值为3.425。基因遗传多样性指数(He)的范围为0.037~0.870,平均值为0.490。Shannon信息指数(I)为0.094~2.076,平均值为1.158。多态性信息量(PIC)范围是0.091~0.857,平均值是0.597。Na、He和I排名前十的位点分别是分布在第9、第3、第7、第7、第2、第2、第6、第10、第1和第8号染色体上的RM257、RM1350、RM336、RM1353、RM1347、RM207、RM1369、RM1374、RM1381和RM264。
2.4 不同群体间的遗传多样性
方差分析结果显示,组群内和组间都存在极显著差异(P<0.01)。其中组群间变异为8%,组群内变异为92%(表4)。
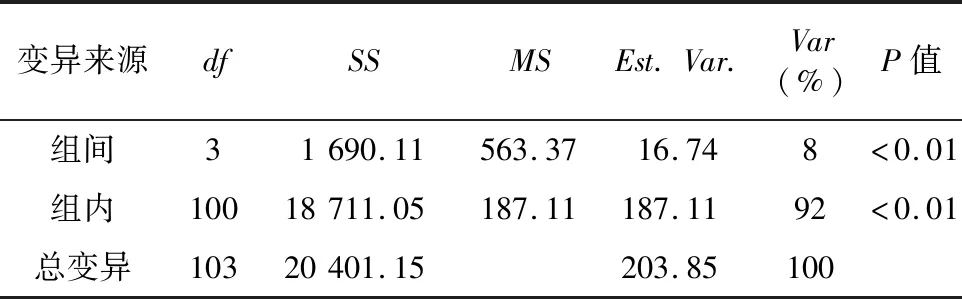
表4 分子方差分析结果
遗传距离和遗传一致性分析结果(表5)表明,地方品种与其他3个年代育成品种之间具有最大遗传距离(0.210 7、0.188 8和0.185 3)和最小遗传一致性(0.810 1、0.828 0和0.830 9)。1971—1999年育成的品种与2000年及以后育成品种间具有最小遗传距离(0.072 0)和最大遗传一致性(0.930 5)。这说明黑龙江省地方水稻品种与其他年代育成水稻品种间存在明显差异。
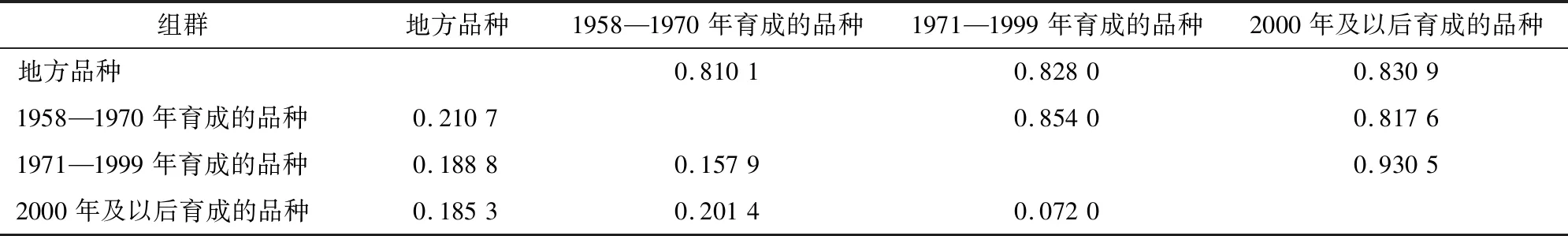
表5 组间Nei’s 遗传距离(左下)和 遗传一致性(右上)
不同组群间遗传分化系数(Fst)和基因流(Nm)的分析结果(表6)表明,组群间Fst最大值为0.104,最小值为0.040,平均值为0.081,这表明各组群间的变异为8.1%,组群内的变异为91.9%,这与方差分析的结果大致相同。不同组群间的Nm最大值是5.715,最小值是2.153,平均值是3.133。地方品种与其他3个年代育成品种间具有最大遗传分化系数(0.104、0.076和0.071)和最小基因流(2.153、3.079和3.217)。1971—1999年育成的品种与2000年及以后育成品种之间遗传分化系数最小(0.040),基因流最大(5.715)。这与上述的遗传距离和遗传一致性分析结果一致。
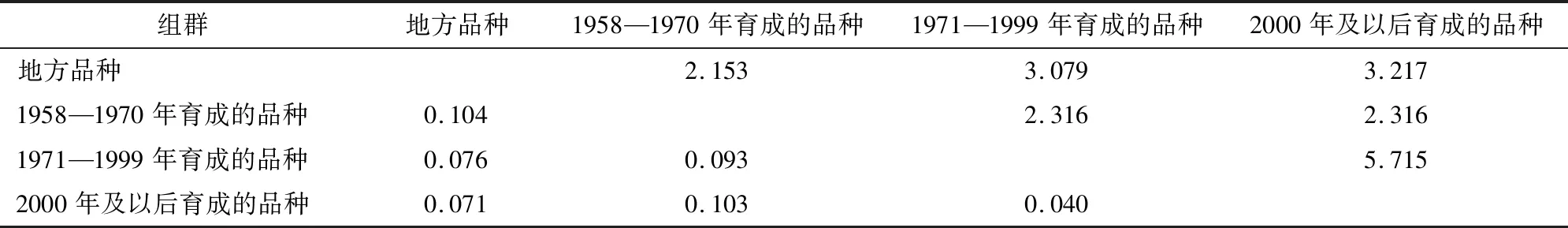
表6 组间分化系数(Fst,左下) 和基因流(Nm,右上)
2.5 聚类分析
当遗传相似系数取值为0.676 3时,将供试的104份材料划分为4个亚群(图1)。第1亚群、第2亚群和第3亚群全部为地方品种,分别包含14、7、2个地方品种。第4亚群包括全部的育成品种,包括30个地方品种和51个育成品种。供试品种间的相似系数全部在0.664 1以上,表明供试材料之间遗传差异较小,遗传背景狭窄。
当遗传相似系数取值为0.668 4时,又将第4亚群再分为6个小亚群。第1小亚群包含龙粳13和松粘1号2个育成品种;第2小亚群包含2个育成品种,分别是东农424和北稻3号;第3小亚群包含5个地方品种;第4小亚群包含12个育成品种,除东农415(1989年选育),其他11个均为2000年及以后育成的品种;第5小亚群包括2个育成品种和23个地方品种,育成品种分别是牡丹江8号(1970年)和合江17号(1971年);第6小亚群包括33个选育品种和7个地方品种,选育品种中龙盾102、龙盾104、龙稻3号、绥粳7号、松粳10号、龙粳16号、龙稻7号、牡丹江29和垦稻12为2000年及以后选育的品种,其他为1958—1999年选育的品种。以上分析结果表明,供试品种的聚类结果与其育成年代密切相关。
3 结论与讨论
3.1 株型演化
水稻株型即水稻植株在空间上的排列方式。前人关于水稻株型的研究有很多,如胡月等将日本粳稻品种和黑龙江省粳稻品种的株型性状进行了比较,结果表明黑龙江省品种的株高、上3节间长度、上3叶的长和宽都显著大于日本品种,而日本品种叶片的直立性优于黑龙江省的品种[26]。解文孝等研究了2015年辽宁省参试的不同熟期水稻材料的株型特征,发现早熟粳稻品种主要为弯曲、轻穂型,中熟粳稻品种多为直立、重穂型,不同熟期品种的叶宽表现为中晚熟品种>中熟品种>早熟品种,二次枝梗结实率呈现出早熟品种>中晚熟品种>中熟品种的规律[27]。莫永生等以4个不同年代育成的水稻品种为材料,并对其株高、根干质量、第2茎节直径、茎壁厚度与产量的关系进行了分析,结果表明供试材料的产量会随根干质量、株高、第2茎节直径、茎壁厚度的增大而提高[28]。本研究通过对104份黑龙江省水稻材料21种株型性状的多重比较及差异性分析,发现黑龙江省不同年代品种的株型特点,即地方品种的株高、穗下第1节间长度、穗下第2节间长度、穗下第3节间长度、穗抽出度均最大;1958—1970年育成的品种具有最小穗下第3、第4节间长度;1971—1999年育成品种的穗数最多,且大于其他年代育成品种;2000年及以后育成的品种的穗抽出度、剑叶基角和张角、倒二叶基角和张角、倒三叶基角和张角都最小,剑叶宽度和倒二叶宽度均最大。育成品种的剑叶宽度、倒二叶宽度逐渐增大,株高降低,穗抽出度、剑叶基角和张角、倒二叶基角和张角、倒三叶基角和张角逐渐缩小;弯曲穗型是黑龙江省育成品种的主流穗型。
3.2 SSR标记分析遗传多样性
利用SSR标记的多态性研究不同区域水稻品种的遗传多样性的报道有很多,如陈于敏等利用48对SSR标记引物分析了81份高原粳稻品种的遗传多样性,共检测到139个等位基因,并得出大多数品种间遗传距离近,遗传基础较窄的结论[29]。马孟莉等利用100对SSR引物对60份云南省哈尼梯田红米地方品种的遗传多样性进行了分析,结果表明,云南省哈尼梯田红米水稻品种具有丰富的遗传多样性[30]。李松等利用18对SSR引物分析了20份云南省腾冲市水稻品种的遗传多样性,结果表明腾冲市糯谷和推广品种具有丰富的遗传背景[31]。唐如玉等利用25对SSR引物分析了三峡库区的81份水稻资源的遗传多样性和群体结构,结果显示供试材料被分为2个类群,地方品种的平均有效等位基因数高于选育品种[32]。本研究中152对SSR引物共检测到749个等位基因变异,Na、He、I和PIC的均值分别为3.425、0.490、1.158和0.597,并得出黑龙江省地方水稻品种与其他不同年代水稻品种间遗传差异最大的结论。
3.3 黑龙江水稻种质间的亲缘关系
聚类分析结果表明,在遗传相似系数为 0.676 3 处,104份供试材料被聚类为4个亚群,所有供试材料间相似系数均大于0.664 1,表明其遗传相似性较大,供试材料遗传距离较近,遗传基础狭窄,且绝大多数育成材料聚合为一类,再一次证明了黑龙江省水稻种质资源间的亲缘关系较近,遗传背景狭窄,这一结果与张科等的结论[33]相符。追溯黑龙江省水稻系谱,许多育成材料均含有骨干亲本石狩白毛、上育397、藤系138和农林8号及由它们衍生品种等的血缘,虽然黑龙江省水稻地方品种较丰富,但利用率低下,骨干亲本多引自日本,这可能是黑龙江省水稻种质遗传背景狭窄的主要原因。为拓宽黑龙江省水稻遗传基础,育种工作者们应广泛引入我国其他各地区优异的水稻种质资源,并利用杂交手段聚合有利基因,培育出适合黑龙江省栽培的优质水稻品种。
