中国27个国家级自然保护区苔藓植物地理成分构成差异与环境和空间因素的关系
2022-12-09周徐平唐录艳何卓冀王顺莉黄丹李大华邓欣妍侯梦丹杨书林彭涛
周徐平 ,唐录艳,何卓冀,王顺莉,黄丹,李大华,邓欣妍,侯梦丹,杨书林,彭涛*
1. 贵州师范大学生命科学学院/生物多样性研究中心/中国-东盟苔藓植物研究中心,贵州 贵阳 550025;2. 深圳市中国科学院仙湖植物园/深圳市南亚热带植物多样性重点实验室,广东 深圳 518004;3. 贵州省瓮安中学,贵州 瓮安 550400
植物区系地理成分,是以植物分类学为基础,基于植物种或者其他分类单元的现代地理分布来进行划分,可以归为若干分布型(王荷生,1992)。吴征镒(1991)将中国种子植物属划分为15大分布区类型和31个变型,建立了完备的理论框架,如今的植物区系研究大多是基于此框架进行。苔藓植物是一类多样性仅次于被子植物的高等植物,是生物多样性的重要组成部分(Patiño et al.,2018)。早在20世纪60年代,我国苔藓植物学之父陈邦杰教授就对黄山进行过苔藓植物区系研究,并将地理成分划分为泛北极分布、东亚分布和旧热带区分布等5种分布型(陈邦杰等,1965),但是该系统在后期被采纳的并不多,目前国内多数学者认为苔藓植物的分布与种子植物密切相关,因此将吴征镒(1991)的划分方法运用于苔藓植物。但苔藓植物是一类有着和种子植物不同生物地理格局的陆生植物(Embryophyta)单系分支,相较于种子植物,其具有更高的长距离扩散能力,分布范围更广,潜在的气候生态位更大(Patiño et al.,2018;Zanatta et al.,2020;Su et al.,2021)。部分学者也注意到了苔藓植物分布的此特殊性,提出了不同的分布区类型划分方法,例如Zhang et al.(2003)、于晶等(2001)和申琳等(2019)的研究。
在植物区系研究中,研究者们为了获得研究区现代植物区系的性质、来源和地位,通常会将研究区区系成分与邻近区域进行比较(李嵘等,2017)。苔藓植物区系是苔藓植物研究的热点话题,以往的研究中,地理成分分异的驱动因素主要归因于空间距离、气候、纬度和海拔等,例如李粉霞(2006)48-50、田晔林(2010)40-43和周徐平等(2022)的研究。将地理成分与现代环境因子和空间因子结合以定量揭示地理成分相似性原因的报道较少,这阻碍了人们对植物区系更深一步的认识。吴征镒等(2010)1-120和沈泽昊等(2017)也曾指出,针对已建立的中国植物区系地理分类与分区体系,还有待针对特定的研究类群或区域,从历史、环境和进化角度进行验证及不同尺度的深入研究。在种子植物中,沈泽昊等(2017)基于地理距离、气候因子和面积等参数,使用Mantel检验对驱动中国高山植物区系地理成分分异的因素进行了探讨,认为地理距离是主要的驱动因素,其次是气候因子。在苔藓植物中也有学者进行过类似的研究,曹同等(2000)基于典范对应分析(CCA)将长白山等9个山地的苔藓植物地理成分与5个影响因子结合初步揭示了影响其成分相似性的因素,但存在研究地区较少,影响因子单一的问题。
自然保护地,是生物多样性富集区、代表性的自然生态系统与关键物种天然集中分布区,对科学研究和生物多样性保护具有重要意义(王伟等,2016)。苔藓植物是保护区中重要的组成部分,在水分平衡、侵蚀防治、氮收支、全球碳循环、森林群落演替和更新等方面发挥着重要功能,同时也能够为其他生物提供栖息场所(Turetsky,2003;Goffinet et al.,2009)。鉴于自然保护区的特殊性,本研究基于19个省和2个直辖市内27个国家级自然保护区的苔藓植物物种数据,以及中心地理坐标、气候因子、黏土含量和海拔等参数,拟回答以下问题:(1)分析27个保护区的苔藓植物分布区类型组成,阐明不同分布区类型在经纬度上的分布格局;(2)揭示影响27个保护区间苔藓植物地理成分相似性的因素。
1 数据与方法
1.1 物种数据
为了揭示影响苔藓植物地理成分相似性的因素,本研究获取了19个省和2个直辖市内27个国家级自然保护区的苔藓植物物种数据,其中,白石砬子等 17个地区的物种数据来源于公开发表的期刊或书籍,其余 10个地区的物种数据来源于硕博论文(表1)。考虑到不同时期使用的分类系统可能会对结果产生影响,我们参照2013版《中国生物物种名录》(贾渝等,2013)对27个地区苔藓植物的种名进行校验和物种归并,得到各地区苔藓植物物种更新名录。重点保护物种根据《国家重点保护野生植物名录》(http://www.forestry.gov.cn/main/3954/20210908/163949170374051.html)进行确定。受威胁苔藓植物参照《中国高等植物受威胁物种名录》(覃海宁等,2017)进行确定。

表1 27个国家级自然保护区的基本统计信息Table 1 The basic information of 27 national nature reserves
1.2 地理成分数据
由于苔藓植物与其他分类类群有着不同的生物地理格局,我们选择采用Zhang et al.(2003)关于香港苔藓植物的分布区类型系统对校对后的各保护区的苔藓植物进行分布区类型划分,据此得到各保护区各个种的分布区类型并进行了分布区类型统计(表2)。

表2 各保护区的苔藓植物分布区类型构成Table 2 The composition of bryophyte distribution types in each reserve
由于部分保护区间的面积和物种丰富度差异较大(表1、2)。面积数据来自中华人民共和国生态环境部(https://www.mee.gov.cn/searchnew/?searchword= 自然保护区名录),单位转换为km2。为了较大程度地削弱保护区间的面积和物种丰富度差异对地理成分组成的影响,苔藓植物地理成分组成采用了植物区系谱(floristic spectrum,FS)(马克平等,1995),为更加充分地反映出地理成分的组成特点,在进行 FS统计时,世界分布类型(Cosmopolitan)和未知分布类型(Unknown)不加以考虑(Zhang et al.,2003;马克平等,1995)。为了降低在进行分布区类型划分时主观因素对结果造成的影响,我们参照Zhang et al.(2003)的标准将除世界分布和未知分布类型外的 12种分布区类型进一步归为 4种地理成分,即温带成分(T1—T2)、热带成分(T3—T4)、东亚成分(T5—T8)和亚洲-澳大利亚-大洋洲成分(T9—T12)。
植物区系谱计算公式为(马克平等,1995):

式中:
Fi——某地区第i个分布区类型所占比率(i=1,2, 3, ……);
Ei——某地区第i个分布区类型的苔藓植物物种数(i=1, 2, 3, ……);
T——某地区除世界分布和未知分布类型外的苔藓植物总种数。
1.3 影响因子数据
1.3.1 环境因子
为探讨不同环境因素对苔藓植物地理成分相似性的相对影响,我们选取了8个气候指标,分别是年均温、最热月最高温、最冷月最低温、年均太阳辐射、年均降水、湿润指数、年均潜在蒸散量及海拔。海拔能够引起温度变化,进而对环境湿度产生影响(Körner,2007),代表一定的热量梯度,因此我们将保护区内最高海拔数据也加入气候矩阵。海拔数据来自美国国家航空航天局的数字高程模型(http://www2.jpl.nasa.gov/srtm/)。同时选取1个土壤质地数据——黏土含量。已有研究都证明这些因子与苔藓植物的分布或植物地理成分的构成密切相关(Chen et al.,2015a,b;Qian et al.,2016;沈泽昊等,2017;田悦等,2022)。其中湿润指数和黏土含量来源于中国科学院资源环境科学数据中心(https://www.resdc.cn/Default.aspx)。年均潜在蒸散量来源于全球干旱指数和潜在蒸散气候数据库(https://cgiarcsi.community/category/data/)。其余气候数据来源于世界气候数据网站(https://worldclim.org/data/worldclim21.html)。以上数据集中,借助ArcGIS(10.6)软件通过“按掩膜提取”和“获取栅格属性”方式提取保护区内湿润指数的均值,其余数据通过将保护区重新采样成1 km×1 km 的网格进行提取并计算均值,各个保护区的功能区划来源于中国自然保护区标本资源共享平台(http://www.papc.cn/html/folder/946895-1.htm)。
1.3.2 空间因子
用各保护区几何中心点的经纬度(表 1)表示该区所处地理位置(沈泽昊等,2017)。对于中心点坐标的获取,若保护区由单个面组成,则借助ArcGIS(10.6)软件直接获取;若保护区由多个面组成,则将保护区重新采样成1 km×1 km的网格,据此获取中心点坐标,然后利用R(4.0.3)软件中SoDA包的geoXY函数将其转化为笛卡尔坐标。空间变量的构建结合转换后的坐标采用 adespatial包中的dbmen函数进行,仅设置参数“silent=FALSE”,表示不对截断水平(truncation level)和计算dbmem的时间进行打印,这一步产生了4个空间变量,即dbMEM1、dbMEM2、dbMEM3和dbMEM4,用于后续的变差分解分析。
1.4 普通最小二乘回归分析
选用普通最小二乘回归(基于R (4.0.3) 软件中的l m函数)拟合重新划分的4种苔藓植物地理成分在纬度和经度上的分布格局,为消除数据本身非正态性分布对回归模型产生的影响,在进行回归模型前,对原始数据进行了log10(x+1)转换。
1.5 地理成分相似性分析
保护区间苔藓植物地理成分的相似性用Jaccard相似性指数进行度量(沈泽昊等,2017),采用R(4.0.3)软件中vegan包的vegdist函数进行计算,其公式为(Oksanen et al.,2022):
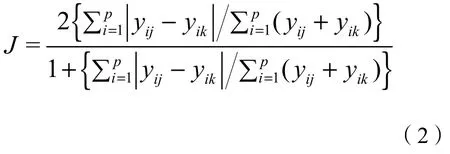
式中:
J——地理成分Jaccard相似性指数;
p——分布区类型数(保护区-分布区类型矩阵中的分布区类型数);
yij——分布区类型i在保护区j中所占比率;
yik——分布区类型i在保护区k中所占比率。
1.6 变差分解分析
在大尺度上,当调查的区域相对于整个区域而言,其覆盖率较低时,由Mantel检验所获得的结论可靠性下降(韩博平等,2022),而Lai et al.(2022)将多元回归中“层次分割(hierarchical partitioning)”的概念引入典范分析,能够定量评估共线性解释变量的相对重要性。因此,为了定量评估环境因子和空间距离对苔藓植物地理成分相似性的影响,本研究基于Lai et al.(2022)的方法在R(4.0.3)软件中进行了变差分解分析。分3步进行,(1)将地理成分与4个空间变量和8个环境变量分别进行典范主坐标分析(Canonical analysis of principal coordinates,CAP)并利用anova函数进行显著性检验,若结果显著则进行下一步。在本研究,两者结果均显著(空间变量:F=6.456,P=0.001;环境变量:F=4.932,P=0.001)。(2)为了减少解释变量,防止整体I型错误的膨胀,采用双重终止原则进行前向选择分析,将全局分析的α显著水平和调整后的R2(R2adj)作为前向选择标准(Blanchet et al.,2008;Astorga et al.,2011),此步骤采用vegan包中的ordiR2step函数基于9999次置换检验进行,筛选后得到的空间变量为 dbMEM1和 dbMEM3,环境变量为最冷月最低温、年均降水和黏土占比。(3)将dbMEM1和dbMEM3组成空间变量组,将最冷月最低温、年均降水和黏土含量组成环境变量组,基于rdacca.hp包的rdacca.hp函数进行变差分解分析,获取空间距离和环境因子的独自效应(unique effect)、共同效应(common effect)和单独效应(individual effect)。此外,单个环境因子的独自效应和单独效应也根据Lai et al.(2022)的方法进行评估。
其中,独自效应指每个变量单独解释的总变量的比例,即与其他解释变量不存在共同解释的部分。单独效应可以简单地估算为解释变量的独自解释部分加上与其他解释变量的共同解释部分的平均分配值,其公式为(Lai et al.,2022):

式中:
IXi——解释变量Xi的单独效应;
p——解释变量的数量;
c——解释变量Xi与其他K个解释变量共享的组合的数量;
S——R2的类型为半偏相关(semi-partial)。
由于多重共线性和校正R2的原因,基于变差分解或层次分割后的组分有可能出现负值的情况,意味着可忽略的贡献,未在图表中显示,此外,置换检验是否显著只有参考的意义,并不能作为解释变量是否重要的依据(Lai et al.,2022),因此本研究中变差分解分析未进行置换检验。
2 结果
2.1 27个国家级自然保护区苔藓植物物种组成
经过名录校对和统计,27个保护区共计苔藓植物130科445属1962种(包含21个亚种和94个变种),占全国总种数的63.66%,其中苔类植物48科106属605种,藓类植物78科335属1350种,角苔植物4科4属7种。包括国家重点保护野生植物 3种,分别是桧叶白发藓(Leucobryum juniperoideum(Brid.) Müll. Hal.)、多纹泥炭藓(Sphagnum multifibrosumX. J. Li & M. Zang)和粗叶泥炭藓(Sphagnum squarrosumCrome),受威胁物种62个,其中极危种(CR)5个,濒危种(EN)18个,易危种(VU)39个。
科含种数排名前 10的科分别是丛藓科(Pottiaceae,包含30属142种,下同)、细鳞苔科(Lejeuneaceae,22,133)、青藓科(Brachytheciaceae,14,90)、灰藓科(Hypnaceae,20,75)、提灯藓科(Mniaceae,10,68)、真藓科(Bryaceae,5,59)、耳叶苔科(Frullaniaceae,1,58)、羽苔科(Plagiochilaceae,3,55)、蔓藓科(Meteoriaceae,19,49)、木灵藓科(Orthotrichaceae,7,48),包含131属777种,分别占总属数的29.44%,总种数的39.60%。属含种数排名前10的属分别是耳叶苔属(Frullania,包含 58种,下同)、羽苔属(Plagiochila,50)、青藓属(Brachythecium,45)、凤尾藓属(Fissidens,41)、疣鳞苔属(Cololejeunea,40)、真藓属(Bryum,40)、光萼苔属(Porella,33)、扁萼苔属(Radula,31)、绢藓属(Entodon,28)、泥炭藓属(Sphagnum,28),包含394种,占总种数的20.08%。
2.2 27个国家级自然保护区苔藓植物地理成分的组成及沿经纬度的分布格局
27个保护区内 1962种苔藓植物可划分为 14种分布区类型,其中世界分布88种,未知分布38种,以温带分布为主,包含486种,占比26.47%(除世界分布和未知分布),其次是东亚分布(包含358,占比19.50%,下同)和热带分布(298,16.23%)。属于温带成分的有592种,占比32.24%,高于热带成分(544,29.63%),此结果与中国种子植物种的地理成分组成一致(吴征镒等,2010)109-114。
27个保护区的 4大地理成分在纬度和经度上的地理格局具体表现为:(1)作为高纬度地区典型代表的温带成分和作为低纬度典型代表的热带成分在纬度和经度上表现出相反的分布格局,温带成分自南向北显著递增(r2=0.835,P=0.001)(图1a),热带成分自南向北显著递减(r2=0.914,P=0.001)(图1c),此结果与云南地区和澜沧江流域(云南段)种子植物以及中国高山带种子植物地理成分的分布格局一致(冯建孟等,2012a,b;沈泽昊等,2017);在经度上,温带成分表现出中间低、东西两边高的分布格局(r2=0.184,P=0.034)(图1b),热带成分则与之相反(r2=0.335,P=0.003)(图1d)。(2)东亚成分在纬度(r2=0.682,P=0.001)上表现出中间高、南北两边低的分布格局(图1e),在经度上自西向东显著递增(r2=0.135,P=0.033)(图1f)。(3)亚洲-澳大利亚-大洋洲成分自南向北显著递减(r2=0.643,P=0.001)(图1g),而在经度上表现出中间高、东西两边低的分布格局(r2=0.268,P=0.009)(图1h)。

图1 27个国家级自然保护区苔藓植物不同地理成分沿纬度和经度的分布格局Figure 1 The distribution patterns of different bryophytes geographical elements along latitude and longitude from 27 national nature reserves
2.3 环境因子和空间距离对27个国家级自然保护区间苔藓植物地理成分分异的影响
基于 CAP模型进行了变差分解分析,结果表明,全局模型解释了 27个国家级自然保护区间苔藓植物地理成分相似性70.50%的变差(图2),环境因子和空间距离的独自效应分别为 18.04%和3.30%,环境因子和空间距离的共同效应为49.15%,环境因子、空间距离的单独效应分别为 42.62%(18.04%+49.15%×1/2)、27.88%(3.30%+49.15%×1/2),表明环境因素对27个保护区间苔藓植物地理成分组成的影响要高于空间距离,此结果与Patiño et al.(2018)的研究结果一致,并且环境因素和空间距离的作用在很大程度上是相互依赖的。环境因子较高的独自效应表明局部微环境对苔藓植物地理成分构成具有较高的影响力。

图2 27个国家级自然保护区苔藓植物地理成分与环境因子和空间因子的变差分解分析Figure 2 Variation partitioning analysis of the geographical elements of bryophytes with environmental and spatial factors from 27 national nature reserves
环境因子中,最冷月最低温拥有最高的单独效应和位于第2位的独自效应(表3),黏土含量的单独效应位于第4位,但拥有最高的独自效应。

表3 27个国家级自然保护区苔藓植物地理成分与各环境因子的变差分解分析Table 3 Variation partitioning analysis of the geographical elements of bryophytes with environmental factors from 27 national nature reserves
3 讨论
3.1 27个国家级自然保护区的苔藓植物区系地理格局
区域植物区系的形成和发展大多与其他地区的植物区系存在不同程度的联系,并且需要经历非常漫长的地质时期(马克平等,1995;吴鹏程等,2001)。本研究中,地理成分以温带成分、东亚成分和热带成分为主,其原因可能与苔藓植物的起源(可能起源于前寒武纪)(Su et al.,2021)和欧洲、北美洲、日本等地区的地质历史有关(Boufford,1998;张宏达,1980;吴鹏程等,2001)。而温带成分高于热带成分的地理成分组成特点可能由以下因素造成:(1)可能与第三纪后期,冰期致使北半球的植物大规模南迁有关(Hewitt,2000;叶建飞等,2012);(2)温带成分的占比一般会随着海拔的增加而逐渐变大(彭华,1996;冯建孟等,2008),而部分保护区的海拔较高(表 1),可能会造成温带成分的比例高于热带成分;(3)温带植物向南拓展分布的能力的总体水平强于热带物种向北拓展分布(彭华,1996),加上27个保护区中,佛坪和喀纳斯等 10个保护区属于温带地区,这可能也是造成温带成分高的原因之一。
由于温带区系植物耐低温,适宜分布在高纬度高海拔区域(冯建孟等,2008),加上我国地形气候特殊,即陆地西部大部分地区属于温带气候,最东部属于高纬度地区。27个保护区中,喀纳斯、西天山等地区位于西部高纬度温带地区,贺兰山、额尔古纳和白石砬子等地区位于中部、东部高纬度地区。因此,27个保护区内苔藓植物的温带成分自南向北显著递增,而热带成分则与之相反。导致东亚成分自南向北呈现中间高两边低的分布格局。亚洲-澳大利亚-大洋洲成分从亚洲的温带区域至热带的澳大利亚地区均有分布(Zhang et al.,2003),而本研究中处于高纬度地区的保护区中该种地理成分占比较少,因此在纬度上呈显著递减。27个保护区的苔藓植物分布区类型在经度上的分布格局可能也与此因素有关,而东亚成分呈现自西向东显著递增的分布格局可能还与日本和韩国等地区与中国的植物区系起源有关,即“扩散假说”认为日本植物区系起源于欧亚大陆,其主要迁移路线途径有库页岛、朝鲜半岛和琉球群岛等陆桥(肖月娥,2014;周徐平等,2022)。
3.2 气候因子、空间距离和黏土含量对27个国家级自然保护区间苔藓植物地理成分相似性的影响
大量研究显示水分和温度是影响苔藓植物分布的重要因素(Möls et al.,2013;Chen et al.,2015b;Qian et al.,2016),而气候因子影响植物的分布通常是通过影响物种特有的温度和水分耐受性生理阈值而进行的,加上苔藓植物结构简单,对气候变化反应敏感,在资源可获得性允许的范围内,苔藓植物能够跟踪不断变化的气候(Walther et al.,2002;Zanatta et al.,2020),而黏土对苔藓植物分布的影响,在一定程度上可能依赖于水分的可利用性,因为细粒土壤具有较高的保水能力(Chamizo et al.,2012;Abay et al.,2014)。因此环境相似的地区,苔藓植物往往拥有更高的区系相似性(物种相似性)(Zhang et al.,2003;Yu et al.,2019),从而形成更相似的地理成分。此结果表明环境因子对27个保护间的苔藓植物地理成分相似性具有重要的影响。
相较于被子植物,苔藓植物较高的长距离扩散能力使它们能够在物种形成后迅速分散,形成苔藓植物较低的地区特有性(Patiño et al.,2018),而物种的扩散过程本身亦可视为空间距离的递减函数(Götzenberger et al.,2012),加上中国是一个多山国家,地势呈阶梯分布,保护区间苔藓植物的远距离扩散可能受到中-高山脉的驱动和阻碍(吴征镒等,2010)1-2,即在一定程度上可能会影响保护区间风的连通性进而对区系相似性产生影响(Munoz et al.,2004)。并且苔藓植物孢子的远距离传播在很大程度上受制于孢子的直径大小(Zanatta et al.,2016)。这些因素导致空间距离较近的地区,苔藓植物往往拥有更高的地理成分相似性,反之则相似性较低。此外,27个保护区间的平均间隔距离高达1482 km(基于保护区几何中心点)。因此,空间因子同样影响 27个保护区间苔藓植物地理成分的构成。
变差分解显示环境因子相较于空间因子拥有更高的独自效应,一方面可能与苔藓植物自身的生长和繁殖特性有关,其体型较小,往往不能远离相关的生态环境,有时只要局部生境适宜,苔藓植物便能生长和繁衍(Patiño et al.,2018;吴鹏程,1998);另一方面可能是苔藓植物拥有较低的空间遗传结构(Patiño et al.,2018)。本研究中,空间因子的独自效应占比了3.30%的贡献,这一部分空间独自效应可能存在一定的不确定性(Anderson et al.,2011;Qiao et al.,2015),即可能是由于具有生态生理重要性的环境因子往往在自然扩散范围内趋于空间自相关(Nekola et al.,1999;Capinha et al.,2015),此过程可能包含本研究中未被纳入的环境因子。
最冷月最低温拥有最高的单独效应和位于第 2位的独自效应,可能是由于温度是限制热带成分和温带成分相对比重的重要因素(彭华,1996;冯建孟等,2012b),而保护区间的最冷月最低温的平均间隔差异最大(基于欧几里得距离),并且亚洲-澳大利亚-大洋洲成分和东亚成分分别与温带成分和热带成分之间存在不同程度的相关性(表 4)。黏土含量拥有最高的独自效应,可能是由于土壤中存在本研究中未纳入的潜在的Na、P和Mg等化学性质对苔藓植物分布的影响(Ma et al.,2020)。
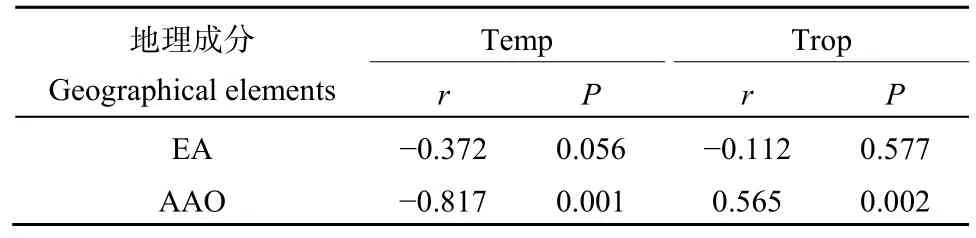
表4 温带成分和热带成分与东亚成分和亚洲-澳大利亚-大洋洲成分间的pearson相关Table 4 Pearson coefficients between Temperate elements,Tropical elements and East Asian elements, Asia-Australia-Oceania elements
总之,本研究结果表明 27个国家级自然保护区间苔藓植物地理成分的相似性是由环境和空间因子共同决定,并且环境因子的贡献要高于空间因子。另外,需要强调的是,分布区的形成除了与种的适应能力和适应生态的范围密切相关,即生态原因以外,历史原因,即种的历史形成过程和形成中的环境条件也是影响分布区形成的重要因素(吴征镒等,200640-41;吴征镒等,2010109-120)。而本研究更多的是从空间距离和现代环境因素影响区系相似性(物种相似性)的角度出发揭示27个保护区间苔藓植物地理成分的相似性,各保护区间地理成分的相似性可能还与各保护区区系形成的历史原因有关(Zhu,2013,2016;Patiño et al.,2018;吴鹏程,1998),值得进一步探讨。
4 结论
通过对 27个保护区的苔藓植物物种进行校对和统计发现,27个保护区物种组成丰富,共计苔藓植物130科445属1962种,有国家重点保护野生植物3种,受威胁物种62个;27个保护区的苔藓植物分布区类型以温带成分为主,随着纬度的增加(自南向北),温带成分显著增加,热带成分和亚洲-澳大利亚-大洋洲成分均显著减少,东亚成分在中部达到最大值;随着经度的增加(自西向东),东亚成分显著增加,温带成分在中部处于最低值,而热带成分和亚洲-澳大利亚-大洋洲成分在中部达到最大值;环境因子和空间距离共同驱动27个自然保护区间的地理成分分异,并且环境因子的驱动作用要强于空间因子。研究结果基于保护区间的空间距离和现代环境因素,初步揭示了影响27个保护区间苔藓植物地理成分相似性的因素,为苔藓植物区系研究提供了一定的理论基础。
致谢:感谢深圳市中国科学院仙湖植物园张力研究员在文章撰写过程中提供的宝贵意见和建议。
