Late Triassic flora of the northern part of the Fore-Caspian depression (Kazakhstan) as exemplified by data from the S. Nurzhanov-509 borehole
2022-12-07NaugolnykhandPronin
S. V. Naugolnykh and A. P. Pronin
1. Geological Institute, Russian Academy of Sciences, Moscow 119017, Russia;
2. Company Zhakhan, Atyrau, Kazakhstan
Abstract: The Late Triassic Nurzhanov flora is characterized on the basis of the material originated from the S.Nurzhanov-509 borehole disposed in the northern part of the Fore-Caspian depression, near the City of Atyrau, Republic of Kazakhstan. Taxonomic composition of the Nurzhanov flora includes equisetophytes Neocalamites sp., ferns Phlebopteris hazarensis Naugolnykh et Pronin, Todites sp., Cladophlebis sp., ginkgophytes Ginkgoites spp., conifers Podozamites sp. Palaeophytogeographic status of the Nurzhanov flora is discussed. According to the data obtained, this flora belongs to a transitional zone (an ecotonous belt), which existed between the Euro-Sinian and the Siberian-Canadian palaeofloristic realms during most of the Mesozoic time.
Keywords: Triassic; palaeobotany; equisetophytes; ferns; conifers; palaeophytogeography
Introduction
Triassic period was the time when the initial developing of the Mesozoic-type ecosystems took place practically around the world. These ecosystems were characterized by high dominating large diapsid reptiles (mostly, dinosaurs and their relatives) in the vertebrate faunas of the terrestrial environments,and by the wide distribution of land vegetation represented by numerous and various gymnosperms belonging to the typically Mesozoic groups, such as ginkgophytes, czekanowskians, bennettitids,cycadophytes s.s., and the Mesozoic families of conifers. Appearance and further developing of the new groups of leptosporangiate ferns (for instance, the family Matoniaceae) were characteristic of the Early Mesozoic floras, as well. The protoleptosporangiate ferns of the family Osmundaceae, which originated in the middle part of Permian period (Esaulova, 1996;Naugolnykh, 2002), reached their real flourishing peak in the Late Triassic and Jurassic (Shorokhova, 1975;Schweitzeret al.,1997; Phippset al.,1998; Anderson& Anderson, 2008).
Localities of the upper Triassic floras, which are characterized by both good preservation allowing to study the cuticular anatomy, and by high taxonomic diversity, are relatively rare. The most well-studied Middle- and Upper Triassic floras of the world are the Keuper flora of the West- and Central Europe(Grauvogel-Stamm, 1978; Kelber & Hansch, 1995),the Molteno flora of Southern Africa (Anderson,Anderson, 2003, 2008, etc), the Madygen flora of Middle Asia (Sikstel, 1962; Dobruskina, 1980, 1982),see these two works for further references on the Triassic flora of Eurasia, the flora of Timan-Pechora region (Khramova & Pavlov, 1971; Khramova, 1977;Kirichkova & Esenina, 2015), and Far-East (Shorokhova, 1975; Sun, 1979, 1981, 1990, 1993; Volynetzet al.,2008).
Any discovery of every new locality of the Late Triassic plant megafossils can be considered as a notable event in palaeobotany, especially if such locality is characterized by the plant remains of good preservation, which allows to study their epidermal-cuticular structure, and if the plant macrofossils are accompanied by dispersed cuticles, spores and pollen. The present paper is devoted to the generalized characteristics of the recently discovered (Naugolnykh & Pronin, 2015)Late Triassic flora of the northern-western part of the Republic of Kazakhstan, named as “Nurzhanov flora”after the locality S. Nurzhanov-509 (see details below).
1 Materials and methods
The collection studied originates from the “S.Nurzhanov-509” borehole (Figs.1A, B; 2), which is disposed in the northern-western area of the Republic of Kazakhstan, Atyrau region, Zhylyorskian district;in eastern part of the Fore-Caspian depression. The plant megafossils and a sample for bulk-maceration by means of the “Three filters” approach were collected from the depth 3223 m.
The fossil plants are preserved as compressions and impressions (Figs.3-9). Products of bulk-maceration (Figs.10-12), including isolated cuticles, tracheids,sporangia, spores and pollen, were obtained by means of the “Three filters” approach (Naugolnykh, 2015).
The micro- and mesofossils were studied in SEM (Vega Tescan MV 2300 scanning electronic microscope) and in the digital optic microscope Meiji Techno (MT 9930) at the Geological Institute (GIN)of Russian Academy of Sciences, Moscow. The collection studied is kept at GIN under number 4851.
2 Brief stratigraphical review
Age of the deposits under study is determined as Late Triassic mostly by the palynological data. The part of the section characterized by the plant megafossils should be assigned to the Sankudukian Horizon,which corresponds to the Aschitaipakskian Series of the Upper Triassic sequences of the so-called “Prorvatype”, see for the further discussion (Yanshin, 1936;Sokolova, 1958; Lipatova, 1982, 1985; Kukhtinovet al.,1980; Kukhtinov & Crasquin-Soleau, 1999; Proninet al.,2012). The Prorva palynological assemblage includes following taxa of palynomorphs: microsporesAratrisporitessp., belonging to a heterosporous lycopodiophyteTomiostrobusNeuburg: see for discussion (Yaroshenko, 1988; Naugolnykh, 2012);sporesCyathidites triangularisRom., presumably belonging to the fernDictyophyllum nilssonii(Brongniart) Goeppert emend. Kruch.; some other palynomorph taxa:Duplexisporites gyratesPlayford et Dettmann,Kyrtomisporites speciosusMädler,K.laevigatusMädler,Stereisporitessp.,Ovalipollissp.,Gnetaceapollenitessp.,Ginkgocycadophytussp.,Sulcatisporitessp.,Alisporitessp.,Chordasporites singulichordaKl. (lower part of the upper Triassic);Zebrasporites interscriptus(Thiergart) Klaus,Cingulizonatesspp.,Kyrtomisporites speciosusMädler,Triancoraesporites ancorae(Reinch.)E. Schulz,Lycopodiacidites rhaeticusE. Schulz,C. seebergensisE. Schulz,Conbaculatisporites longdonensisClarke, etc), and plant megafossilsNeocalamites carcinoidesHarris,Clathropterissp.,Podozamites lanceolatus(Lindley et Hutton) Braun,P. angustifolius(Eichwald) Heer,Glossophyllum(?)sp.,Sphenobaierasp., the assemblage was collected from the Upper part of the upper Triassic of the Prorva section, see for details (Lipatova, 1982, 1985).
Distribution of the Triassic deposits of the Fore-Caspian depression is mostly traced by the data obtained after regional bore-hole projects. Most of the bore-holes are disposed at the central part of the large but local salt diapirs (so called “salt domes”)located mostly in the southern part of the Fore-Caspian depression, since these diapirs are here located just near the earth “day surface. The Upper Triassic deposits are covering with the discontinuity and partial rewashing the older deposits, mostly Lower Permian,because the Lower Triassic and Middle- and Upper Permian deposits are absent here. Average thickness of the Upper Triassic deposits in these sections is about one hundred meters.
The Upper Triassic deposits of the region studied can be divided into three lithologically distinct formations: (1) lower formation of the sand-pebbles deposits; (2) middle formation of the motley-colored clay; (3) upper formation of the green-gray clay and sandstones. There are some variations in lithology of these formations regarding local geographical peculiarities, but in general all these three formations can be easily traced through the whole region studied.The borehole S. Nurzhanov-509 belongs to the lower sand-pebble formation (Fig.2A). The plant megafossils occur mostly in the lenses of the fine-grained graygreenish sandstones and siltstones.

Fig.2 Correlation chart of the Upper Triassic deposits of the Fore-Caspian depression.
3 Taxonomic composition of the Nurzhanov flora
The taxonomical composition of the Nurzhanov flora includes all the major groups of higher plants,which are characteristic of the Mesozoic vegetation around the world, i.e. equistophytes, ferns, ginkgophytes, and conifers. Presence of cycadophytes in the Nurzhanov flora is not proved yet, although there is some indirect evidence (dispersed pollen and cuticles of putative cycadophyte affinity), which shows that the cycadophytes s.l. could be present as accessory elements in the initial plant communities. A brief description of the main plant groups of the Nurzhanov flora is given below.
3.1 Equisetophytes
The equisetophytes of the collection studied are represented by the fragments of large segmented stems with the fine and narrow prolonged ribs, and with the distinct well-pronounced nodes (Fig.3A, G). Width of the stems reaches 6 cm. On the basis of general extrapolation of the width and average speed of decreasing of the stem width typical of equisetophytes(Milde, 1851; Meuselet al.,1971), one can suggest that the parent plant was as tall as at least 3 m or even more. Width of the stem ribs is about 1 mm. There is a small rib of second order, which is disposed in axial/medial part of each rib of the first order. The ribs are alternating in the stem node. Such a pattern of the rib position is characteristic of both recentEquisetumL.and of the fossil representatives of the family Equisetaceae. The node bears small leaf scars. These leaf scars are of round to ovoid shape, slightly prolonged along the stem axis, with the well-pronounced margins,which are slightly uplifted above the stem surface, and with the small depression at the central part of the scar.Diameter of the leaf scar is about 0.7 mm. The leaf scars are disposed in more or less equal distance from each other. This distance is approximately 10 mm. Each couple of the leaf scars is segregated by three to four stem ribs located between the scars. As far as we can judge from the shape of the leaf scars, the leaves themselves were narrow, linear and most probably relatively long. The leaves were not fused by their margins and didn’t form the leaf sheaths. According to all the characters listed above, these equisetophytes of the Nurzhanov flora can be attributed to the genusNeocalamitesHalle, discussion of the generic status ofNeocalamtessee in Naugolnykh, 2009; Yanget al.,2011. The most similar species isNeocalamites merianii(Brongniart) Halle( Kelber, 1990; Kelber &Hansch, 1995). The equisetophytes of the Nurzhanov flora most likely grew in wet near-water habitats,as it was typical for this plant group since the Late Paleozoic (Isaevet al.,2018).

Fig.3 Nurzhanov flora
3.2 Ferns
The Nurzhanov flora includes some representatives of the fern families Matoniaceae and Osmundaceae. The matoniacean ferns are represented byPhlebopteris hazarensisNaugolnykh et Pronin (Naugolnykh& Pronin, 2015). This species is characterized by structures of the fertile leaves (Fig.4F, G), sterile leaves(Fig.5A, B), sori (Figs. 4D; 5D), sporangia (Fig.4A,I), and the spores preservedin situ(Figs. 4B, C, E, H;5C).

Fig.4 Matoniacean fern Phlebopteris hazarensis Naugolnykh et Pronin
The osmundaceaen ferns in the collection studied are represented by the sterile leavesCladophlebissp.(Fig.3, D) and by the isolated spores ofTodisporites–type (Fig.3C). Spores of this type are characteristic of the osmundaceaen ferns, for discussion see Balme,1995; Naugolnykh, 2002. An isolated sporangium of long-ovoid shape with the group of the thick-walled cells located around the apical stomium (Figs.10H; 11,H) most probably also belongs to an osmundaceaen fern.
There are several dispersed cuticles, which are imaged here (Figs.10C, D, J; 11C, D, J). They have many features in common with the cuticles of some of the recent leptosporangiate ferns, i.e.Pteridium aquilinum(L.) Kuhn, compare, for instance, the cuticles figured here: Figs.10C, D, J; 11C, D, J, and the cuticles figured in Naugolnykh, 2017, Fig.5.

Fig.5 Matoniacean fern Phlebopteris hazarensis Naugolnykh et Pronin
3.3 Ginkgophytes
The ginkgophytes of the Nurzhanov flora are represented by the leaves belonging to two distinctly different morphotypes. Leaves of the first morphotype,which is conditionally named here in open nomenclature asGinkgoitessp.1 are most numerous. These leaves are of classicGinkgo-type morphology. The leaves are relatively wide, with the wedge-like base,deep central sinus and additional sinuses of dissection of second and third orders (Figs. 3H; 6E). The resulting lobes of second and third orders are widecuneate with round to spatulate apices (Figs. 7A, B,D, G; 8A, B, D, K). The morphotypeGinkgoitessp.1 is represented in the collection in hand by both almost complete leaves, and their fragments. Taphonomically the leavesGinkgoitessp.1 often form dense mass aggregations directly reflecting initial thanatocoenoses.Several preparations of the cuticle were made from the leaves ofGinkgoitessp.1 (Figs. 6A-G; 7A-G;8A-L; 9A-J). Numerous round to ovoid resin bodies are well-observed inside the leaf mesophyllous tissues (Figs. 6A, B, D, F, G; 7A-D, F, G; 8A-G, J,K; 9A, C). Cell walls of the lower cuticle of the leaf are straight to slightly curved. Epidermal cells are prolonged, pentagonal to hexagonal (Figs.6C; 9H),sometimes rectagonal/quadrigonal. Average size of the common epidermal cells is 20×70 μm. The stomata are unsunken, with the lip-like cutinization of the guard cells (Figs.8H, L; 9F, I). Numerous monocolpate pollen ofCycadopitys(=Entylissa) type are preserved on the cuticle surface (Figs.8I; 9D, E, G). Obviously,these pollen grains belonged to or were produced by the same parent plant with the leavesGinkgoitessp.1.Similar pollen aggregations are well-known for other fossil Mesozoic ginkgophytes (Harriset al., 1974).

Fig.6 Ginkgophytes of the Nurzhanov flora; Gingoites sp.1

Fig.7 Ginkgophytes of the Nurzhanov flora; Gingoites sp.1

Fig.8 Ginkgophytes of the Nurzhanov flora; Gingoites sp.1
Leaves of the second morphotype, which is conditionally named here asGinkgoitessp.2, occur not that often asGinkgoitessp.1. This morphotype is represented by a single although practically complete leaf (Fig.3E) and several less distinct fragments. The leaves of this morphotype (in contrast toGinkgoitessp.1) have much longer and narrower segments(lobes), and this character makes them similar to some representatives of the genusBaieraBraun (Shorokhova et al., 2009, Plate C, Figs.1–7). Ginkgophyte leaves of the same morphology are known from a number of Permian and Triassic floras (Anderson &Anderson,2003; Naugolnykh, 2018).
3.4 Conifers
Conifers of the Nurzhanov flora include the genusPodozamitesBraun (Fig.3B, F) and dispersed bisaccate pollen (Fig.12A, B, E, F, I), which most probably belonged to the same parent plant. Similar bisaccate pollen are typical of many Permian and Triassic conifers, mostly of Voltzialean affinity (see discussion in: Naugolnykh, 2014). The leavesPodozamitesof the Nurzhanov flora are lanceolate, reach 5–6 cm length and 0.8 cm width, with the distinct parallelodrome venation. Apex of the leaf can often be truncate(Fig.3B) or sometimes can bear a small terminal sinus.Base of the leaf is cuneate (Fig.3F).

Fig.1 Map of the geographical position of the localities studied (A) and litho-facial map of the Upper Triassic deposits of the Fore-Caspian depression (B)
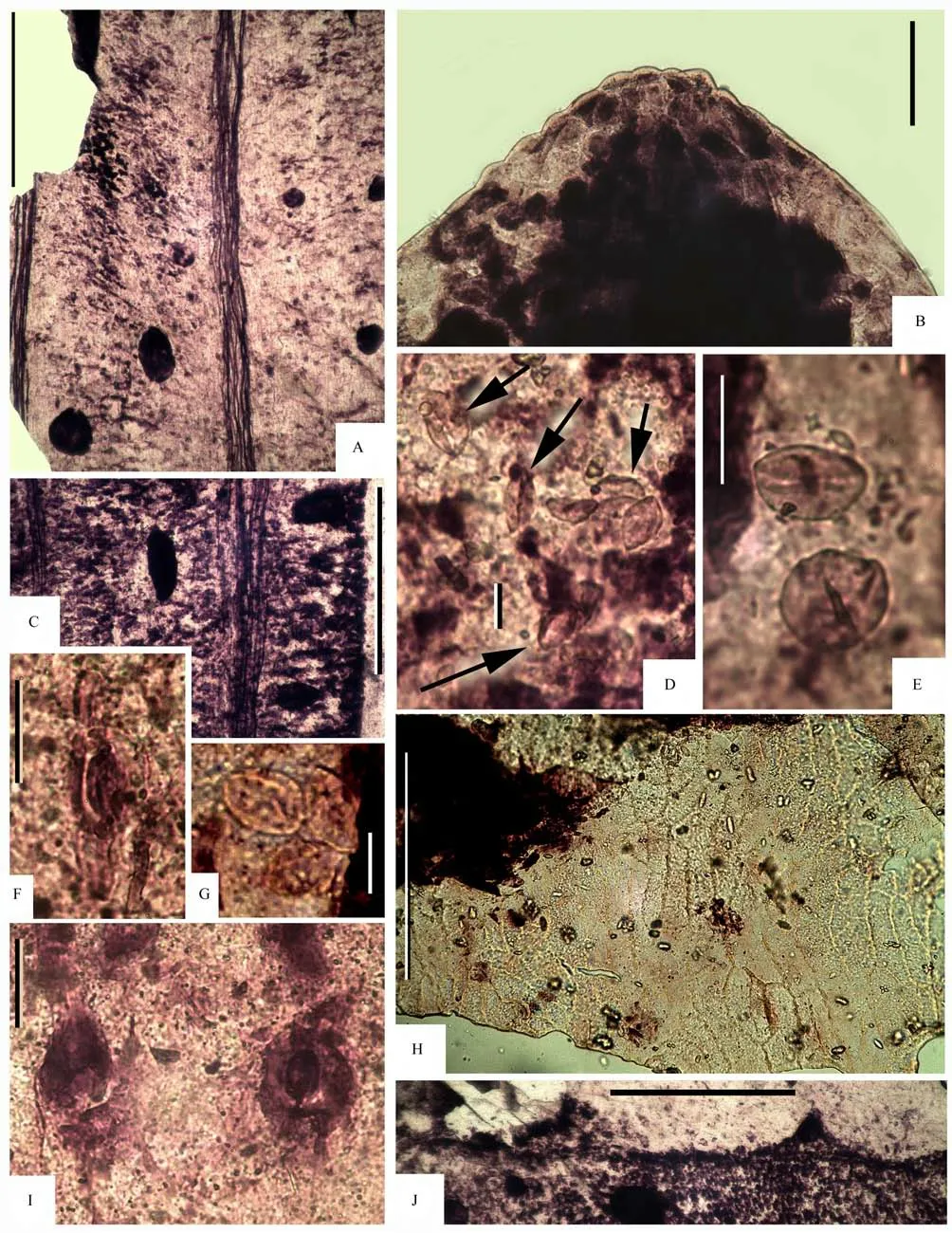
Fig.9 Ginkgophytes of the Nurzhanov flora; Gingoites sp.1, cuticular structure
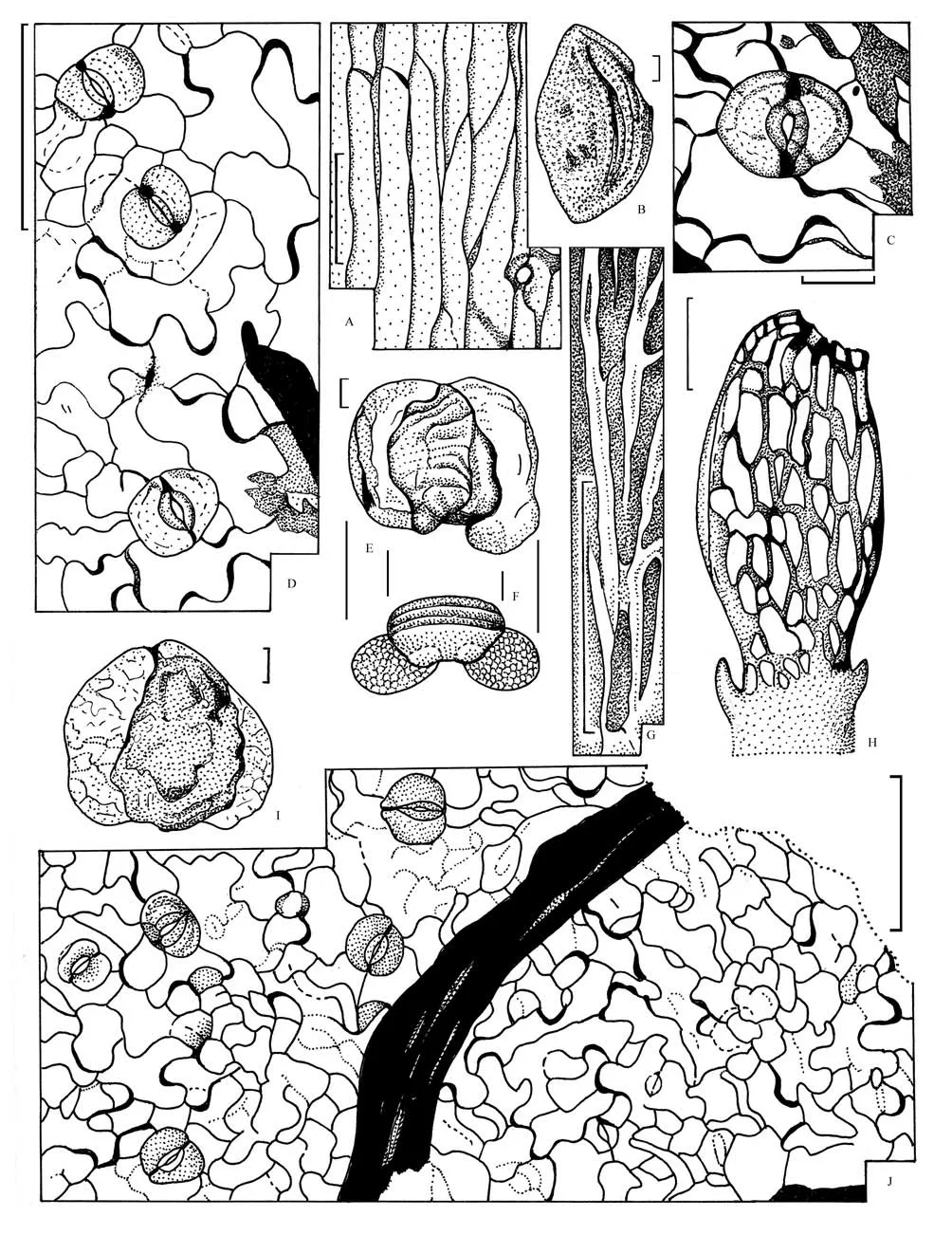
Fig.11 Nurzhanov flora (Palynomorphs and isolated cuticles obtained as a result of bulk-maceration by “Three Filters” approach)

Fig.12 The Nurzhanov flora (Palynomorphs, (A)-(F), (H) and (I); isolated tracheid, (G), obtained as a result of bulk-maceration bymeans of the “Three Filters” approach)
Besides of the plant remains, which are briefly characterized above, the Nurzhanov flora includes numerous palynomorphs, fragments of conducting tissues (mostly, tracheids), isolated cuticles of several types and other micro- and mesofossils, exact taxonomic attribution of which is still unclear.
To sum up the data listed above, the current list of the plant taxa of the Nurzhanov flora includes equisetophytesNeocalamitessp., osmundacean fernsToditessp.,Todisporitessp.,Cladophlebissp., matoniacean fernPhlebopteris hazarensisNaugolnykh et Pronin,ginkgophytesGinkgoitessp.1,Ginkgoitessp.2,conifersPodozamitessp. It is quite possible that the Nurzhanov flora includes also the heterosporous lycopodiopsids of the family Isoetaceae, since there are several microspores of the genusAratrisporitesLeschik (Fig.10B), discussion of the taxonomic position ofAratrisporitessee in Yaroshenko, 1988 and Naugolnykh, 2012.

Fig.10 Nurzhanov flora (Palynomorphs and isolated cuticles obtained as a result of bulk-maceration by “Three Filters” approach)
4 Paleogeographic conclusions
The Late Triassic phase of sedimentation in the Fore-Caspian depression is characterized (in contrast to the middle Triassic) by the general peneplainization of the relief through out of all this region and by the contemporaneous weak effects of adding of the terrestrial clastic particles from the local upliftings or even higher elevations of the Paleo-Urals, and appearing of the salt diapirs. According to the general geologic data (Zhidovinov, 1994), the conditions of sedimentation on the Fore-Caspian depression in the late Triassic time were continental. A wide alluvial plain with the well-developed net of the rivers and lakes existed on this territory during all the late Triassic. Some temporal water channels took place in the eastern part of the plain. Sedimentation on all the plain was totally terrigenous, and was characterized by cyclic intercalation of the alluvial s.s., limnic,and swamp deposition enriched by clastic material transported from the northern part of the Fore-Caspian depression and from southern margin of the Palaeo-Urals. Presence of the fine-grained siderite in these deposits supports a conclusion that the terrestrial palaeoenvironments of the Fore-Caspian depression had the humide paleoclymatic conditions. According to the authors viewpoint, the plant communities of the Nurzhanov flora were clearly divided into hydro- and hygrophilous associations inhabited by the equisetophytesNeocalamitesand possibly by the ferns, and mesophilous subplacor associations with the dominating ginkgophytesGinkgoitesspp. and conifersPodozamites. In palaeogeographic terms these plant associations were adapted for different parts of the palaeoenvironments/palaeolandscapes, and only as a taphonomic mixture were preserved together in one and the same thanatocoenosis. Humidity of the climate was resulted in a flourishing of the terrestrial vegetation on the Fore-Caspian depression in the Late Triassic time, with the exception of the areas where the salt diapirs were exposed on the earth “day surface”and where the taxonomically poor plant communities were influenced by the hyper-salt water.
5 Florogenetic conclusions
On the one hand, a general type of the Nurzhanov flora, as well as the presence of the matoniacean ferns and large equisetophytes clearly point to the conclusion, that this flora was thermophile, welladapted for warm and wet tropical climate, as it was assumed for many other low-latitude fern-containing Late Paleozoic and Mesozoic floras (Yanget al.,1987;Wang & Mei, 1999; Wang, 2002). But, on the other hand, the local dominating of the ginkgophytes and absence of numerous cycadophytes both show that this flora has a similarity with the boreal Siberian-Canadian floras (Vakhrameev, 1990). According to our point of view, the Nurzhanov flora was disposed in the ecotonous zone between the Euro–Sinian and Siberian-Canadian palaeophytogeographic realms.Florogenetically this ecotonous zone (belt) partly repeated a position of the Subangaran ecotonous zone of the late Palaeozoic, which was located between Euramerian (on south-west) and Angaran (on northeast) palaeophytogeographic realms (Meyen, 1987).Similar Triassic floras existed in neighboring areas of Kazakhstan and were reported and described earlier(Brick, 1952). In general, such ecotonous zones could serve as effective gateways for inter-realm migrations of the higher plants both in Late Paleozoic and Early Mesozoic (Levenet al., 2011).
Acknowledgements
This work was funded by the State Program of the Geological Institute of Russian Academy of Sciences,Moscow.
杂志排行

Global Geology的其它文章
- Glossopterids survived end-Permian mass extinction in North Hemisphere
- The first occurrence of Scincomorpha lizard from Nenjiang Formation (lower Campanian) of Jilin, Northeast China
- Characteristics of source rocks and paleoenvironment of Lower Silurian Longmaxi Formation in western Hubei
- Characterization of mineral element fingerprints under soilcrop systems in eastern and western rice-producing areas of Jilin Province, China
- Unveiling Ceremony of Academician Li Xingxue’s Former Residence Scientists’ Spiritual Education Base held in Chenzhou of Hunan
- CONTENTS