“通督调脏”针法对CUMS大鼠行为学及海马组织蛋白表达的影响
2022-12-06金红齐泰煜樊帅李光英钱家坤谢雨虹张琪高美云高敏香张敏
金红 齐泰煜 樊帅 李光英 钱家坤 谢雨虹 张琪 高美云 高敏香 张敏,4
(长春中医药大学 1中医学院,吉林 长春 130017;2针灸推拿学院;3附属医院脑病中心神志门诊;4附属医院针灸临床中心)
抑郁症以显著而持久的心境低落为临床特征,为疾病负担最重的精神疾病〔1〕。目前西药抗抑郁谱窄、易复发等缺陷〔2〕、抗抑郁药物研发进展缓慢。督脉和足太阳经被选择频率最高〔3〕,但在辨证治疗上经络辨证和脏腑辨证分庭抗礼。通督调脏针法源自《黄帝内经》中督主神志、五神脏理论和长白山通经调脏手法流派的经络-脏腑相关理论,以百会、印堂、五脏俞为主穴,以脏腑-经络结合辨证为特点,前期研究发现其能改善抑郁、焦虑等核心及周边症状,起效速度快于五羟色胺再摄取抑制剂(SSRIs)药物〔4〕,并在实验中明确了百会、印堂的抗抑郁疗效和作用机制〔5〕,但还需利用现代生物工程技术对通督调脏针法进行深入研究。
定量蛋白质组学为分子分析、生物标志物识别、探索发病机制的重要方法,其整体性、系统性、综合性与中医脏腑-经络辨证相符,应用4D Label free定量蛋白质组学检测技术发现针刺可通过保护海马突触和线粒体功能改善发挥抗抑郁作用〔6〕,相关差异蛋白可能为疾病诊断和治疗的新靶点〔7〕。但目前尚无督脉联合五脏俞抗抑郁的蛋白质组学研究。慢性不可预见性应激抑郁模型(CUMS)在行为学表现、生化及神经病理改变上与临床变化高度相似,为最有效和最常用的抑郁模型〔8〕。海马为情绪产生和调节的重要脑区,其神经元丢失、蛋白改变被认为与抑郁症关系密切〔9〕。本研究探讨通督调脏针法对CUMS大鼠海马组织蛋白表达的影响及可能的作用机制。
1 材料与方法
1.1实验动物及分组 由长春中医药大学动物实验中心提供,健康SPF级SD雄性大鼠(4周龄)27只,体重180~220 g,饲养温度23~25℃,湿度50%~60%,自然节律光照,自由进食进水,适应饲养1 w,饲养环境清洁。该实验已通过长春中医药大学实验动物伦理委员会审查(批准编号:2020336),全程遵守《关于善待实验动物的指导性意见》中相关规定。将动物随机分为正常组、模型组、针刺组,每组9只,进行行为学数据分析,各组随机选取3只进行蛋白质组学检测。
1.2主要试剂和仪器 蛋白酶抑制剂(德国Merck Millipore)、胰蛋白酶(美国Promega)、二硫苏糖醇、碘乙酰胺、甲酸(美国Fluka)、尿素(美国Sigma-Aldrich)、二喹啉甲酸(BCA)蛋白检测试剂盒(上海碧云天生物技术有限公司)。乙腈、Bruker timsTOF Pro质谱仪(德国Bruker)、Orbitrap ExplorisTM480高分辨质谱仪EASY-nLC 1 200超高效液相系统、Q Exactive系统(美国ThermoFisher Scientific)。
1.3针具和其他材料 一次性不锈钢毫针(规格:0.25 mm×25 mm,贵州安迪药械有限公司)、无菌棉签、75%消毒酒精。其他器材:离心管、医用橡皮管、自制游泳池(61 cm×35.5 cm×43 cm)、华佗牌电针仪(SDZ-II,苏州医疗用品厂有限公司)等。
1.4大鼠CUMS模型制备 造模方法参照Willner等〔10〕方法建立CUMS抑郁模型,造模各组孤养并每日随机给予以下刺激的一种:断食24 h、断水24 h、昼夜颠倒24 h、夹尾3 min、束缚3 h、10℃冷水游泳5 min、潮湿垫料24 h,共21 d。
1.5各组干预方法 将SD大鼠随机分成空白组、模型组、针刺组,每组9只。空白组正常群养,每日给予等量纯净水;模型组慢性不可预见性应激结合孤养,每日给予等量纯净水。针刺组慢性不可预见性应激结合孤养,于造模24 h后开始“通督调脏”针法治疗,穴位参照全国针灸学会实验针灸研究会制定的实验动物针灸穴位图谱,选取相当于人体解剖部位的百会、印堂、心俞、肝俞、脾俞、肺俞、肾俞。大鼠固定于鼠台上,取穴消毒后毫针平刺百会、印堂7.5 ~12.5 mm,施平补平泻法,选用华佗牌SDZ-Ⅱ型电子针治疗仪,连接百会、印堂,疏密波,频率2 Hz/15 Hz,强度为 0.1~1.0 mA,电流强度以针身微微颤动为度;针刺双侧心俞、肝俞、脾俞、肺俞、肾俞,斜刺7.5~12.5 mm,留针30 min,中间提插捻转1次,1次/d,7 d为1个疗程,共3个疗程。
1.6行为学干预
1.6.1强迫游泳实验 游泳池为长方形水池,长61 cm,高35.5 cm,宽43 cm,水深25 cm,水温控制在(25±1)℃。测试时捏住大鼠尾巴距根部2/3 处将大鼠缓慢放入游泳池内,测定大鼠5 min内四爪均不动行为的累计时间,每只大鼠测定1次,在实验第1天、第21天进行测试。
1.6.2糖水消耗实验 大鼠断水24 h,测1 h内蔗糖水摄入量(即测试前后瓶中水量之差)。给予1%蔗糖水,糖水与瓶的重量共计250 g,计算大鼠1 h饮用前后的总重量之差。分别在实验的第1天和第21天测定。
1.7蛋白质组学检测
1.7.1海马组织蛋白提取 21 d后,大鼠断颈处死,冰台上迅速分离出海马组织,记录编号后液氮速冻,-80℃冰箱保存。各组选取3个有明显表型变化大鼠的海马样本,至液氮预冷的研钵中,加液氮充分研磨至粉末。各组样品分别加入粉末4倍体积裂解缓冲液(8 mol/L尿素,1%蛋白酶抑制剂),超声裂解。4℃,12 000 r/min离心10 min,去除细胞碎片,上清液转移至新的离心管,利用BCA试剂盒进行蛋白浓度测定。
1.7.2胰酶酶解 各样品蛋白取等量进行酶解,用裂解液将体积调整至一致。缓慢加入终浓度20%三氯乙酸(TCA),涡旋混匀,4℃沉淀2 h。4 500 r/min,离心5 min,弃上清,用预冷的丙酮洗涤沉淀2~3次。晾干沉淀后加入终浓度200 mmol/L的四乙基溴化铵(TEAB),超声打散沉淀,以1∶50的比例加入胰蛋白酶,酶解过夜。加入二硫苏糖醇(DTT)使其终浓度为5 mmol/L,56℃还原30 min。之后加入碘乙酰胺(IAA)使其终浓度为11 mmol/L,室温避光孵育15 min。
1.7.3液相色谱-质谱串联分析 肽段用液相色谱流动相A相溶解后使用EASY-nLC 1 200超高效液相系统进行分离。肽段经由超高效液相系统分离后被注入Capillary离子源中进行电离然后进Orbitrap ExplorisTM480质谱进行分析。离子源电压设置为1.6 kV,肽段母离子及其二级碎片都使用高分辨的TOF进行检测和分析。二级质谱扫描范围设置为100~1 700。数据采集模式使用平行累积串行碎裂(PASEF)模式。一张一级质谱采集后进行10次PASEF模式采集母离子电荷数在0~5范围内的二级谱图,串联质谱扫描的动态排除时间设置为20 s避免母离子的重复扫描。
1.8统计学方法
1.8.1行为学 采用SSPS26.0软件,组间比较符合方差齐性检验采用单因素方差分析,方差不齐采用H检验;组内比较差值符合正态性分布采用配对样本t检验,偏态分布采用符号秩和检验。
1.8.2蛋白鉴定和定量 通过质谱分析得到样本的肽段质量和信号强度(一级谱)、肽段碎裂后碎片离子的质量和信号强度(二级谱),使用Proteome Discoverer(v2.4.1.15)数据库将蛋白序列构建的理论二级谱图数与质谱分析得到的二级谱图数比对,共鉴定到5 913个蛋白质,根据蛋白特异性肽段和蛋白质数总体情况及肽段在不同样本中的信号强度值等信息计算蛋白相对定量值,使用Pearson相关性统计方法进行评估蛋白定量的重复性。
1.8.3差异蛋白筛选和生物信息学分析 将每个蛋白在两个比较样品中的相对定量值进行t检验,差异蛋白表达量变化超过1.5倍和小于1/1.5作为显著上调和下调的变化阈值。筛选出的差异蛋白采用基因本论(GO)、KEGG通路和亚细胞定位等方法进行功能注释和富集、聚类分析,3组差异蛋白富集分析使用Fisher精确检验,以观察差异表达蛋白在不同功能类型中的富集趋势和功能的相关性;根据数据库编号或蛋白序列,通过STRING(v.11.0)蛋白网络互作数据库观察差异蛋白互作关系。
2 结 果
2.1行为学结果 各组体重均有上升,7 d时模型组体重明显低于空白组(P<0.000 1),模型组与针刺组相比体重变化不明显;与模型组相比,针刺组在治疗14 d时体重显著上升(P<0.001),并在21 d时具有极显著差异(P<0.000 1),但与空白组相比仍具有极显著差异(P<0.000 1)。强迫游泳实验中,21 d时,与空白组相比,模型组不动时间明显增加(P<0.000 1);针刺组不动时间明显低于模型组(P<0.000 1),但与空白组相比仍具有显著差异(P<0.01)。糖水消耗实验中,21 d时,空白组、针刺组糖水消耗量明显增加,模型组变化不明显;模型组糖水消耗量显著低于空白组(P<0.000 1),针刺组糖水消耗量显著多于模型组(P<0.000 1),但与空白组相比仍具有显著差异(P<0.01),见表1。
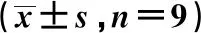
表1 各组体重变化、强迫游泳实验及糖水消耗实验的比较
2.2生物学重复检验 随机从空白组、模型组、针刺组中各选取3个样本,共9个样本进行3组3次生物学重复实验,3组Pearson系数经3次重复后数值均>0.950。见图1。
Pearson系数越接近-1为负相关,越接近1为正相关,越接近0为正相关
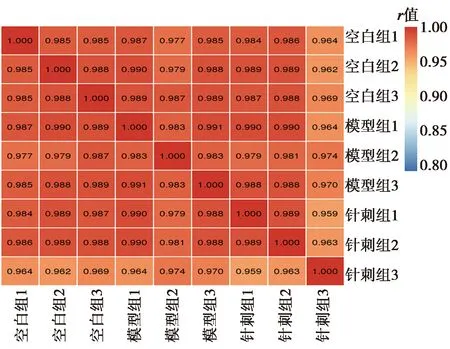
2.3差异蛋白鉴定 共鉴定到39个差异蛋白(阈值为1.5倍),上调蛋白19个,下调蛋白20个;其中模型组相比空白组有10个上调蛋白(Mrps24、Rdh13、Bbs2、Tex2、Dmxl1、Snap29、Rlbp1、Afg1l、Cpped1、Cbx3),12个下调蛋白(Cds1、Ighm、Arhgef6、Vtn、Slc12a9、Slco1c1、Cyp51a1、Plekhd1、Cib1、Pdf、Zfp512、C1qc);针刺组相比模型组有9个上调蛋白(Clcc1、Sdf2、rCG_52671、Pdf、Tmem254、Gpr37、Scd2、Ttyh2、Ddx46),8个下调蛋白(Coq8a、Ik、Specc1l、Phf5a、Ptn、Prpf40a、Bloc1s1、Cdk5rap3),见图2、表2。
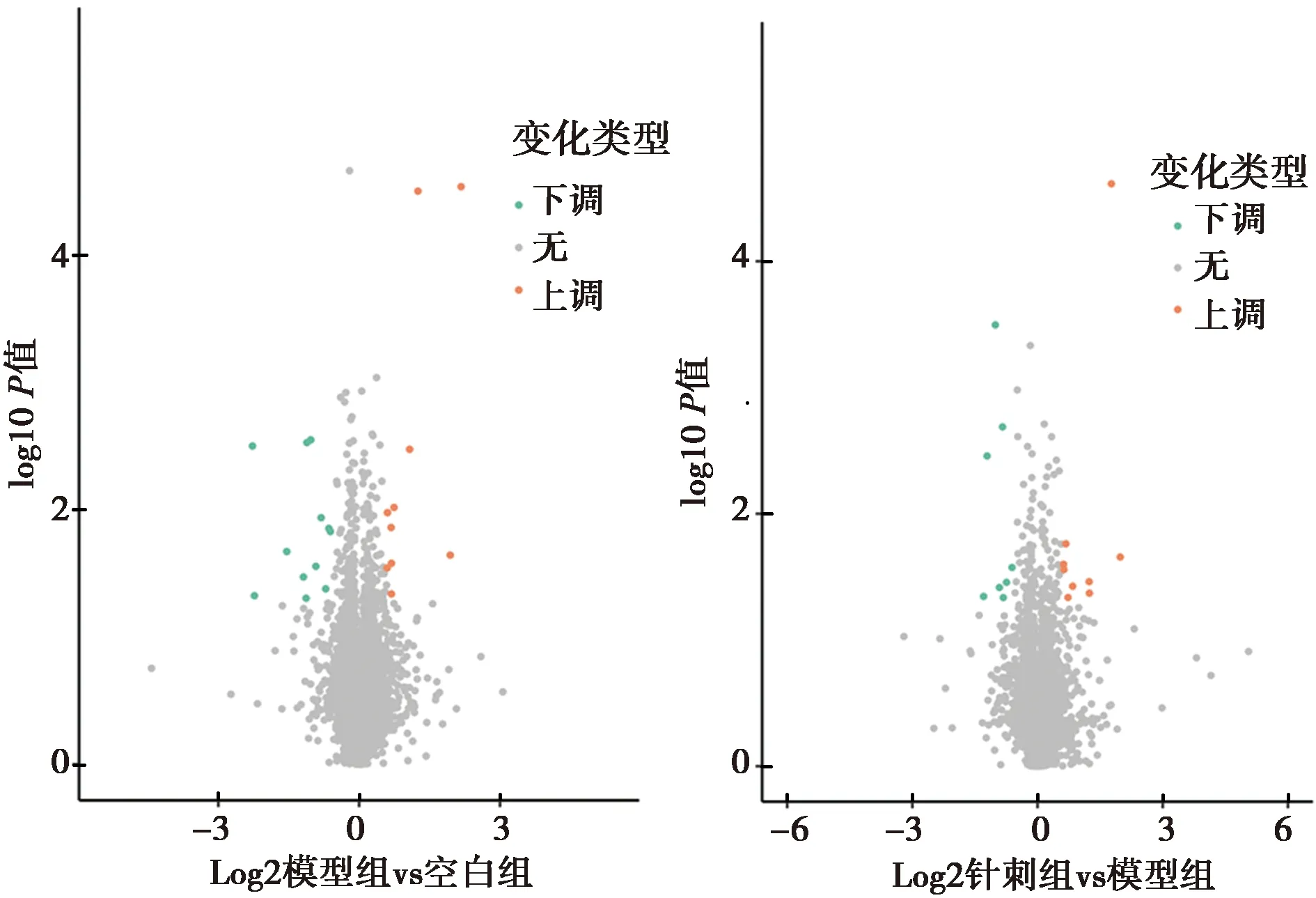
图2 差异蛋白火山图

表2 差异蛋白变化
2.4GO富集分析 针刺组/模型组的差异蛋白主要参与补体激活、调控炎症反应等生物学过程,并在补体激活、体液免疫应答反应、细胞投射组装上具有极显著的富集趋势(P<0.01);与针刺组/模型组的差异蛋白主要参与细胞周期相关调控、RNA代谢等生物学过程,并在细胞周期过程的调控中表现出极显著的富集趋势(P<0.01),见图3。
图3 差异蛋白生物学过程GO富集
2.5聚类分析 根据富集分析得到的富集检验P值使用分层聚类法将不同组相关功能联合绘制为热图,与空白组相比较,模型组蛋白在生物学过程上主要参与蛋白质活化调节、补体激活、炎症反应、免疫调节等;与模型组相比较,针刺组蛋白主要参与细胞周期调节、RNA代谢过程、细胞形态调节等。KEGG聚类分析结果显示模型组参与补体和凝血级联激活过程,针刺组参与基因剪接过程,见图4。
2.6PRM对目标蛋白定量分析 基于GO注释及KEGG富集分析结果,选择了D-多巴色素互变异构酶(Ddt)、Snap29、C1qc、Vtn、Cpped1、Phf5a、Prpf40a、Cdk5rap3、Bbs2、Afg11、Sdf2、Specc1l、Cnrip1等一系列与神经递质合成及补体激活等密切相关的蛋白质进行PRM蛋白定量分析。结果显示:与4D Label-free 结果一致的是,大鼠海马中的Ddt蛋白在模型组下调,而在针刺组上调,Cnrip1蛋白在模型组上调,而在针刺组下调。此外,通过PRM未能识别到Snap29、C1qc、Vtn、Cpped1、Phf5a、Prpf40a、Cdk5rap3、Bbs2、Afg11、Sdf2、Specc1l等蛋白的表达。
3 讨 论
抑郁症发病机制复杂,额叶、丘脑边缘系统等形成的神经网络发生神经和细胞活动改变与抑郁症密切相关,其中海马区域神经元的萎缩和凋亡在抑郁症发病中起重要作用,故常通过海马组织理化变化作为观察和评价抗抑郁疗效的有效指标。本研究表明,针刺可纠正相关蛋白的异常表达来发挥抗抑郁作用;生物信息学分析发现共有18个差异蛋白参与了30种生物学过程,涉及神经递质代谢、神经元自噬、补体激活、信号转导、细胞周期调控、线粒体代谢等。本研究表明通督调脏针法在改善绝望情绪和快感缺失症状上有明显疗效。研究者的前期研究已发现电针百会、印堂可通过拮抗CUMS大鼠海马各亚区蛋白激酶(PK)A、环磷酸腺苷反应元件蛋白(CREB)信号降低改善行为学症状〔11〕,且能修复五羟色胺(5-HT)系统缺陷进而增加脑和突触间隙内的5-HT含量,达到与5-HT再摄取抑制剂(SSRIs)类药物相同的药理作用〔14〕,已有研究表明针刺五脏俞后的抗抑郁疗效与SSRIs药物相当,副作用低且稳定〔12〕,提示通督调脏针法具有明确的疗效和科学依据。
在PRM验证中得到的Ddt是一种蛋白质编码基因,是具有趋化因子样特征的细胞因子。Ddt是巨噬细胞迁移抑制因子(MIF)蛋白超家族的成员,其基因在序列、酶活性和基因结构方面与MIF相关,且两者具有相似的互变异构酶活性及生物相似性〔13〕。MIF与多种疾病相关,在阿尔茨海默病等精神疾病中起重要作用,同时MIF的保护作用和病理作用又与抑郁症有关,研究表明MIF可能通过神经内分泌以及免疫炎症参与抑郁症的发生、发展过程〔14〕。只有Ddt基因是唯一与MIF具有同源性的基因,且与MIF共同定位于22号染色体,在22号染色体上紧密相连〔15〕,笔者推测,Ddt可能和MIF有着一样的作用路线,即在神经内分泌机制调节方面,由分泌促肾上腺皮质激素(ACTH)的垂体细胞分泌和产生醛固酮、糖皮质激素(GC)的肾上腺皮质的上皮细胞表达,当机体处于应激状态时,下丘脑-垂体-肾上腺(HPA)轴激活,外周血中ACTH、GC水平升高,从而对宿主发挥保护作用,此时HPA轴的激活导致Ddt大量产生,并释放到血液循环中。本次研究中,通督调脏针刺后,CUMS大鼠的Ddt过表达,会正向影响黑色素合成,可能与MIF产生相同的作用,从而干扰多巴胺和5-HT等神经递质代谢,影响抑郁发生、发展。神经元自噬的过度激活或抑制可影响神经突触的传递过程而介导抑郁症的发生。CUMS模型可引起海马神经元发生自噬,Snap29通过形成SNARE复合体参与细胞自噬,其过表达可抑制突触传递过程〔16〕,这一发现与本实验结果相符,据此可推测通督调脏针法可通过降低Snap29的表达解除受抑制的突触传递过程而发挥抗抑郁作用。补体经典激活途径与抑郁症存在明显富集关系,通过 GO分析发现这些差异蛋白主要参与了补体激活、体液免疫反应、蛋白质活化级联、蛋白质成熟及炎症反应的调节。有研究表明, C1q和C3定位于未成熟的突触,C3与小胶质细胞表达的补体蛋白C3a受体(C3aR)结合,在无功能的网状突触的发育修剪方面起了重要作用〔17〕。而抑郁模型小鼠杏仁核内发生神经炎症,胶质细胞激活,表达补体C1q、C3,进一步激活小胶质细胞和星形胶质细胞修剪突触,导致突触含量降低,促使小鼠产生抑郁样行为〔18〕。本研究表明,针刺可通过调节补体激活途径的蛋白表达抑制神经炎症,发挥抗抑郁作用。
磷脂酰肌醇-3激酶(PI3K)/蛋白激酶B(Akt)信号通路是近年来研究抗抑郁作用机制的重要通路之一〔19〕,PI3K通过磷酸化下游核心蛋白Akt进而启动下游分子的表达,抑制细胞自噬和细胞凋亡。CUMS大鼠体内PI3K/Akt通路活性被抑制,而Cpped1通过去磷酸化Akt抑制其活性,加重细胞凋亡〔20〕,本实验表明,针刺能够通过降低Cpped1表达解除对Akt的抑制,减少CUMS大鼠的细胞凋亡。此外,本研究发现Prpf40a、Cdk5rap3广泛参与细胞周期调控、细胞周期相变等多个生物学过程,Prpf40a在针刺后的表达量发生显著下调,已有研究表明Prpf突变可导致细胞周期中前mRNA的积累和G1/S转换受损〔21〕,Prpf40a和Prpf属于同源家族,故推测CUMS大鼠受损细胞周期在针刺后得到修复。细胞周期素蛋白依赖性激酶(Cdk)5活性与应激相关疾病存在关联〔22〕,并作为细胞周期抑制剂抑制异常细胞进入,但需要相关调节亚基激活其活性,Cdk5rap3为Cdk5调节亚基相关蛋白,本研究中针刺后的Cdk5rap3由上调转为下调, Cdk5rap3过表达意味着抑郁症发生中存在大量的异常细胞周期,而其表达下调可作为观察抗抑郁作用发挥的有效表征。
线粒体能量代谢失常可导致活性氧(ROS)增加,加剧氧化应激反应,促进细胞凋亡。同时线粒体为神经突触可塑性的重要细胞器,电针后突触和线粒体蛋白发生显著变化,提示电针通过保护海马突触和线粒体功能改善大鼠的抗抑郁性能。本研究中,Bbs2、Afg1l、Specc1l和Sdf2的亚细胞定位于线粒体,Bbs2、Afg1l、Specc1l针刺后表达量显著降低,Sdf2针刺后表达呈上升趋势,尽管目前尚无以上蛋白与抑郁症线粒体代谢失常的明确证据,突触与线粒体在电针干预中的靶向关系亦尚不明确,但这些蛋白可能是通过参与氧化磷酸化过程,降低ROS数量,修复发生断裂的氧化电子呼吸链,进而调控突触可塑性。综上,通督调脏针法可能通过干扰神经递质代谢、调节突触传递过程、抑制神经炎症,从而保护海马神经元免受损伤,进而发挥抗抑郁作用。
本实验还有许多不足之处,如对Ddt蛋白的文献报道较少,需进行针对性研究才可进一步挖掘其抗抑郁途径;再者Bbs2、Afg1l、Specc1l等蛋白在抑郁症的发生、发展过程中的作用,也尚未完全清楚,对这些关键蛋白的深入研究和基因敲除将是我们下一步的研究方向。同时,联合组学技术和静息态功能磁共振成像(fMRI)技术开展经穴特异性研究,观察脏腑辨证和经络辨证结合下穴位个体之间、穴位与经络之间的蛋白表达差异,构建脏腑-经络-经穴-分子的针刺抗抑郁本体理论体系,将是循证医学体系下针刺现代化研究的新走向。
